不同黃瓜砧木對低溫弱光脅迫的響應及與ABA含量的關系
2019-06-04 10:58:48李福德畢煥改艾希珍
中國蔬菜 2019年5期
李福德 付 鑫 畢煥改艾希珍
(山東農業大學園藝科學與工程學院,作物生物學國家重點實驗室,農業部黃淮地區園藝作物生物學與種質創制重點開放實驗室,山東泰安 271018)
選用抗性砧木進行嫁接栽培可提高瓜類、茄果類等蔬菜作物對低溫、高溫、干旱和鹽漬等非生物脅迫的抗性(Zhou et al.,2007;Han et al.,2013;Li et al.,2014;Xing et al.,2015),但不 同砧木對接穗的影響存在顯著差異。于賢昌等(1998,1999)研究了不同耐冷性的南瓜砧木與黃瓜接穗對嫁接苗的影響,發現耐冷性均較強的接穗與砧木嫁接時才可獲得耐冷性最強的嫁接組合,且嫁接苗耐冷性的增強與ABA、GA等激素的含量變化有關。李思思等(2018)研究指出,選用五葉香絲瓜作為砧木嫁接黃瓜較黑籽南瓜、日本南瓜等砧木能更好地提高黃瓜嫁接苗對根際高溫的耐受性。田雪梅等(2012)選用不同耐鹽性的南瓜砧木分別與新泰密刺黃瓜嫁接,嫁接苗對NaCl脅迫的響應不同,且嫁接苗耐鹽性的高低與葉片Na+/K+有關。因此,探明不同砧木對非生物脅迫的響應機制有利于生產上不同抗性砧木的篩選。
黃瓜(Cucumis sativusL.)是北方日光溫室的主栽蔬菜,在栽培過程中經常遭遇非生物脅迫逆境,尤其是秋冬茬日光溫室黃瓜栽培中低溫弱光環境最為常見,造成黃瓜產量和品質大幅下降(艾希珍 等,2004)。嫁接因其經濟、簡單、有效成為增強設施黃瓜耐冷性的主要手段,黑籽南瓜耐冷性較強,被廣泛用于黃瓜嫁接生產,但因其不具備脫蠟粉能力,常導致嫁接苗果實外觀和品質下降(Lee et al.,1999;李紅麗 等,2006;劉青 等,2012)。