藏香豬放牧對滇西北高原濕地土壤CO2通量的影響
2019-06-04 08:05:30展鵬飛閆鵬飛劉振亞崔海軍肖德榮
生態學報 2019年9期
展鵬飛,閆鵬飛,劉振亞,王 行,張 贇,張 勇,崔海軍,肖德榮
西南林業大學國家高原濕地研究中心/濕地學院, 昆明 650224
青藏高原濕地碳“匯”功能巨大,其中,沼澤化草甸土壤的碳儲量約占全國土壤碳儲量的23.44%,占全球土壤碳儲量的2.5%,在全球碳循環與碳平衡中發揮著重要的作用[1]。過度放牧是導致高原生態系統結構與功能退化的主要驅動因子。據統計,自20世紀80年代以來,青藏高原就有37%的高寒草甸濕地因過度放牧而嚴重退化[2-3],從而威脅了高原濕地生態系統的碳循環及碳匯功能[4]。土壤CO2通量作為生態系統碳循環重要體現,研究放牧作用下的土壤CO2通量,可為科學掌握放牧對高原濕地碳循環與碳匯功能的影響及其作用機制奠定基礎,并已受到國內外的廣泛關注[5]。
通常認為,放牧通過改變土壤的水熱條件、以及植被組成特征,從而對土壤CO2通量造成影響[6-7]。放牧強度的增加,導致地上生物量逐漸減少,從而減緩了地上部分的碳進入土壤再循環的速率,影響了碳在整個生態系統中的分布格局[8]。研究表明,放牧活動促進了牲畜排泄物的歸還,增加了生態系統中易于降解的有機碳源,導致土壤CO2通量與放牧強度呈顯著的正相關關系[9-10]。也有研究表明,放牧降低了青藏高原東北部高寒草甸土壤的CO2通量,其中,在旱季、重度放牧區域較輕度放牧區,放牧活動減少了地下的生物量,進而導致微生物易分解的碳源也隨之減少,土壤CO2通量降低了近1倍[11-12]。另外,還有研究發現,不同放牧強度下高寒草甸由于觀測時間不足和實驗區域的空間異質性問題,從而導致其CO2通量并無顯著差異[13-14]。由此可見,目前放牧對土壤CO2通量的影響及其作用機制的研究結果還存在較大爭議。因此,開展放牧活動下高原濕地土壤CO2通量的野外觀測及研究具有重要的現實意義。
滇西北高原濕地位于我國青藏高原東南緣、屬我國封閉與半封閉的獨特濕地類型[15]。因地處高寒區域、常年冷濕環境使得濕地生態系統積累了大量的有機物質,成為滇西北自然生態系統最為重要的碳庫之一,對區域碳循環以及大氣溫室氣體的平衡有著重要的功能作用[16]。藏香豬是分布于青藏高原特有豬種,對寒冷與缺氧的自然環境具有較強的適應性[17]。自2009年以來隨著市場對藏香豬需求增加,藏香豬的放牧數量和放牧區域不斷擴大,其中,納帕海濕地藏香豬的載畜量平衡在115頭/km2,超出當地實際承載力(40頭/km2),放牧面積約占整個納帕海面積的5%左右,已經超出了當地的生態閾值。與傳統的牛羊放牧對濕地土壤踐踏干擾不同,藏香豬放牧通過對土壤的翻拱作用,使土壤與植被受到嚴重破壞[18]。據初步統計,由于藏香豬放牧,該區域有近1/4的濕地面積受到不同程度的破壞,并有不斷擴大的趨勢。雖然,藏香豬放牧對滇西北高原濕地的影響現已逐漸受到關注,但是,針對藏香豬這一高原濕地強干擾放牧類型,開展藏香豬放牧對高原濕地生態系統土壤CO2通量影響及作用規律的研究至今還鮮見報道。
