棉花線粒體基因cRT-PCR改良及其在尋找CMS相關基因中的應用
2019-05-28 03:46:42王裴林周利利梁成真孟志剛郭三堆
生物技術進展 2019年3期
關鍵詞:差異
王裴林, 周利利, 梁成真, 孟志剛, 郭三堆, 張 銳
中國農業科學院生物技術研究所, 北京 100081
線粒體是細胞內氧化磷酸化和合成三磷酸腺苷(ATP)的主要場所,為細胞的活動提供了能量。細胞生命活動所需的能量95%來自線粒體,所以其又有“細胞動力工廠”之稱[1]。除了為細胞供能外,線粒體還參與諸如細胞分化、細胞信息傳遞和細胞凋亡等過程,并有調控細胞生長和細胞周期的能力。因此,線粒體基因組中基因的異常表達常常會影響細胞的能量代謝,從而引發一系列的表型變化,如抗病性降低或細胞質雄性不育的發生等。對線粒體基因組編碼基因的全長轉錄本進行分析,有利于深入研究細胞質基因的功能以及某些與細胞質關聯性狀的分子機理。
細胞質雄性不育(cytoplasmic male sterility,CMS)是由細胞質基因控制的并可被核恢復基因抑制的母性遺傳性狀。近幾年,棉花功能基因組學進展良好,但棉花細胞質雄性不育機理的相關研究報道不是很多,尤其是胞質不育的機理,尚缺乏足夠的分子生物學證據[1,2]。基于目前的研究進展,多數科學家認為CMS不育發生的原因大多是由于線粒體基因的重組重排導致,所以對線粒體基因全長轉錄本的獲得顯得尤為重要。
在傳統方法中,cDNA末端快速擴增(rapid amplification of cDNA ends, RACE)方法往往被用于精確定位RNA分子的5′或3′末端,但由于線粒體基因大部分不存在轉錄后加工的過程,即不會在轉錄后進行5′加帽子結構和3′端加polyA序列,也沒有內含子,而是采用邊轉錄邊翻譯的方式,所以不適用于RACE技術。為克服這一弊端,Kuhn和Binder于2002年提出環式反轉錄聚合酶鏈式反應(cyclic reverse transcription polymerase chain reaction, cRT-PCR),該技術極大簡化了操作步驟,成功實現一次性確定目的RNA的5′和3′末端的設想,具有明顯的高效與便捷性[3]。
cRT-PCR技術現已廣泛應該用于確定基因的轉錄起始位點和啟動子、鑒定核糖體RNA前體的5′和3′末端以及評估成熟mRNA的3′末端的長度和轉錄后延伸。目前,cRT-PCR技術應用于水稻和擬南芥等植物的細胞基因研究,但研究報道中均是利用該方法對特異基因進行研究[4,5],尚未見從轉錄組的水平獲得全部線粒體基因全長轉錄本的應用。
目前,根據諸多的文獻報道,CMS基因通常有以下五種特點:①不同的CMS之間通常不具備同源性;②CMS的位點經常出現在ATP基因的上下游;③它們往往通過重組產生嵌合的ORF;④CMS編碼的蛋白通常具有細胞毒性;⑤CMS編碼蛋白通常會形成跨膜結構域。尤其是嵌合ORF的形成是不育基因目前的重點研究方向,通過cRT-PCR技術可以較為簡單的獲得靶標基因的UTR區,為尋找嵌合的ORF提供了一個新的方向[6~8]。
近幾年棉花功能基因組學進展良好,但棉花細胞質雄性不育機理的相關研究報道不是很多,同其他作物雄性不育機理研究相比有所滯后。因此,我們希望通過該方法的改良及應用能為尋找不育基因帶來新的方法。本研究在獲得高質量RNA的基礎上,對RNA進行高效環化,并通過隨機引物對線粒體全基因組基因進行cRT-PCR,獲得了全部線粒體基因的全長轉錄本,并以ATP6基因為例,比較了利用特異引物和隨機引物進行cRT-PCR后所獲得的全長轉錄本的差異。棉花細胞質雄性不育的研究對于更好地利用棉花雜種優勢,以及加深對植物不育機理的認識,具有重要的理論和現實意義。
1 材料與方法
1.1 材料與試劑
1.1.1實驗材料 本實驗所用材料為中國農業科學院抗蟲棉課題組實驗室培育的不育系P30A。
1.1.2試劑 原平皓RNA提取試劑盒、RNA酶抑制劑、DNA酶、Superstar max超保真mix購自Genestar公司;反轉錄試劑盒購自全式金生物科技有限公司;T4 RNA連接酶購自Promega公司;DNA瓊脂糖凝膠回收試劑盒購自Axygen生化科技有限公司;T載體、感受態購自擎科新業生物技術有限公司;高溫滅菌冷卻至室溫的DEPC水。
1.2 實驗方法
1.2.1引物設計 ①RT-PCR引物使用本實驗室發表的PrimerServer: PCR Primers Batch Design & Specificity Check引物設計網站[9],對參考基因序列設計普通引物,引物位置在參考序列兩邊留出100~300 bp為宜,并由網站檢測為特異引物。
②cRT-PCR引物由RT-PCR引物反向互補得到,估計其大小,約為原始序列兩邊預留的200 bp加上可能的UTR區域(約100~600 bp)(表1)。
1.2.2RNA提取 按照原平皓試劑盒RNA提取說明書提取RNA,并在加入去蛋白液前增加DNA酶,去除DNA,為后續實驗去除DNA污染。提取完成后通過瓊脂糖凝膠電泳檢測RNA的完整度;并通過Nano-drop對RNA的濃度和純度進行測量。

表1 4類線粒體基因cRT-PCR引物信息Table1 Four gene families mitochondrial gene cRT-PCR primer information.