因此,篩選耐冷性強,且對接穗品質影響較小的白(黃)籽南瓜砧木對日光溫室黃瓜嫁接生產具有重要的意義。本試驗以不同白籽南瓜為試材,以常規砧木黑籽南瓜為對照,通過分析低溫脅迫下不同砧木耐冷指標和ABA含量的變化,篩選出耐冷性強的白籽南瓜砧木,并探明其高耐冷性與ABA的關系,以期為優化黃瓜嫁接栽培提供優良砧木。
1 材料與方法
1.1 試驗設計
試驗于2017年在山東農業大學南校區試驗站進行。以津優35號黃瓜和常規砧木黑籽南瓜為對照,以金媽媽519、金媽媽619、格頓聯豐、格頓西光、格頓金凱、博強102、博強103、博強105、博強4號等白籽南瓜砧木為試材(表1),4月5日播種于日光溫室8cm×8cm營養缽中。每品種播種100株,選取80株正常生長幼苗進行試驗。溫室環境為:光量子通量密度(PFD)日均值為600μmol·m-2·s-1,晝 /夜溫度均值為25℃/18℃。待幼苗長至一葉一心時移入日本山崎黃瓜專用營養液(含0.5mmol·L-1NH4H2PO4,2.0mmol·L-1Ca(NO3)2·4H2O,3.2mmol·L-1KNO3和1.0mmol·L-1MgSO4·7H2O,微量元素為全濃度,電導率2.2~2.5mS·cm-1,pH值6.8~7.0)進行水培,至三葉一心時(5月3日)轉移至寧波產光照培養箱中進行低溫弱光處理,條件如下:晝/夜溫度8℃/5℃,光照100μmol·m-2·s-1,光周期11h/13h(晝/夜)。
表1 試驗材料及來源
1.2 測定項目
1.2.1 冷害指數 冷害指數的調查參照于賢昌等(1998)的方法。幼苗冷害癥狀分級標準:0級,葉片正常,無受害癥狀;1級,葉片稍皺縮,第1葉或第2葉葉緣發黃或略失水,第3葉和心葉無受害癥狀;2級,葉片皺縮,第1葉和第2葉葉緣嚴重失水,第3葉葉緣發黃或略失水,心葉無明顯受害癥狀;3 級,第1葉和第2葉中部出現脫水斑,第3葉葉緣嚴重失水,心葉輕微失水;4 級,第1葉和第2葉中部脫水斑連接成片,葉片萎蔫,第3葉中部始現脫水斑,心葉失水較明顯,但常溫下心葉尚能恢復;5 級,所有葉片嚴重失水萎蔫,幼苗在常溫下不能恢復。