本研究以滇西北高原典型濕地納帕海作為研究對象,選擇藏香豬放牧的3個典型區域(布倫、哈木谷、伊拉),并以未受藏香豬破壞的相鄰區域為對照,開展土壤CO2通量研究。主要內容包括:土壤CO2通量的(1)日變化,(2)季節變化對藏香豬放牧的響應,以及(3)土壤CO2通量與土壤理化指標、藏香豬放牧強度之間的關系。通過研究,擬掌握藏香豬放牧對高原濕地土壤CO2通量的影響規律與作用機制,為加強放牧管理、增強高原濕地的保護提供科學依據。
1 材料與方法
1.1 研究區域概況
高原濕地納帕海位于青藏高原東南緣、橫斷山脈中段的香格里拉市境內(27°49′—27°55′N,99°37′—99°41′E),最低海拔3260 m,面積3100 hm2,是我國低緯度高海拔濕地的獨特類型。該區域氣候屬高原寒溫性濕潤氣候,受南北向排列的山地和大氣環流的影響,全年盛行南風和南偏西風,年均溫僅為5.4 ℃,旱雨季分明,6—10月為雨季,11月至次年5月份為旱季。由于旱雨季所帶來濕地水位變化、以及海拔高程的差異,納帕海在空間上分為水體、沼澤、沼澤化草甸、草甸等土地利用類型。其中,沼澤化草甸與草甸面積2174.85 hm2,占整個濕地總面積的70.1%,為該地區的主要放牧區域[19]。
納帕海具有豐富的生物多樣性,分布有黑頸鶴(Grusnigricollis)、黑鸛(Ciconianigra)等國家I級保護鳥類,極具生物多樣性和特有性保護價值。另外,該地還是許多植物物種的交匯區,植物多樣性極其豐富,分布有杉葉藻(Hippurisvulgaris)、水蔥(Schoenoplectustabernaemontani)、茭草(Zizanialatifolia)等一些水生植物。1984年,經云南省政府批準建立納帕海省級自然保護區,2005年,被指定為國際重要濕地,成為國內外關注生物多樣性保護的熱點地區[20]。納帕海濕地積累了大量有機物質,有些地區甚至堆積了極其深厚的泥炭層,成為濕地生態系統碳匯的基礎,對該區域碳循環和大氣溫室氣體的平衡起著重要的調節作用[21]。
1.2 樣地設置
藏香豬放牧快速增長于2009年,但由于藏香豬對土壤的反復翻拱,現已無法準確判斷其具體的干擾時長。因此,選擇布倫、哈木谷和伊拉3處藏香豬放牧的典型的區域,其中,布倫草原屬于沼澤化草甸,土壤類型為泥炭土,季節性淹水。哈木谷和伊拉草原屬于陸生草甸,土壤類型為草甸土,常年無淹水。布倫區域的土壤母質為高度發育的泥炭層,哈木谷和伊拉區域的土壤母質為河流沖積物,草根層直接發育在潛育層上,潛育層常年積水,土體下部常呈淺綠色。采樣點分別設置藏香豬干擾區(Disturb area)和對照區(Control area)。結合現場調查、高分衛星遙感影像數據判讀以及草根層發育狀況分析,確保所選取的對照樣方在歷史上沒有被藏香豬干擾過,能夠反映該區域的長期放牧效應。同時,在3個草原分別設置藏香豬干擾和對照的平行研究樣帶,不同放牧方式下植物生物量指標如表1所示。各研究樣帶上每隔5 m設置1個1 m×1 m的研究樣方,每條樣帶共設置研究樣方6個,3個研究區域共計36個研究樣方。各研究樣帶與研究樣方四角用木樁標記并用GPS定位(圖1)。
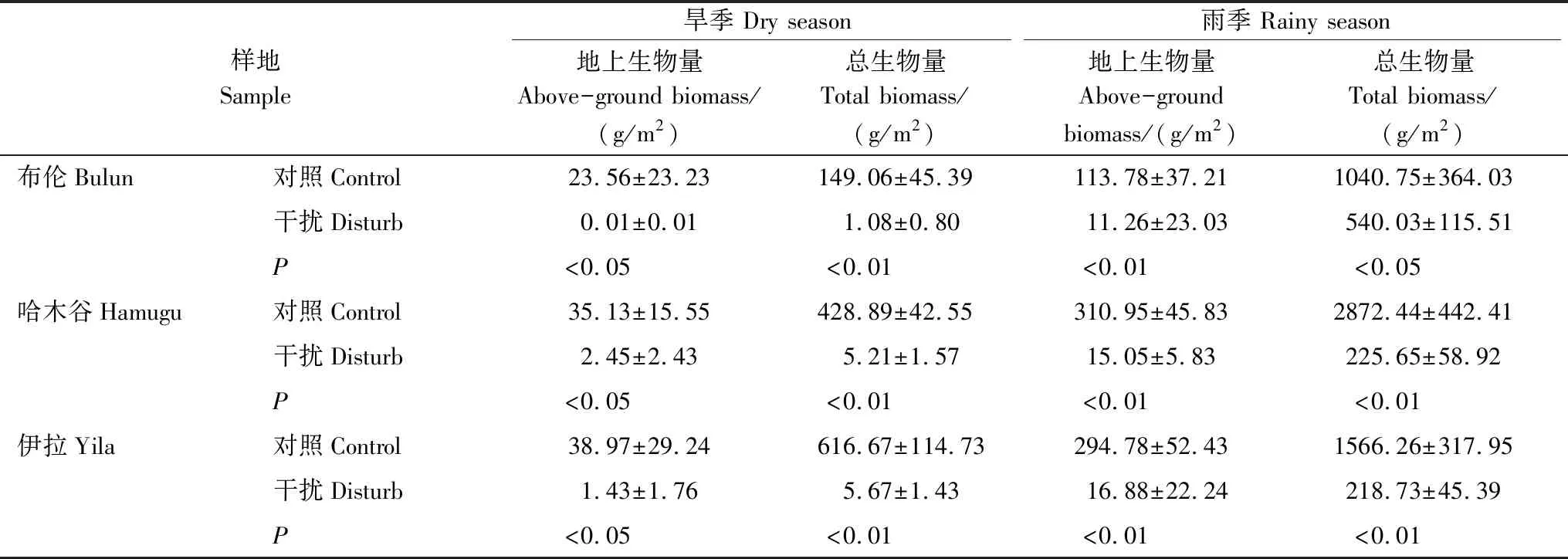
