銅對意大利蜜蜂工蜂生理機能的影響
2019-05-27 08:16:56趙曉冬王紅芳胥保華
中國蜂業 2019年4期
趙曉冬 王紅芳 胥保華
(山東農業大學動物科技學院,泰安 271018)
銅是動物機體內必需的微量元素,其影響機體的代謝和生長發育、免疫功能,其不僅通過構成機體內的含銅酶(如超氧化物歧化酶、銅藍蛋白等)參與機體代謝過程,而且參與機體造血、自由基防御等活動[1]。蜂蜜是蜜蜂在自然狀態下主要的能量食物,含有多種礦物質。蜂蜜中常見的微量元素包括K、Na、Mg、Zn、Fe 和Cu 等20 多種,常量元素約占所有礦物質的98%~99%[2]。Bounias 等研究了有機銅源葡萄糖酸銅和乳酸銅對蜜蜂蜂螨防治的效果,發現葡萄糖酸銅效果更佳[3]。
銅參與機體的抗氧化功能主要是通過影響含銅氧化酶的活性而發揮作用的。銅是銅鋅超氧化物歧化酶的重要組成部分,可以催化機體代謝的有毒物質轉化為氧自由基和H2O2。蜜蜂中腸位于前胃之后,是消化和吸收的器官。中腸腸腔內有多層的圍食膜,可保護中腸細胞減少磨損,使中腸消化酶穿過圍食膜消化食物,再由中腸細胞吸收分解的營養物。在分子水平,低濃度的Cu2+可以食物中銅對昆蟲生長發育有重要作用,銅水平對棕尾別麻蠅的影響得出:飼料中添加低濃度的銅能夠提高棕尾別麻蠅的生理功能,而高濃度的銅則會產生抑制作用[4]。在蜜蜂人工飼料中添加銅的有機鹽——葡萄糖酸銅可以提高蜂群的抗病能力,但是銅的適宜添加水平及其對蜜蜂生理機能的影響還有待進一步研究證明。
1 材料與方法
1.1 試驗材料
試驗動物為意大利蜜蜂,取自蜂群健康、群勢基本相同的蜂群。試驗用葡萄糖酸銅(山東西亞化學有限公司),貨號為L103479-25g,純度為98%。
1.2 意大利蜜蜂工蜂幼蟲飼糧配制
試驗采用6 種不同銅水平的人工幼蟲飼糧飼喂意大利蜜蜂工蜂,6 種飼糧分別標記為A、B、C、D、E、F,飼糧組成見表1。
1.3 飼養管理