于低溫弱光脅迫3、5、7 d后不同品種幼苗分別選取10株調查冷害癥狀,3次重復,按上述分級方法判定冷害級數,計算冷害指數。
冷害指數 =(1×S1+2×S2+3×S3+4×S4+5×S5+0×S0)/(低溫脅迫總株數×5)
式中S為每一冷害級的株數。
1.2.2 電解質滲漏率與丙二醛含量 分別于低溫脅迫0、7 d時,取不同品種幼苗3株,3次重復,選擇由上向下數第2片葉,打取直徑0.8 cm的葉圓片,放入具塞試管中,用美國產 ORION TDS 型電導率儀測定葉片的電導率,計算電解質滲漏率(EL)。
EL=(E1-E0)/(E2-E0)×100%
式中,E1:初電導率,E2:沸水30 min后電導率,E0:去離子水電導率。
低溫脅迫7 d后取不同品種幼苗3株,3次重復,將由上向下數第2片葉剪碎稱取0.5 g,采用硫代巴比妥酸(TBA)顯色法測定丙二醛(MDA)含量(李合生,2000)。
1.2.3 根系活力 低溫脅迫7 d后,取不同品種幼苗3株,3次重復,將根系均勻剪成1 cm的小段并稱取0.2 g至試管中,采用TTC法測定根系活力(李合生,2000)。
1.2.4 氣體交換參數 采用Ciras-3型光合儀(PPSystems公司,美國)測定脅迫0 d和3 d時不同砧木由上向下數第2片葉的凈光合速率(Pn)、胞間CO2濃度(Ci)、氣孔導度(Gs)和蒸騰速率(Tr),測定時PFD、CO2濃度和葉溫分別由儀器的可調光源、內置式 CO2供氣系統和溫度監控裝置調控。每個品種測定幼苗6株,3次重復。
1.2.5 幼苗生長量 在低溫弱光脅迫0 d和7 d時,分別選取不同品種幼苗10株,3次重復。用直尺測量株高及第1、2、3片葉葉柄到葉尖的長度,用游標卡尺測量莖粗,并按照龔建華和向軍(2001)的方法計算葉面積,將植株烘干至恒重后用稱重法稱量干物質量。