表1 不同放牧方式下植物生物量指標
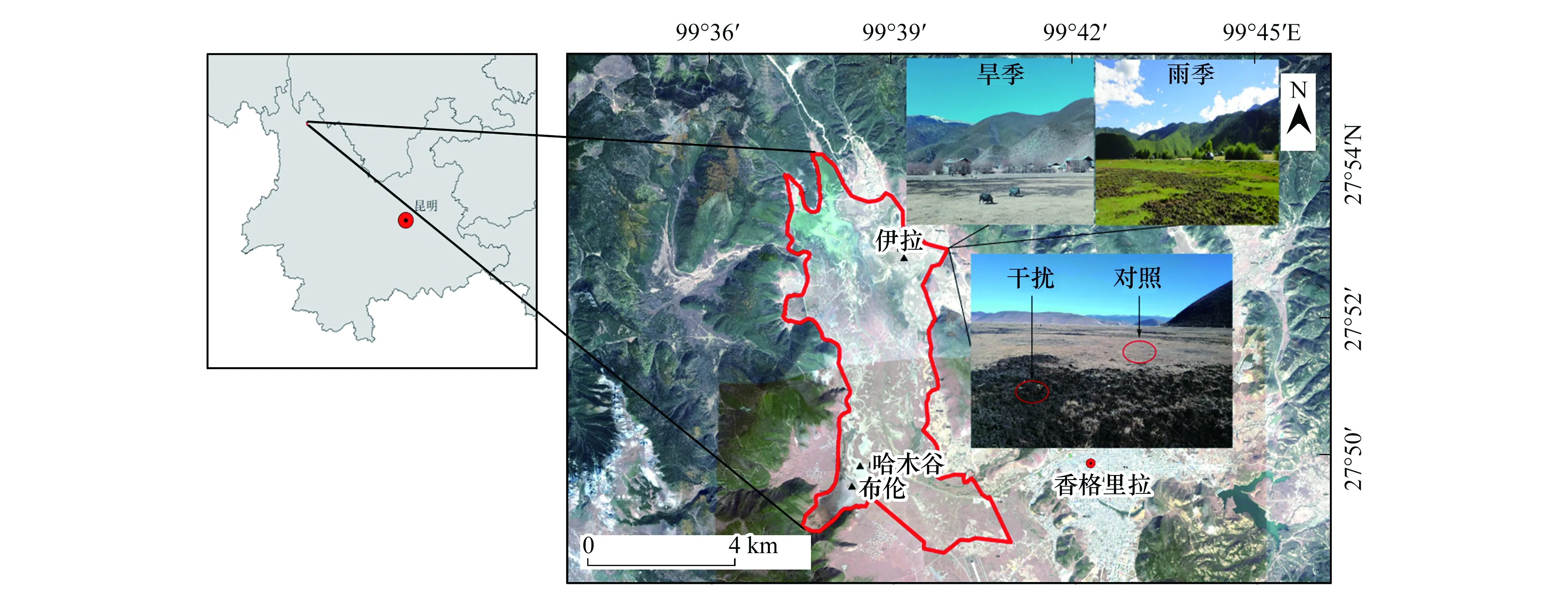
圖1 納帕海位置圖和研究地點示意圖Fig.1 Location of Napahai and sampling sites
1.3 野外監測與室內分析
1.3.1土壤CO2通量測定
土壤CO2通量采用便攜式土壤呼吸儀(LI-8100A,LI-COR,USA)現場測定CO2排放通量。分別在2016年雨季(7月)和2017年旱季(1月)選擇有代表性的時間、天氣狀況對布倫、哈木谷和伊拉草原上藏香豬翻拱和對照樣方土壤CO2通量進行測定。野外采樣時,將測定系統的氣室與基座密切相連,連接電池后,測定前對機器進行開機預熱20 min,設置網絡,將測定系統與配套筆記本電腦(用于存儲數據)連接,調整好參數。每次測量時,首先,將預先制好的PVC環墊(ID=20 cm,H=12 cm)打入每塊樣地內的待測點,打入土壤中的深度大約1—2 cm,并砸實外圈土壤以防漏氣,并在整個測定過程中保持PVC環靜止不動。在避免破壞土壤表層結構的前提下將PVC環墊內的活體植物的地上部分剪掉并移出,以消除測定時植物自養呼吸對土壤CO2排放通量產生的影響,并盡量避免對土壤表層結構的破壞。安排的測定時間為每天的8:00—17:00,連續3d對3個采樣點進行分別測定。每個樣方設置測量時間長度為10 min,測定結束后迅速移動至下一個樣方,每條樣帶6個樣方共計1 h,然后對干擾和對照樣帶上的樣方進行循環測定,且保證干擾和對照樣帶同時進行。
1.3.2植物生物量與土壤理化性質測定
在布倫、哈木谷、伊拉草原樣每個待測樣點周圍選擇一個樣方(大小為50 cm×50 cm),采用五點混合法在每塊樣方使用直徑5 cm環刀,取樣深度為20 cm,每條樣帶6個重復。將環刀裝好的土樣置于標記好的自封袋中,帶回實驗室,風干、過篩后分離根系和土壤,在65 ℃烘箱中烘干,稱重,計算植物地上和地下生物量指標。參考《土壤農化分析》對土壤基本土壤理化指標進行測定(見表2)[22]。其中,土壤有機質采用重鉻酸鉀容量法;pH值采用酸度計法;總活性碳庫利用KMnO4氧化法。利用環刀法一次性連續測量土壤容重、總孔隙度、土壤含水率,根土比等基本理化指標。
1.4 藏香豬放牧強度的確定方法
放牧干擾通常直接作用于濕地植被、土壤等,而濕地植被、土壤本底特征可較好體現放牧的強度。因此,對采樣區植物、土壤環境本底的相關參數進行同步監測與研究,通過植物-土壤指數(plant-soil index,PSI)進而分析藏香豬干擾型放牧對高原濕地的影響[23],即:
式中,P為植物群落狀態,S為土壤狀態。其中,植物群落狀態(P)的評估指標包括植物地上生物量和地下生物量,Pi為植物群落指標i,Wpi為該指標的權重。土壤狀態(S)的評估指標包括土壤的物理指標(容重,含水率,根土比,孔隙度)和化學性質(Ph,有機質,總活性碳)。Sj為土壤指標j,Wsi為該指標的權重。采用熵權法,按標準化計算熵值和確權,以確定植物群落狀態(P)和土壤狀態(S)各指標的權重[24]。PSI值越大表明植物-土壤系統狀態越好,其受藏香豬放牧的影響越小,PSI越小表明植物-土壤系統受藏香豬放牧的影響越大,放牧強度較大。進而通過PSI確定藏香豬放牧對高原濕地的影響。