1.2.3RNA環化 體系:10 μg總RNA,1 μL RNA酶抑制劑,2 μL T4 RNA連接酶Buffer,1 μL T4 RNA連接酶,1 μL ATP, DEPC水補足至20 μL。將混合好的樣品置于金屬浴上,16℃連接過夜。在連接液的每管中加入10 μL 氯仿并用移液器反復吹打;加入100 μL無水乙醇移液器吹打混勻,冰浴15 min;將混合物12 000 r/min,10 min離心,棄上清;最后,加入14 μL DEPC水溶解[11,12]。
1.2.4反轉錄 按照全式金生物科技有限公司反轉錄試劑盒對產物進行反轉錄,完成后用Nano測定濃度,ddH2O稀釋至終濃度200 ng/μL ,凍存備用。
1.2.5cRT-PCR程序 我們在棉花線粒體基因的主要4種類型的基因ATP、COX、RPS、NAD基因中隨機選擇7個作為實驗對象,并根據普通PCR引物反向互補鏈設計相應的cRT-PCR引物,其特點是僅能以環化RNA為模板獲得擴增條帶。擴增體系:25 μL Superstar max超保真mix(Genestar),1 μL cDNA,正反向引物各1 μL,ddH2O 22 μL。擴增程序:94℃預變性5 min;94℃變性30 s;54℃(取決于引物)退火30 s,72℃延伸40 s,35~40個循環;72℃延伸10 min,4℃保存。完成PCR之后,將PCR產物于1%瓊脂糖凝膠中上樣,120 V,30 min。電泳結束后,按照DNA瓊脂糖凝膠回收試劑盒(Axygen生化科技有限公司)回收條帶。
1.2.6ATP6的測序分析 為進一步分析測序結果的準確性,從上述7個棉花線粒體基因中隨機選取ATP6為代表進行分析。對ATP6基因的cRT-PCR引物進行擴增,然后將該序列回收后連接T載體,挑單克隆進行測序,最終挑選出一個一致性最好的克隆子進行分析,并與NCBI上的序列進行比對。然后采用隨機引物和特異引物共同對其進行cRT-PCR(圖1)。
2 結果與分析
2.1 線粒體基因cRT-PCR電泳分析
對棉花線粒體基因的主要4種類型的基因中隨機選擇的7個基因的擴增結果顯示,7個線粒體基因的電泳條帶清晰(圖2),表明RNA模板已經環化,為后續的實驗分析提供必要的前提。

圖1 隨機引物和特異引物ATP6全長轉錄本對比Fig.1 Comparison of random primer and specific primer ofATP6gene full-length transcript.