日增量=(低溫脅迫7 d的生長量指標-未經低溫處理的生長量指標)/7
1.2.6 隸屬函數值 參照宋洪元等(1998)模糊數學中隸屬函數的方法,計算參試材料耐冷性指標的隸屬函數值,并累加比較不同品種的耐冷性。
1.2.7 ABA含量 低溫脅迫0 d和1 d分別剪取不同品種幼苗由上向下數第2片葉和根系,用錫箔紙包好置于液氮中速凍,參照Andygene公司生產的酶聯免疫試劑盒(貨號:AD0021Pl)說明書測定葉片和根系中的ABA含量,每品種3次重復。
1.3 數據處理
試驗數據采用Microsoft Excel軟件進行處理,運用DPS軟件進行單因素方差分析,并運用Duncan檢驗法對顯著性差異(P<0.05)進行多重比較,利用SigmaPlot軟件作圖。
2 結果與分析
2.1 低溫弱光脅迫下不同砧木幼苗的冷害指數
從表2可以看出,隨著低溫弱光脅迫時間的延長,不同品種幼苗的冷害指數均逐漸增加,冷害加劇。津優35號的冷害指數除在脅迫3 d時顯著低于格頓金凱,其余時間均為最高,受冷害影響最大。常規砧木黑籽南瓜的冷害指數一直較低,低溫耐受性表現較好。
從表2還可以看出,整個脅迫過程中,博強105和金媽媽519白籽南瓜的低溫耐受性表現亦較好,二者的冷害指數與黑籽南瓜無顯著差異;其余砧木幼苗的低溫耐受性在脅迫5~7 d時均強于津優35號,但低于黑籽南瓜。結合圖1不同砧木幼苗的表現亦可知,低溫弱光脅迫7 d后,津優35號幼苗的受害程度最大,葉片嚴重發黃萎蔫,而其余砧木表現則均優于津優35號,受害程度較輕,其中冷害指數較低的黑籽南瓜、博強105和金媽媽519表現較好。
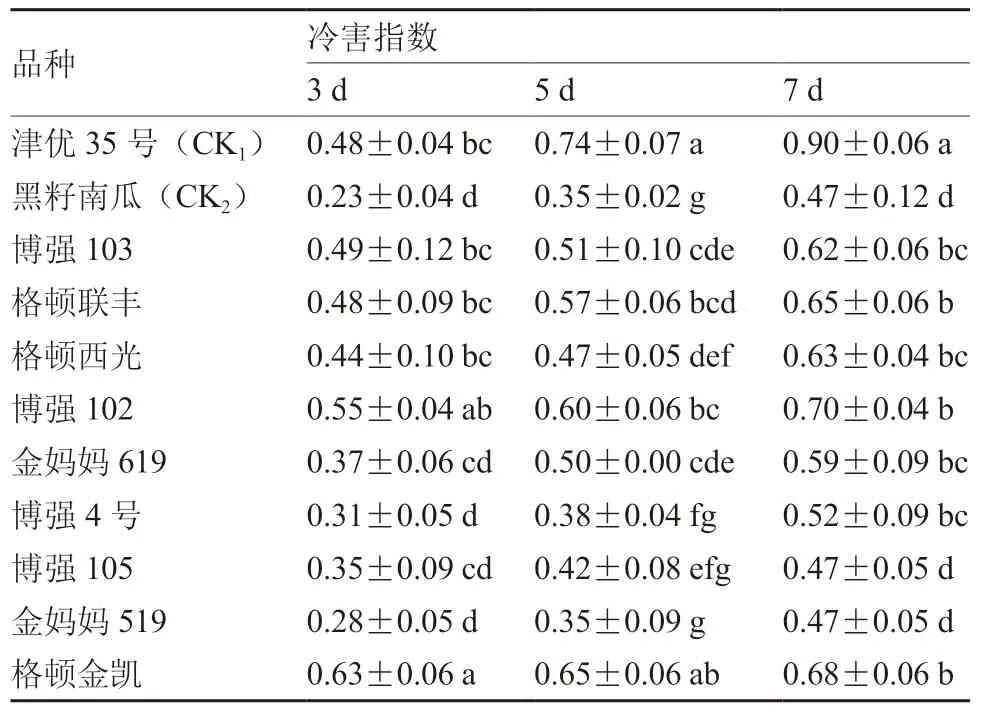
表2 低溫弱光脅迫下不同砧木幼苗的冷害指數