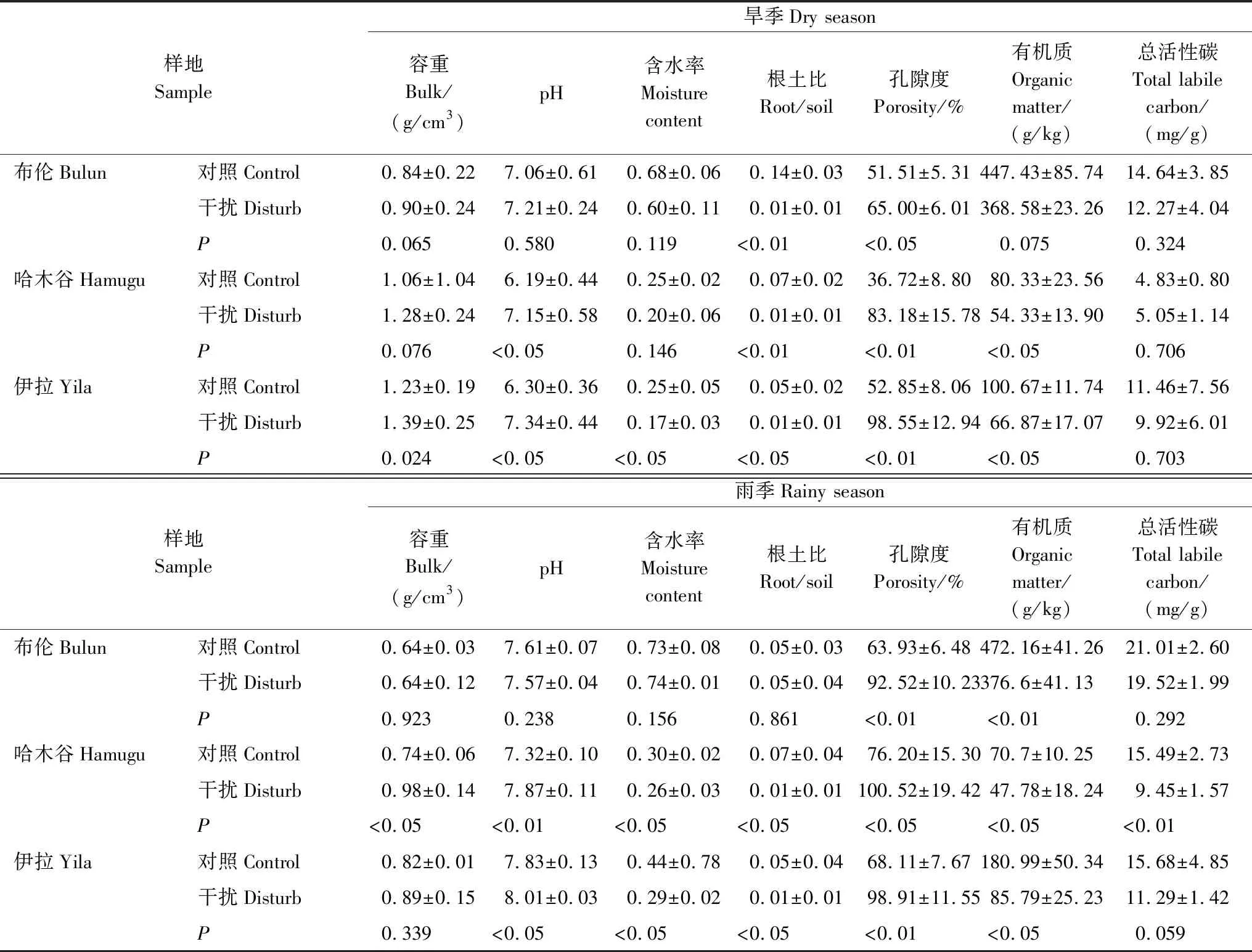
表2 土壤基本理化指標
1.5 統計分析
采用單因素方差分析法 (one-way ANOVA) 分析了不同放牧方式下土壤物理、化學性質及生物量之間的差異性。利用多因素方差分析法(Multiple-way ANOVA),分析了季節、放牧方式、采樣點對土壤CO2通量及植物-土壤指數的交互影響。最后,分別利用每個采樣點土壤理化指標、生物量指標、植物-土壤指數與土壤CO2通量的統計學差異,構建與CO2通量的線性回歸方程。上述分析均采用SPSS 16.0進行統計分析,其中P<0.05為顯著,P<0.01為極顯著,作圖采用Sigmaplot 10.0。
2 結果
2.1 土壤CO2通量的變化特征
從季節上整體來看(圖2),雨季影響下的土壤CO2排放通量的平均值較旱季明顯升高,且藏香豬干擾型放牧嚴重消弱了土壤CO2的排放速率,其中布倫、哈木谷、伊拉草原較對照分別降低了70.4%、87.5%、60.7%。從采樣點來看,不同季節影響下的布倫草原對照下的土壤CO2通量均大于干擾樣地,其中旱季布倫草原藏香豬干擾較對照CO2排放通量降低了30.2%,而雨季降低了70.4%。相比布倫草原,藏香豬干擾影響下的哈木谷和伊拉草原在旱季時則出現了土壤CO2通量大于對照的現象,其中哈木谷草原藏香豬干擾下的土壤CO2通量與對照相比差異十分顯著(P<0.01)。
由圖3可知,不同季節影響下的布倫、哈木谷、伊拉草原藏香豬干擾與對照影響下的土壤CO2通量的日變化曲線存在差異,具有明顯的日波動變化特征。旱季時,布倫草原無論是對照還是干擾樣地,土壤CO2通量的最高值均出現在10:00—11:00之間,其數值分別為1.05 μmol CO2m-2s-1和0.94 μmol CO2m-2s-1,相比布倫草原,哈木谷和伊拉對照影響下的土壤CO2通量在一天之內呈不斷上升的趨勢,且在傍晚時分超過了對照樣地且達到全天最大值。雨季時,各采樣點里藏香豬干擾影響下的土壤CO2通量均小于對照土壤,各采樣點土壤CO2通量的全天最高值均出現在午后,數值分別為3.06 μmol CO2m-2s-1、7.49 μmol CO2m-2s-1和5.01 μmol CO2m-2s-1。同時,無論是對照還是藏香豬干擾,其CO2通量日變化波動曲線與旱季相比更為平穩。
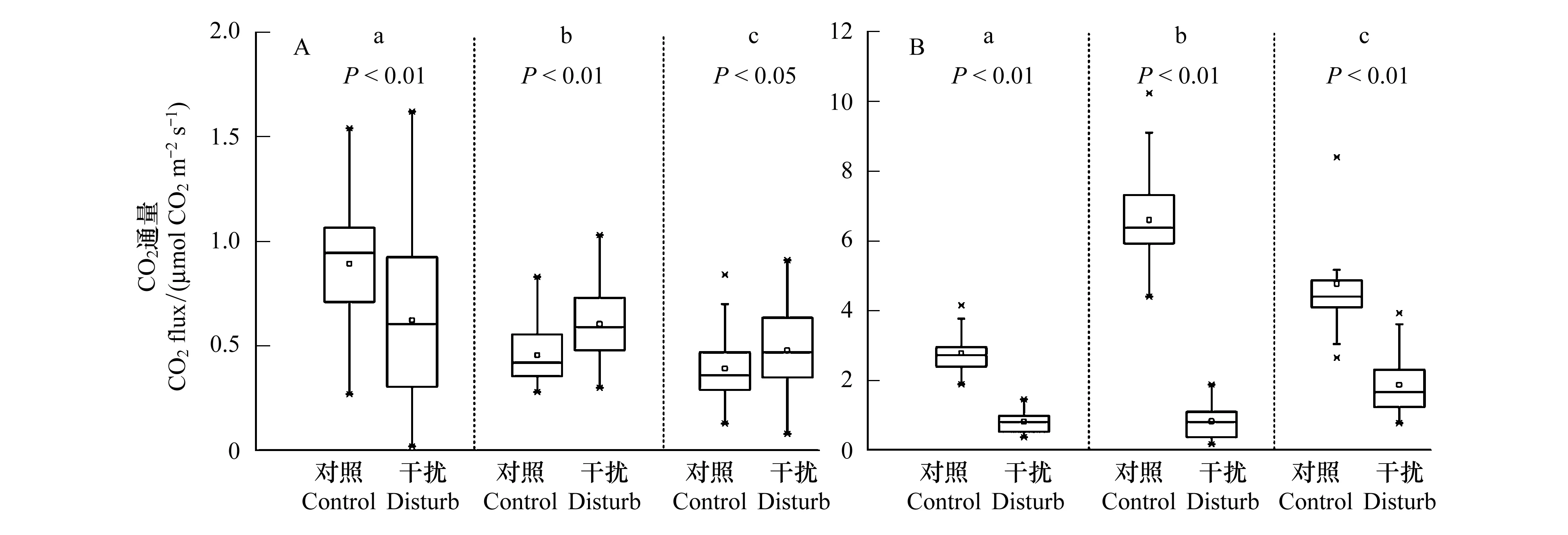
圖2 不同采樣點土壤CO2通量的變化特征Fig.2 Characteristics of soil CO2 fluxes in different sampling siteA:旱季; B:雨季; a:布倫, b:哈木谷, c:伊拉
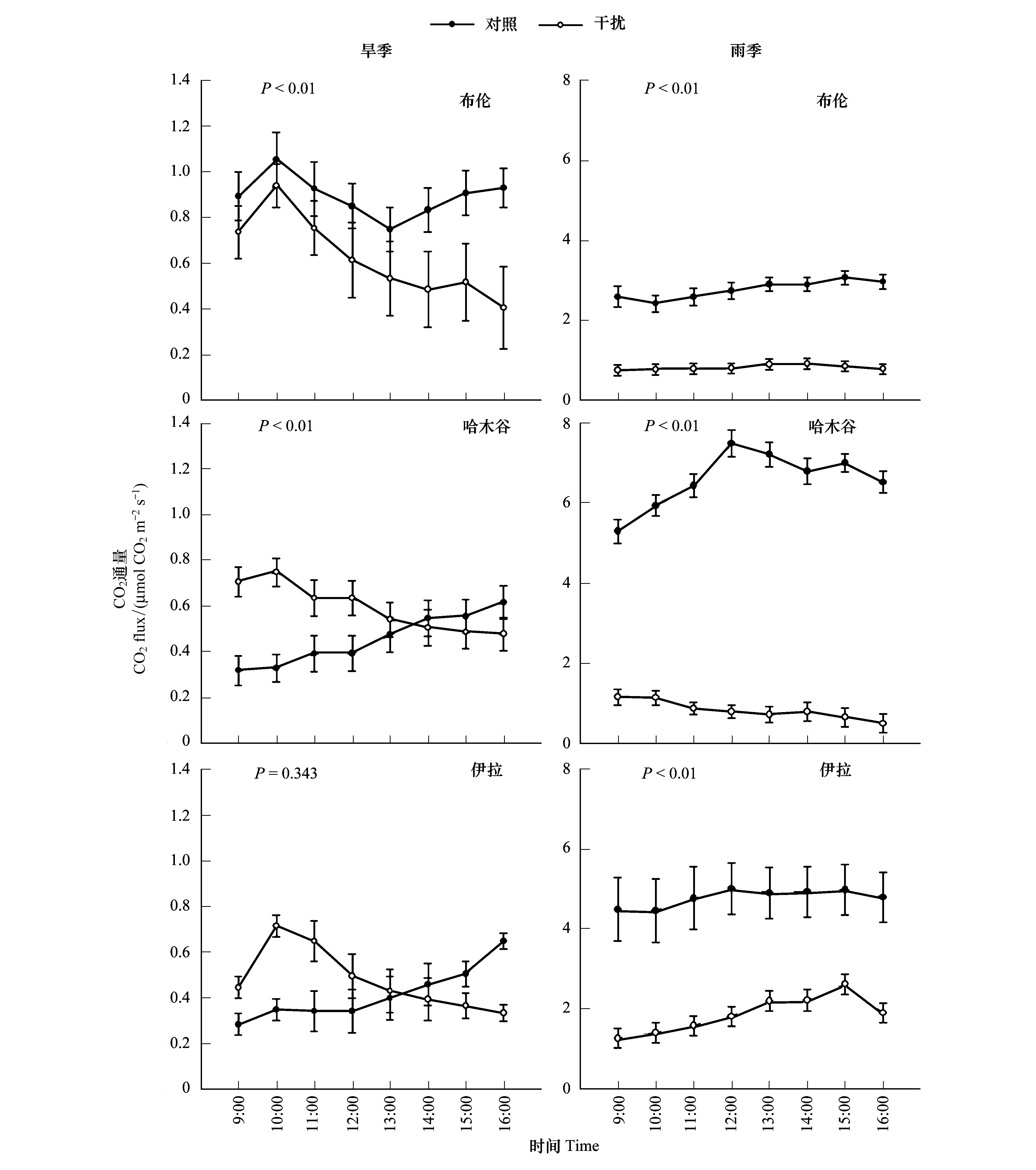
圖3 不同采樣點土壤CO2通量的日動態變化特征Fig.3 The daily dynamic characteristics of CO2 fluxes at different sampling site