我們對棉花線粒體的33編碼蛋白基因(ATP1、ATP4、ATP6、ATP8、ATP9;COX1、COX2、COX3;RPL2、RPL5、RPL10、RPL16;NAD3、NAD4、NAD4L、NAD6、NAD7、NAD9;RPS3、RPS4、RPS7、RPS10、RPS12、RPS14;CCMB、CCMC、CCMFC、CCMFN;SDH3、SDH4;MATR、COB、MTTB)進行了cRT-PCR,發現大多數的基因UTR都在300~500 bp左右。我們使用特異引物和隨機引物同時進行擴增,發現兩者所獲得結果基本保持一致,通過測序驗證,僅有個別基因出現了極少數的SNP。
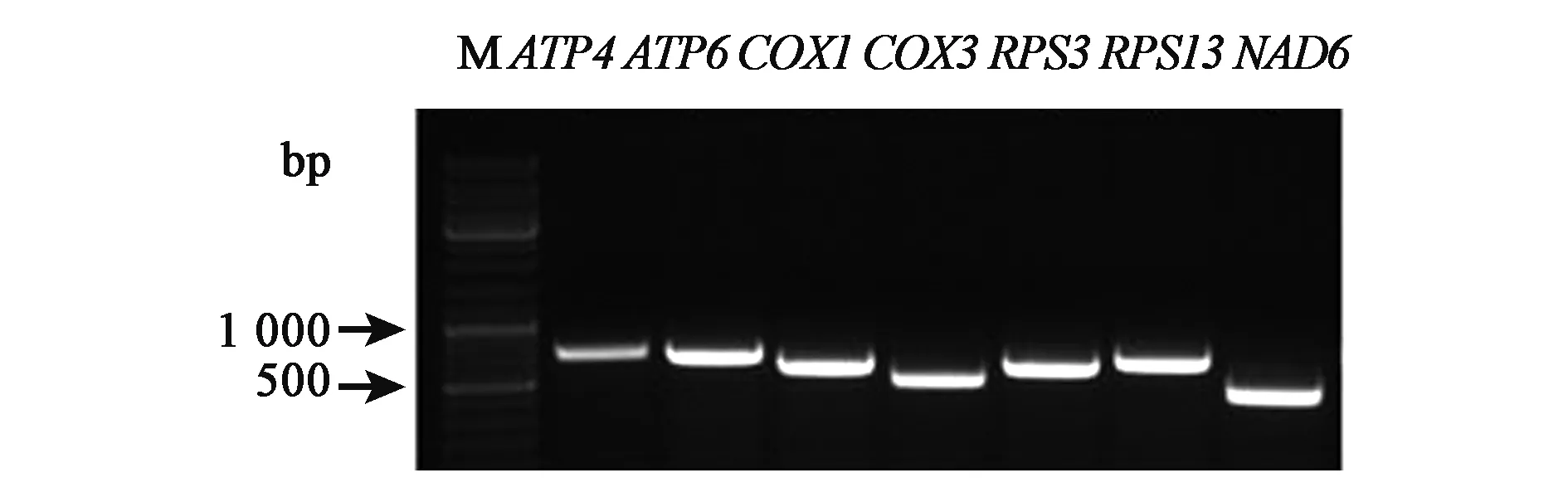
圖2 線粒體基因cRT-PCR電泳分析Fig.2 Analysis of mitochondrial gene by cRT-PCR.
2.2 ATP6基因測序結果分析
通過ATP6基因的cRT-PCR引物進行擴增,獲得一段全長567 bp的序列(圖3),測序并與NCBI上的序列進行比對,我們獲得了284 bp的5′UTR和22 bp的3′UTR,并且原基因上的一段重復區域也完全匹配,獲得了ATP6的完整全長轉錄本。測序結果與參考序列保持一致,之后,采用隨機引物和特異引物共同對其進行cRT-PCR,發現兩者獲得條帶一致。測序后,兩者序列僅出現了一個SNP。其余基因驗證測序結果同樣保持一致,所以我們認為采用隨機引物替代靶基因特異引物的方法在保證了正確率的情況下,更為方便快速。
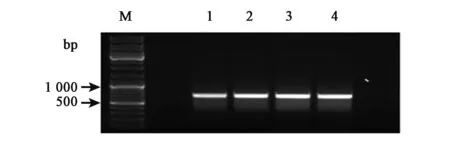
圖3 ATP6基因cRT-PCR擴增結果Fig.3 cRT-PCR results ofATP6gene.注:M:Marker;1,2:隨機引物;3,4:特異引物。
綜上所述,通過隨機引物進行反轉錄cRT-PCR同特異引物一樣能獲得目的基因的全長轉錄本,但在比對序列時也發現極少部分基因的測序結果存在基因的不完整性和突變性,主要表現為基因兩端堿基缺失與單核苷酸多態性。前者的發生取決于RNA的完整度以及環化效率,而單核苷酸多態性的發生則多半是由于特異引物和隨機引物的差異造成的。因此,在方便快捷的基礎上,首先采用隨機引物對大批量的基因進行cRT-PCR,如若所獲基因轉錄本的序列異常,為保證準確性,可輔助設計特異引物進行反轉錄。
2.3 cRT-PCR在尋找不育基因中的應用
通過對本課題組繁育的不育系和保持系同時進行線粒體基因的批量cRT-PCR,根據條帶大小以及測序結果對線粒體編碼蛋白基因進行考量,找到一些具有差異的候選基因,認為可能是嵌合ORF的插入導致了結果的差異。
如圖4所示,差異表達出現了四種情況,一是不育系和保持系條帶完全一致;二是不育系和保持系條帶存在明暗亮度差異;三是不育系有而保持系沒有條帶;四是在不育系和保持系中條帶大小不一致的情況。
另外有的基因在電泳條帶上表現一致,但通過測序出現了幾個氨基酸的缺失,或是出現一些SNP的情況,結果如表2所示: 線粒體編碼蛋白基因中ATP1、ATP4、ATP6、COX1在不育系和保持系的擴增中出現了明顯差異,尤其是ATP1、ATP6、COX1,在兩段序列中出現了大段的插入或者缺失,我們認為這些差異片段可能是由于線粒體的某種重組或者重排造成,或可能引起作物的不育。除此之外,線粒體的其他基因ATP8、COX2、NAD3等出現了在不育系和保持系中序列有SNP的情況,可能是由于PCR或者測序的原因,也有可能是線粒體的某種突變引起,但這種方式引起不育的可能性過低。
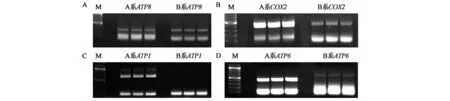