圖1 低溫弱光脅迫7 d后不同砧木幼苗的冷害癥狀
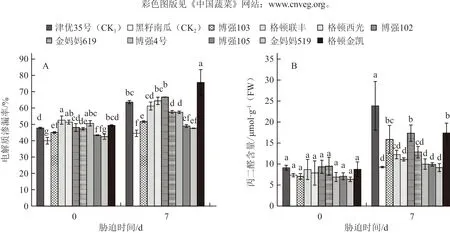
圖2 低溫弱光對不同砧木幼苗電解質滲漏率(A)和丙二醛含量(B)的影響
2.2 低溫弱光對不同砧木幼苗電解質滲漏率(EL)和丙二醛(MDA)含量的影響
圖2-A表明,低溫弱光脅迫7 d后,不同品種幼苗的電解質滲漏率均出現不同程度的上升,其中格頓金凱的電解質滲漏率與0 d相比增加了53.1個百分點,增加幅度最大;津優35號增加了33.1個百分點,而黑籽南瓜僅增加了11.4個百分點,博強105、金媽媽519分別增加了12.9和12.1個百分點,增幅較小。低溫弱光脅迫7 d后不同幼苗的電解質滲漏率從小到大依次為:黑籽南瓜<金媽媽519<博強105<博強103<博強4號<金媽媽619<格頓聯豐<格頓西光<津優35號<博強102<格頓金凱,說明砧木黑籽南瓜、博強105和金媽媽519幼苗受到的低溫傷害較輕。
低溫弱光脅迫后不同品種幼苗葉片中MDA含量的變化與電解質滲漏率的變化趨勢相似,即脅迫7 d后MDA含量均增加,津優35號的上升幅度較大,黑籽南瓜、博強105和金媽媽519的增加幅度較小(圖2-B)。
2.3 低溫弱光對不同砧木幼苗根系活力的影響
由圖3可以看出,不同品種在常溫條件下的根系活力差異較大。低溫脅迫前黑籽南瓜、博強105、金媽媽519根系活力較高,低溫脅迫7 d后除格頓聯豐外其余品種根系活力均有不同程度的升高,其中金媽媽519根系活力最高,黑籽南瓜、博強4號次之。
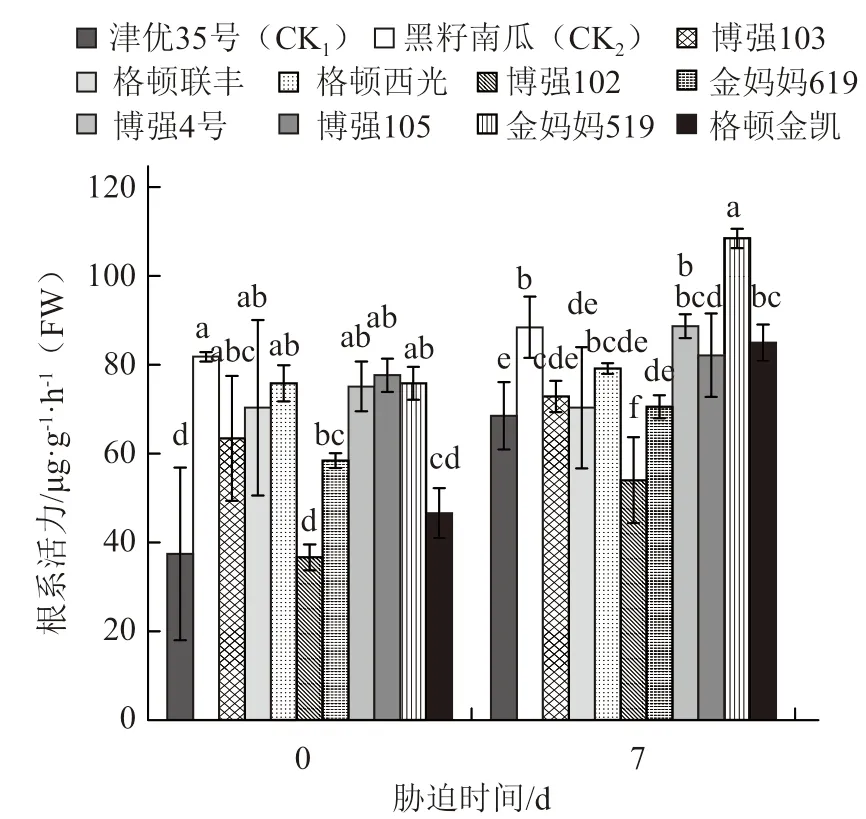
圖3 低溫弱光對不同砧木幼苗根系活力的影響
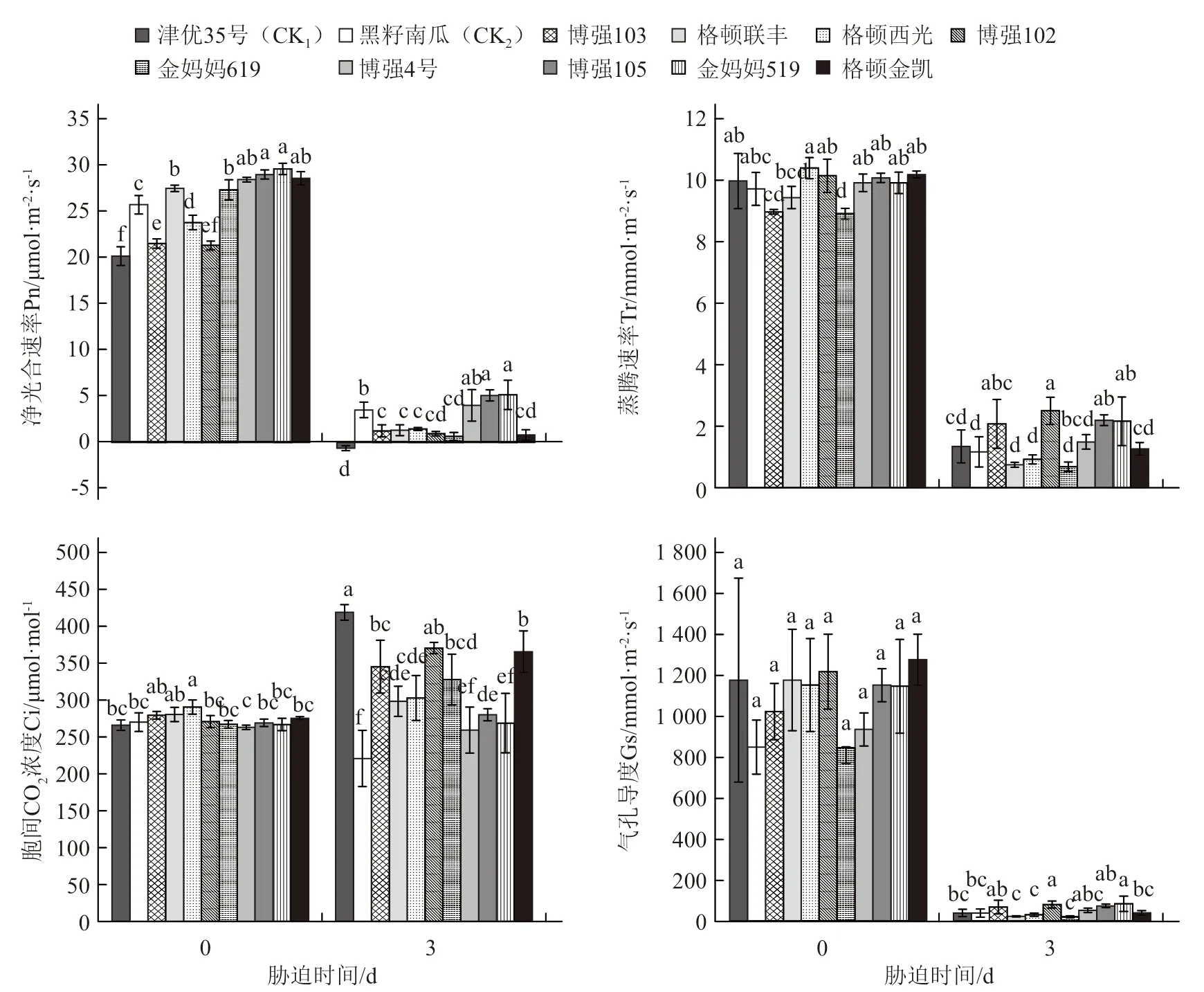
圖4 低溫弱光對不同砧木幼苗氣體交換參數的影響
2.4 低溫弱光對不同砧木幼苗氣體交換參數的影響