2.2 土壤CO2通量與土壤理、化性狀的相關性
由圖4可知,對照樣地下土壤容重與CO2通量存在顯著的負相關關系(P<0.01),且土壤CO2排放通量隨著土壤容重的增加呈下降趨勢。同時,無論是對照還是藏香豬干擾,不同采樣點里的土壤含水率、根土比與CO2通量之間均不存在相關性關系(P>0.05)。然而,通過對比分析藏香豬干擾和對照下的土壤孔隙度與CO2通量的關系表明,對照和藏香豬干擾下的土壤CO2通量與孔隙度都擁有顯著的正相關關系(P<0.05),且土壤CO2通量隨著土壤孔隙度的增加呈線性增長模式,其中R2值平方分別為0.3642和0.147。
通過對土壤化學指標與CO2通量進行線性擬合分析(圖5),研究發現,無論是對照還是藏香豬干擾下的土壤pH都與CO2通量呈明顯的正相關性(P<0.05)。而土壤有機質在藏香豬干擾和對照影響下均與CO2通量無任何相關性關系(P>0.05)。同時,通過對比分析藏香豬與對照下的土壤CO2通量與總活性碳的關系,結果表明,對照較藏香豬干擾放牧土壤CO2通量與總活性碳擁有更加顯著的相關性。
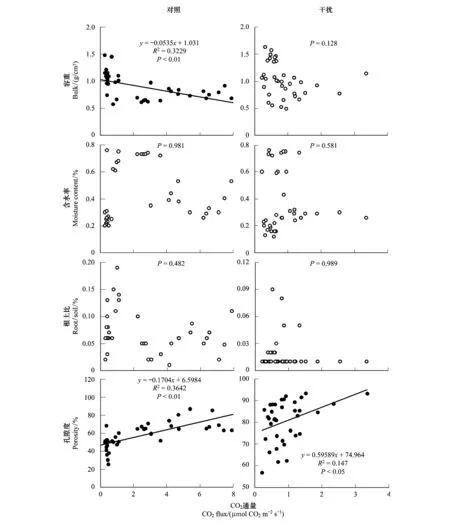
圖4 土壤CO2通量與土壤物理性狀的線性回歸關系Fig.4 Linear regression relationship between soil CO2 fluxes and soil physical properties
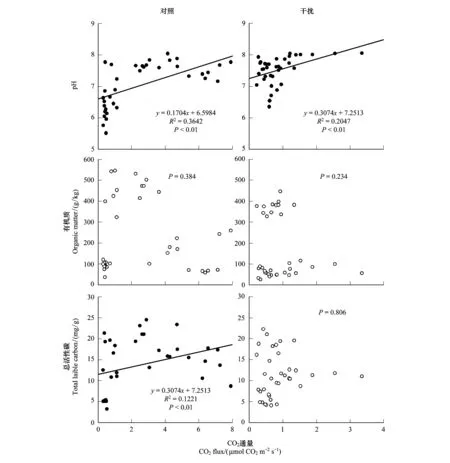
圖5 土壤CO2通量與土壤化學性狀的線性回歸關系Fig.5 Linear regression relationship between soil CO2 fluxes and soil chemical properties
2.3 土壤CO2通量與植物生物量的相關性
通過對采樣點植物生物量與土壤CO2通量數值進行擬合分析,如圖6可知,采樣點中對照樣地下的植物生物量與CO2通量呈顯著的正相關關系,土壤CO2排放通量隨著植物生物量的增加呈現出顯著的增加趨勢(P<0.01)。另外,我們也發現藏香豬干擾影響下的生物量與CO2通量的變化無相關性(P>0.05)。由此表明,生物量是影響土壤CO2排放通量的關鍵要素,藏香豬干擾型放牧,破壞了濕地植被,進而引起土壤CO2排放通量發生了無規律的波動變化。
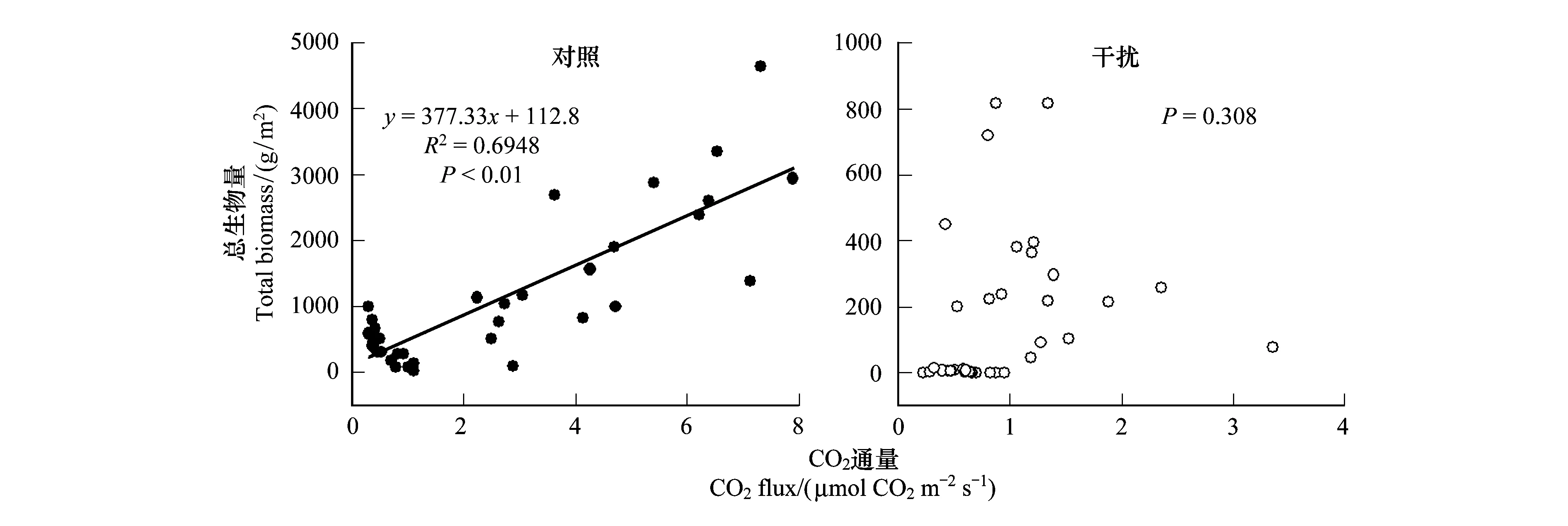
圖6 土壤CO2通量與植物生物量的線性回歸關系Fig.6 Linear regression relationship between soil CO2 fluxes and plant biomass
2.4 土壤CO2通量與PSI的關系
采用線性回歸方程對不同樣帶下高原濕地土壤CO2通量日平均值與植物-土壤指數進行擬合分析,得到了對照及藏香豬干擾下的高原濕地土壤CO2通量與植物-土壤指數之間的關系(圖7)。如圖所示,對照樣地中土壤CO2通量與PSI達到了極顯著相關的關系(P<0.001),R2為0.6606。由此說明該線性模型能很好的模擬土壤CO2通量與植物-土壤指數之間的關系。總體來看,PSI對非干擾條件下CO2排放通量預測效果更好,而放牧增加了CO2排放通量的不確定性,減弱了PSI的預測能力。