圖4 線粒體基因在不育系與保持系中的擴增條帶Fig.4 Amplification bands of mitochondrial genes in sterile line and maintainer line.注:A:ATP8;B:COX2;C:ATP1;D:ATP6;M:Marker;A系:不育系;B系:保持系。

基因名稱差異情況基因名稱差異情況ATP1A系260 bp,B系沒有RPS3SNPATP4A系:5′-AACGCGTTTAGGTAAGACTT-3′B系:5′-AAC--G---A-------CTT-3′RPS4RPS7-ATP6A系360 bp,B系526 bp,SNPRPS10SNPATP8SNPRPS12-ATP9-RPS14SNPCOX1A系B系161 bp序列差異,SNPCCMB-COX2SNPCCMCSNPCOX3-CCMFC-NAD3SNPCCMFNSNPNAD4-SDH3SNPNAD4LSNPSDH4未獲條帶NAD6-MATRSNPNAD7SNPCOB未獲條帶NAD9SNPMTTBSNP
3 討論
長期以來,cRT-PCR技術已廣泛應用于擬南芥、水稻等植物的細胞研究中,但鮮有涉及棉花細胞線粒體基因的研究。在本研究中,我們對轉錄本序列分析發現,cRT-PCR技術適用于獲得棉花線粒體目的基因的全長轉錄本,為后續技術改良奠定了實驗基礎。在傳統cRT-PCR技術的基礎上,我們優化了RNA提取方法和環化連接體系,顯著提高了環化效率。反轉錄階段,在保證與特異引物獲得相同完整轉錄本的基礎上,創新性地采用隨機引物進行大批量的基因檢測,操作更方便快捷。
基于本研究中改良的cRT-PCR技術存在上述提及的基因的不完整性和突變性現象,本研究提出如下針對性措施:首先保證RNA提取方法的科學準確性,鑒于RNA的降解起始于5′和3′末端,直接決定環化效率并影響擴增中基因序列的完整性,因此反應試劑中應保證一定濃度的RNA酶抑制劑,以從源頭上確保基因序列的準確性;其次,我們發現環化的效率極大的影響了擴增的結果,無論是特異引物還是隨機引物,都會出現區段缺失的情況,但在必要時將隨機引物改為傳統的特異性引物,以從反應途徑上確保降低單核苷酸多態性的發生,從而提升基因序列的特異性。在本研究中,需要綜合考慮影響實驗的諸多環節,其中需要格外關注的是RNA環化連接的質量。過夜可以為充分環化提供必要的時間保障,但鑒于單鏈RNA的易降解性,保證充分環化時間的前提是確保所加入的RNA酶抑制劑達到一定濃度,能夠保障RNA在環化期間的穩定性。此外,在過夜后環化效率依然不高的特殊情形下,加入適量ATP可以明顯加速RNA環化,提高環化效率。
我們通過改良的cRT-PCR技術對不育基因進行探索,通過對不育系和保持系的擴增,發現在ATP和COX家族中存在UTR區有差異的基因(在其他線粒體基因中大多是SNP的差異)。在此,我們暫不討論條帶明暗的差異,這可能和擴增效率或是拷貝數相關。差異基因中有無的差異、條帶大小的差異,都很可能是由于線粒體內部重排或者ORF的插入導致的,而這些差異可能會引起作物的CMS。根據本實驗結果,發現線粒體編碼蛋白基因中ATP1、ATP4、ATP6、COX1在不育系和保持系的擴增中出現了明顯差異,我們認為這些差異片段可能是由于某種重組或者插入造成,或可能引起作物的不育。UTR區段的差異傾向于ATP和COX基因,這和目前發表的CMS相關文獻保持一致,提升了作為CMS相關基因的可能性,因此我們認為該方法用來尋找不育基因是可行的。當然,這些差異的出現不一定是CMS基因造成的嵌合ORF,具體結果如何還需要進一步的實驗驗證。但是,通過cRT-PCR技術,為尋找CMS基因提供了新的方法和思路。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