因監測的Pn在脅迫3 d時開始出現負值,因此圖4只顯示不同品種幼苗低溫弱光處理0 d和3 d的光合氣體交換參數。從圖4-A可以看出,脅迫前所有砧木幼苗葉片的Pn均明顯高于津優35號,以博強105和金媽媽519最高,甚至顯著高于黑籽南瓜;脅迫處理3 d后,砧木Pn明顯降低,但仍表現為所有南瓜砧木幼苗的Pn值均高于津優35號,其中金媽媽519和博強105的表現最好,亦顯著高于黑籽南瓜。
低溫脅迫前,各品種的Tr(圖4-B)、Ci(圖4-C)和Gs(圖4-D)多差異不顯著,而脅迫3 d后,不同品種幼苗的Tr和Gs均明顯降低而Ci除黑籽南瓜、博強4號、金媽媽519出現降低,其他品種均出現不同程度上升。與津優35號相比,南瓜砧木幼苗的Gs與Tr均較高或與津優35號差異不顯著,而Ci值均低于津優35號。
2.5 低溫弱光對不同砧木幼苗生長量的影響
從表3可以看出,黑籽南瓜、金媽媽519及博強105的株高、莖粗、葉面積和干物質量的日增量均表現較好,其中黑籽南瓜表現最好,株高、干物質量的日增量均顯著高于其他品種,莖粗、葉面積的日增量與博強105、金媽媽519差異不顯著。津優35號和博強102、格頓金凱表現較差,因受冷害較重,葉緣嚴重失水卷曲,葉面積的日增量甚至出現負值。
2.6 低溫弱光脅迫下不同砧木幼苗耐冷性綜合評價
如表4所示,各砧木幼苗的隸屬函數值均高于津優35號,其中金媽媽519最高,其次為博強105,且二者均高于黑籽南瓜;博強102和格頓金凱的隸屬函數值較低。說明白籽南瓜金媽媽519和博強105具有高于常規耐冷砧木黑籽南瓜的低溫耐受性,博強102和格頓金凱的低溫耐受性較弱。
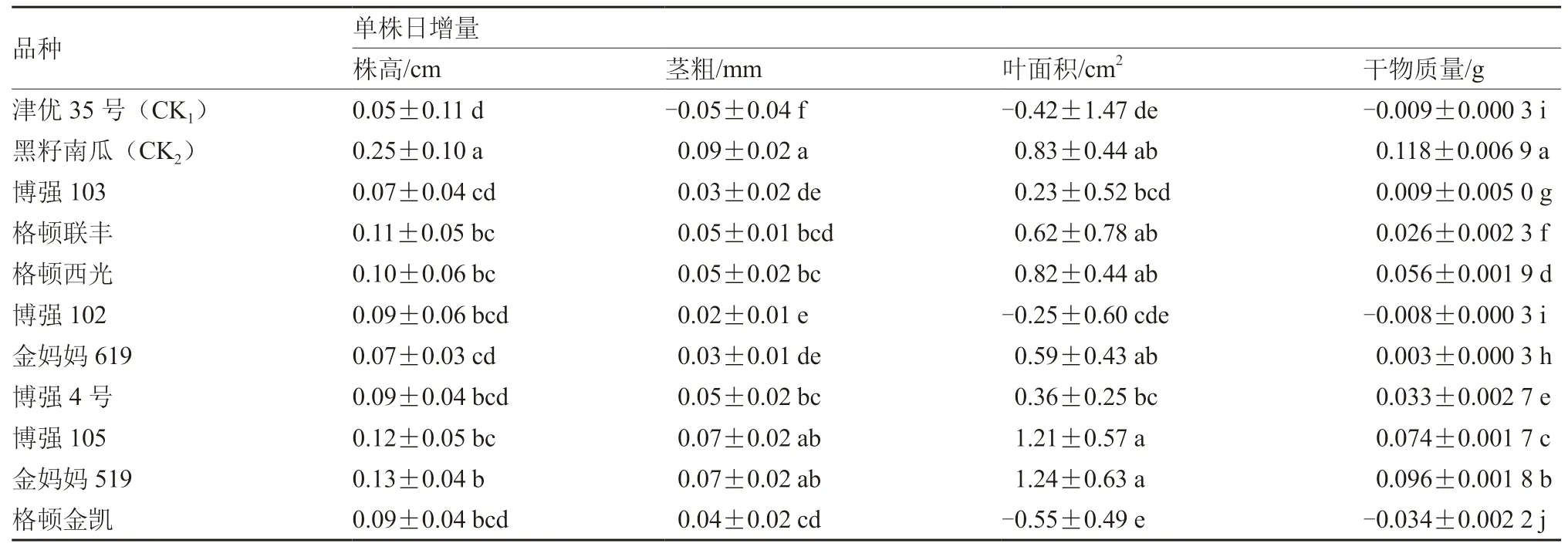
表3 低溫弱光對不同砧木幼苗生長量的影響
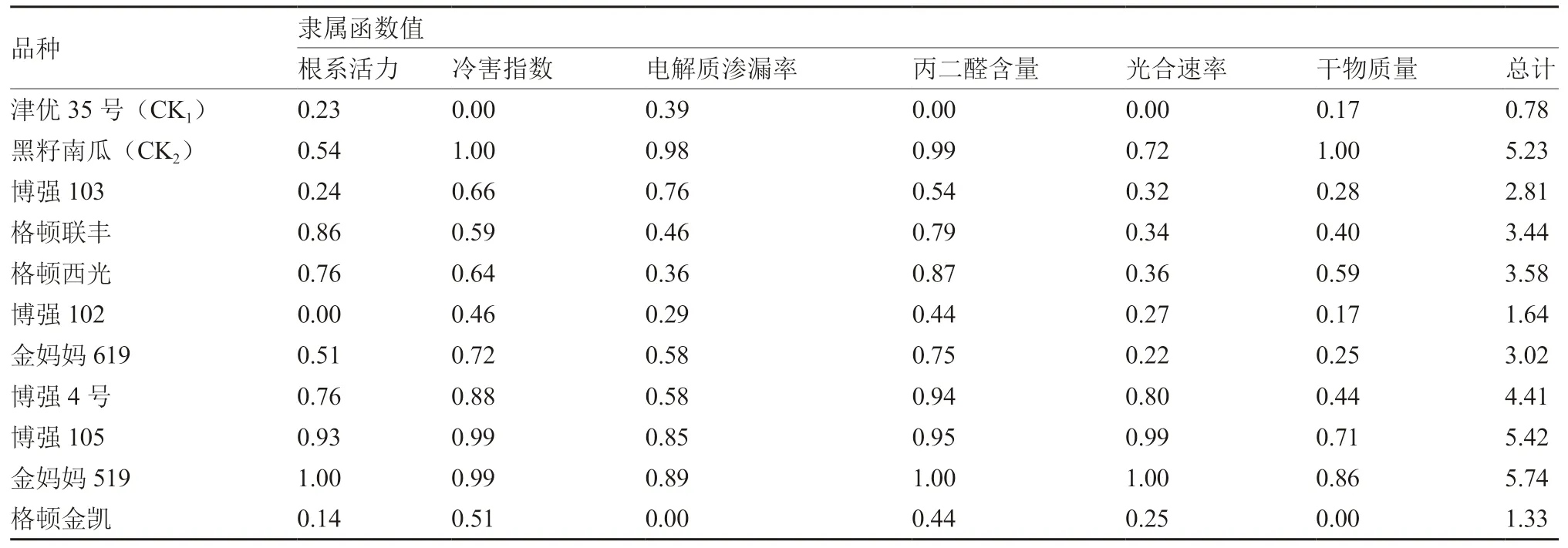