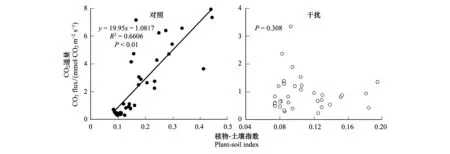
圖7 植物-土壤指數與CO2通量的線性回歸關系Fig.7 Linear regression relationship between plant-soil index and CO2 fluxes
通過植物-土壤指數與CO2通量日平均值的多因素方差分析(表3)表明,不同季節,放牧方式,采樣點對土壤CO2通量均產生了顯著影響(P<0.01),其中,不同季節與放牧方式對植物與土壤狀態的影響十分顯著(P<0.01),而采樣地點的不同對植物-土壤指數無顯著影響(P>0.05)。季節、采樣點與放牧方式之間存在明顯的交互作用,其中,植物-土壤指數受到不同放牧方式的影響,其影響程度與季節和采樣地點均相關(P<0.05),而土壤CO2通量只有在不同季節與不同放牧方式的交互影響下才更為顯著(P<0.01)。
3 討論
3.1 放牧干擾對高原濕地土壤CO2通量的影響
土壤CO2排放通量是一個受眾多因素影響的生物學和非生物學的過程[25]。放牧作為草地最主要的利用形式,同時也是影響高原濕地土壤CO2通量的主要人為干擾因素[26]。放牧過程通過牲畜采食植物、踐踏土壤以及排泄物歸還等多種途徑直接或是間接影響了草地CO2的排放通量[27]。研究發現,放牧往往會促使生物因子和非生物因子發生改變,導致高原濕地不斷退化,土壤肥力、微生物數量與種類降低,土壤生化活性減少,從而影響了土壤CO2的排放通量[28]。適度放牧使草地生物量、根系和凋落物數量、土壤動物和微生物的多樣性提高,從而促進土壤CO2通量的排放[29]。過度放牧則導致草地退化、蓋度降低、草地地上和地下生物量下降,草地碳匯功能減弱,微生物易分解的碳源及微生物自身的數量也隨之減少,從而降低土壤CO2通量[30]。藏香豬翻拱型放牧作為強干擾放牧的典型代表,研究發現,雨季藏香豬干擾下的CO2通量要明顯低于對照土壤,分析原因,首先,雨季氣溫高降水多,植被生長茂盛,而在藏香豬干擾放牧條件下,地表植被和地下根部被大量啃食,導致了地表植物生物量急劇減少,CO2通量明顯降低。藏香豬干擾型放牧改變了植物冠層結構以及植物根系分泌物的量,降低了植物的自養呼吸,從而緩解地上部分中的碳進入土壤再循環的速率,影響了碳在整個生態系統中的分布格局[31-32]。除此之外,土壤微生物與CO2排放速率也具有十分密切的關系。受強放牧干擾的影響,土壤微生物的數量、結構、組成、功能都會發生顯著的變化,微生物多樣性顯著降低[33]。外界干擾改變了土壤理化性質進而影響了土壤微生物群落結構,隨著放牧強度增加,微生物數量增加,土壤呼吸作用減弱[34]。由此表明,藏香豬干擾放牧破壞了土壤原有的結構,改變了微生物的分布與組成,進而抑制了土壤CO2排放通量。
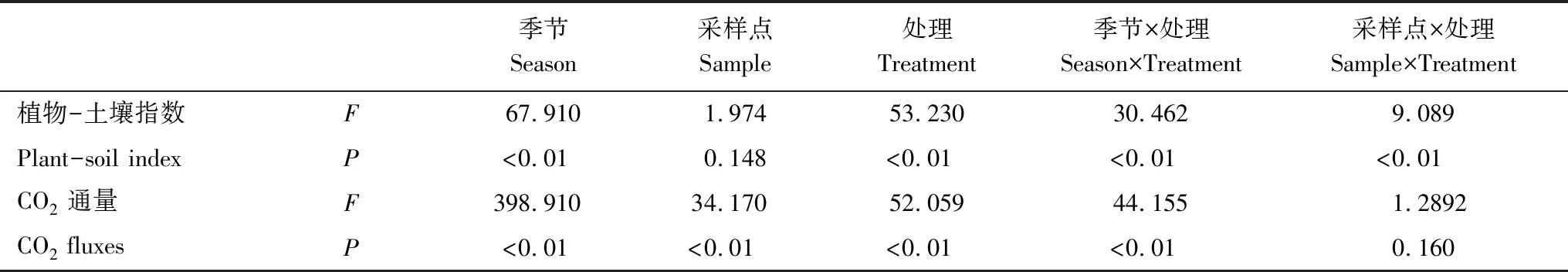
表3 植物-土壤指數與CO2通量的多因素方差分析表
還有研究表明,在眾多土壤理化因子中,容重對土壤CO2的排放通量影響最大,這是因為土壤是一個多孔的系統,根系和土壤微生物呼吸釋放的CO2首先聚集在這些空隙中,然后遵循物理學擴散原理逐漸釋放到大氣[35]。我們研究發現,對照中土壤容重、孔隙度、pH、總活性碳與土壤CO2通量均呈現出顯著的相關性關系,而藏香豬干擾影響下的土壤除了與pH和孔隙度表現出較為明顯的正相關性外,與其他土壤物理化學性質均無相關性。分析原因,藏香豬強干擾型放牧較對照更容易造成土壤緊實度的增加,進而導致土壤孔隙度與水穩性團聚體減少,引起透水性、透氣性下降,阻礙了氣體的順暢排放。盡管如此,我們還是發現無論是藏香豬干擾還是對照,土壤CO2通量與土壤pH均呈現出較好的相關性,這很可能與長期放牧影響下土壤環境下微生物活性有關[36]。另外豬拱強干擾放牧破壞了土壤原有的結構,導致土壤有機質降低,植被覆蓋減少,使土壤物理與化學性質呈現出無規律的變化特征。
然而,還有研究者通過對若爾蓋高原泥炭地和內蒙古天然草原CO2通量的動態監測發現,放牧干擾對若爾蓋高原泥炭地和內蒙古天然草原CO2通量并沒有顯著影響[37-38]。這與我們研究發現的旱季高原濕地不同采樣點CO2通量的差異性不顯著的結果基本一致。同時,我們還發現旱季,在區域日動態變化上藏香豬干擾影響下出現了土壤CO2通量大于對照的現象。探究其原因,一方面,哈木谷和伊拉草原原本屬于陸生草甸,采樣地多為草甸土,旱季時,本身失去了地表植被保護的高原濕地土壤,受藏香豬干擾破壞的程度明顯加深,另外,藏香豬放牧導致土壤劇烈翻滾,促使表層營養物質持續更新,提高了土壤營養物質的循環速率,增強了土壤CO2排放通量[39]。另一方面,藏香豬翻拱取食行為,將導致植被蓋度降低,促使土壤表層容易接受到更多的陽光,藏香豬不斷地取食與踐踏使得地表土壤溫度迅速提升,而這些均有利于表層土壤有機質的分解,使生物活性得以改善,提高了CO2的排放通量[40]。此外,我們還發現,旱季對照下的布倫草原CO2通量的日平均值顯著大于干擾土壤,由此可以預測出冬日里的泥炭地可能是主要的土壤CO2釋放源。雖然如此,但是關于旱季藏香豬干擾放牧較對照增強了土壤CO2通量的主要原因,還有待我們后期進一步探索。
3.2 植物-土壤指數對高原濕地土壤CO2通量的影響
高原濕地生態系統的地上、地下部分是相互聯系的。地表植被種類以及生長狀況的不同,會相應的改變所在地下土壤微環境,從而影響了土壤CO2通量的變化[41]。植物與土壤作為高原濕地生態系統中的重要組成部分,它們之間的相互作用是高原濕地生態系統地上、地下結合的重要紐帶[42]。雨季時,植物生物量的多寡較好的指示了土壤CO2通量的變化,較多的地上生物量增加了土壤有機碳來源,提升了土壤微生物可分解底物的濃度,同時地下生物量根系呼吸作用強,從而增加土壤CO2速率[43-44]。旱季時,土壤失去了植物覆蓋,影響CO2通量的主要因素將由地上植物轉變為枯落物及土壤本身,在同樣受到藏香豬干擾影響的高原濕地上,枯落物及土壤本身較地表植物對土壤CO2通量的影響相對較小。因此,這也就能很好地解釋了在季節尺度上,同樣受到藏香豬干擾放牧的影響下旱季的土壤CO2通量比雨季更低的原因。