表4 低溫弱光脅迫下不同砧木幼苗耐冷性指標綜合評價
2.7 低溫弱光對不同砧木幼苗根系和葉片中ABA含量的影響
從圖5可以看出,幼苗經過1 d的低溫脅迫后根系和葉片ABA含量均有所增加,其中金媽媽519幼苗根系ABA含量最高,黑籽南瓜、博強105、博強4號幼苗根系的ABA含量也顯著高于其他品種;金媽媽519幼苗葉片中的ABA含量在常溫下雖然最低,但脅迫1 d后卻是最高的,其ABA增加量明顯高于其他砧木幼苗。對不同砧木根系和葉片中ABA增加量與砧木幼苗隸屬函數值進行相關性分析,得出相關系數r=0.765 8,說明不同砧木耐冷性的強弱與根系中及根系+葉片中ABA的增加量呈顯著正相關,但與葉片中ABA含量的增加量無顯著正相關性(圖6)。
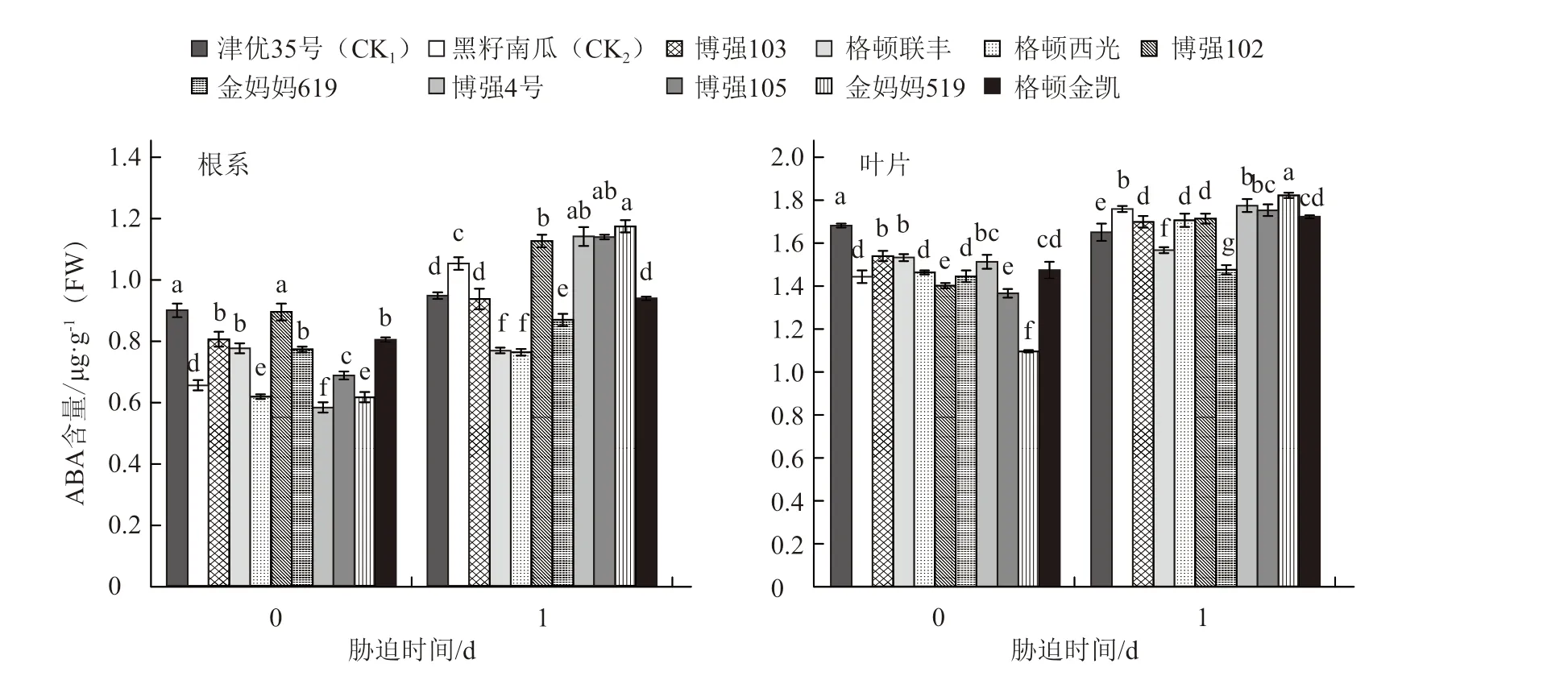
圖5 低溫弱光對不同砧木幼苗ABA含量的影響
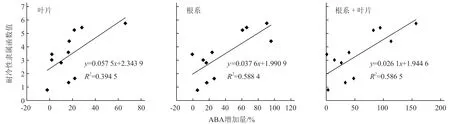
圖6 ABA增加量與耐冷性隸屬函數的相關性
3 結論與討論
本試驗中,隨著低溫弱光脅迫時間的延長,幼苗的冷害指數逐漸增加,受害程度加劇。電解質滲漏率和MDA 含量均可反映細胞膜脂過氧化程度及對非生物脅迫耐性的強弱(Chaoui et al.,1997;李天來和李益清,2008;馬樂元 等,2017)。本試驗中,低溫弱光脅迫7 d 后,所有幼苗的電解質滲漏率和MDA 含量均出現不同程度的上升,常規砧木黑籽南瓜的冷害指數、電解質滲漏率和MDA含量最小,且與金媽媽519 和博強105 無顯著差異;說明低溫弱光脅迫下黑籽南瓜、博強105、金媽媽519 的受傷害程度較低,對低溫弱光的耐受性強于其他砧木材料。