部分學者根據生態學中經典的“中度干擾理論”,結合“植物-動物間互作的補償性”觀點,提出放牧優化假設,認為植被生物量與放牧強度之間的關系是非線性的,即隨著放牧強度的增加,草地的生物量先增加,后下降的趨勢特征[45]。為此,我們結合實際情況對采樣區植物、土壤環境本底的相關參數進行同步監測與研究,通過構建植物-土壤指數,用于指示放牧活動對土壤狀態的影響。在整個研究階段,藏香豬干擾放牧破壞了土壤原有的結構,形成大量次生裸地,對高原濕地牧草生產力造成了極大破壞,植物-土壤指數明顯低于對照土壤,且相應的土壤CO2通量也較低。總體來看,PSI在對照樣地里對土壤CO2排放通量預測效果更好,而藏香豬放牧增加了土壤CO2排放通量的不確定性,減弱了PSI的預測能力。
3.3 藏香豬放牧影響下的高原濕地碳循環機制
陸地生態系統碳循環以大氣-陸生植物-土壤-大氣的形式循環,在全球碳循環中起著特別重要的作用[46]。而高原濕地生態系統碳循環的主要過程是在土壤中,在這種意義上,高原濕地生態系統近似于土壤-大氣系統[47],高原濕地的地上生物量中的碳貯量和土壤中貯存的碳含量相比是較少的,而且碳素由植被部分進入土壤的循環途徑相對單一,通常在植物雨季結束后即可完成。在這種近似土壤-大氣系統碳循環的過程中,土壤有機質(含凋落物)的分解速度以及植物向土壤的輸入速度是支配整個系統碳循環功能最關鍵的變量。藏香豬干擾型放牧破壞了土壤結構,導致地表植被單一,土壤有機碳含量降低,進而影響了CO2的排放通量,破壞了高原濕地碳循環的正常進程。自然狀態下,高原濕地生態系統的碳收支基本保持平衡,而高原濕地退化將會造成植被生產力的降低,植物凋落物減少,從而導致土壤碳庫入不敷出,高原濕地生態系統碳由“匯”向“源”轉變[48]。高原濕地作為生態系統中重要的碳源,我們研究發現,藏香豬干擾影響下的CO2的排放通量與對照相比明顯降低。由此表明,散養型藏香豬在尋覓食物的過程中對土壤進行劇烈翻拱,一方面破壞地表植物造成亞表層土壤裸露,另一方面破壞地下草根層造成土壤結構發生改變。土壤有機質礦化、養分流失,從而使濕地生態系統功能退化,嚴重影響生態系統的碳“匯“與“源”的功能。
4 結論
藏香豬干擾型放牧消弱了高原濕地土壤的CO2的排放通量,具有明顯的日波動和季節變化特征。同時,藏香豬放牧對高原濕地土壤和植被擁有較強干擾與破壞能力,進而導致土壤理化性狀、植物生物量與CO2通量基本呈現出無規則的變化。通過植物-土壤指數與土壤CO2通量進一步擬合分析可知,藏香豬干擾下的植物-土壤指數顯著低于對照土壤,且相對應的土壤CO2通量也較低,構建植物-土壤指數可以有效的估算納帕海高原濕地與大氣之間的碳交換量。總之,我們需要加強對藏香豬放牧的科學管理及其生態閾值研究,并對其破壞力進行客觀評價,分析藏香豬放牧的環境容量,并找出能夠維持土壤生產力較為合理的藏香豬放牧強度,進而為滇西北高原生態環境的保護、恢復及重建提供理論支持。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年3期)2017-02-21 01:22:28
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