嫁接增強接穗抗性的主要機理是砧木具有發達的根系。韓敏等(2018)的研究也證實了砧木根系對嫁接苗耐冷性高低的貢獻遠大于接穗。本試驗中低溫脅迫0 d 時除博強102 砧木的根系活力與津優35號的根系活力無顯著差異外,其余材料的根系活力均顯著高于津優35號。前人研究指出,低溫脅迫會導致根系活力下降(周艷虹 等,2003;劉玉鳳 等,2017),但在本試驗中,低溫弱光脅迫7 d后根系活力多出現不同程度的增加,這可能是因為參試砧木幼苗是在水培條件下進行低溫處理的,這與王佩(2014)的研究結果一致,即水培條件會增強植物的根系活力,其中以金媽媽519的根系活力最高。
Schupp和Ferree(1990)研究指出,光合速率的變化與根系存在一定的聯系。本試驗中,低溫弱光脅迫3 d后不同品種幼苗的Pn、Tr和Gs明顯下降,津優35號的Pn甚至出現負值,但根系活力最高的金媽媽519的Pn亦是最高。這可能是因為根系活力較高有助于吸收更多的礦質元素,促進葉片中色素的合成(王元征 等,2011),有利于Pn增加,積累更多的光合產物,從而使得低溫弱光下金媽媽519的株高、莖粗、葉面積和干物質量的日增量均表現較好。
同時,從表4亦可以看出所有砧木的隸屬函數值均高于津優35號,其中金媽媽519最高,其次為博強105,且二者均高于常規耐冷砧木黑籽南瓜。于賢昌等(1999)研究指出,嫁接苗耐冷性的增加與ABA、IAA、GA等激素的變化有關;同樣,Li 等(2014)發現,以絲瓜為砧木的嫁接黃瓜高溫抗性的增加主要是因為砧木根系中ABA含量的積累。本試驗結果顯示,低溫弱光脅迫后砧木材料葉片和根系中ABA 含量均有所增加,且不同砧木耐冷性的高低與根系中及根系+葉片中ABA的增加量呈顯著正相關,說明砧木材料耐冷性的高低確與ABA含量的變化有關,可將其作為日后篩選耐冷砧木的一個重要參考指標,但其機理有待于進一步研究。
綜上分析可知,金媽媽519和博強105可選作新的黃瓜耐冷砧木,尤其是低溫弱光脅迫下金媽媽519仍具有很高的根系活力,更適合在黃瓜嫁接生產上推廣應用。