試驗于2017年在山東農業大學動物科技學院完成,供試蜂場為山東農業大學南校區試驗蜂場,蜜蜂在30℃和50%±5%R.H.的恒溫恒濕培養箱內進行飼養。在飼養過程中保證培養箱和飼喂盒的衛生,防止其他不利因素對蜜蜂產生的影響,飼養至蜜蜂全部死亡。分別于9日齡測定體成分、血淋巴生化指標、抗氧化指標、相關基因的相對表達量、制作中腸切片,并測定壽命。
1.4 測定指標及相關方法
1.4.1 離群意大利蜜蜂壽命的測定
在23cm×9.5cm×5cm 蜂盒中飼養,并置于培養箱中,調節恒溫恒濕培養箱的溫度為30℃,濕度為50%±5%,每天定時記錄死亡的蜜蜂數,直至蜜蜂全部死亡。
1.4.2 離群意大利蜜蜂工蜂中腸切片的制作
選用組織石蠟包埋切片。
1.4.3 離群意大利蜜蜂蜂體銅含量的測定
從每試驗組取9日齡的蜜蜂30 只,每10 只蜜蜂為一個重復。采用硝酸-雙氧水消解,ICP-ms 測定銅含量。
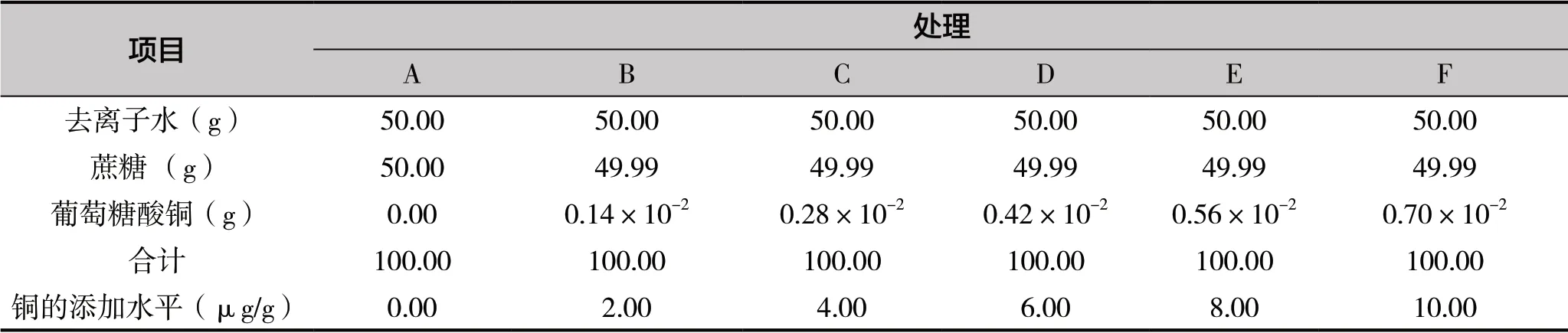
表1 飼養離群意大利蜜蜂飼糧組成及銅水平
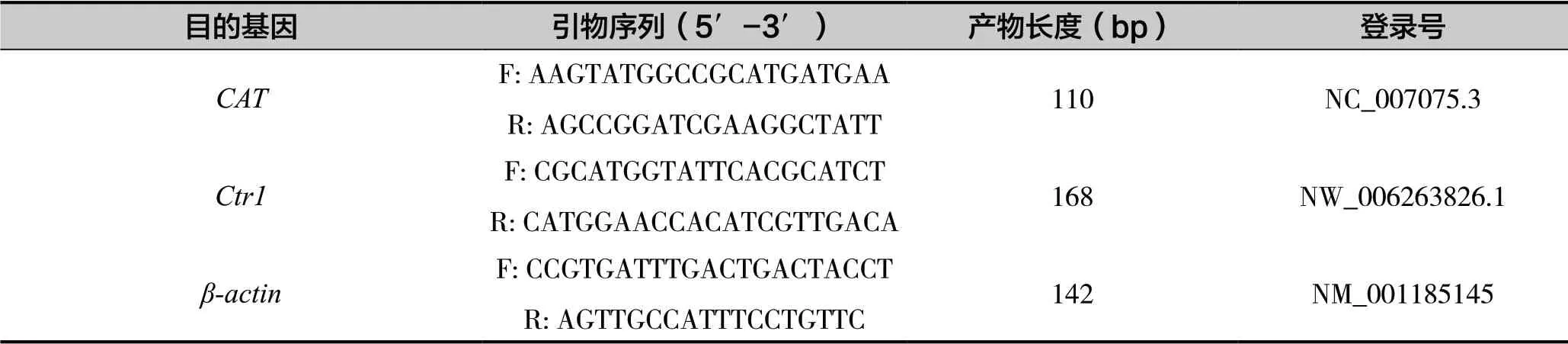
表2 試驗中PCR引物
1.4.4 血淋巴生化指標的測定
采用體積為20μl 的毛細管吸取9日齡蜜蜂血淋巴50μl 于加有苯基硫脲的1.5ml 離心管中,于-80℃進行保存。測定時,4℃條件下3000r/min離心10min,取上清備用,采用日立7020 型全自動生化分析儀測定蜂體血淋巴中總蛋白(TP)、總膽固醇(TCHO)。
1.4.5 蜜蜂中抗氧化酶活性的測定
從各試驗組9日齡蜜蜂分別取出12 只蜜蜂,每3 只蜜蜂為一個重復,使用SOD 試劑盒A007(南京建成生物工程研究所)測定SOD 活性。利用可見光分光光度計(UV-2000)測定吸光度并計算蜂體抗氧化能力。
1.4.6 工蜂幼蟲蟲體溶菌酶、防衛素1 基因相對表達量的測定
采用Trizol 法提取總RNA,使用反轉錄試劑盒(TaKaRa)將提取的總RNA 樣品反轉錄為cDNA,-20℃保存備用。目的基因引物設計參考序列來自于NCBI 數據庫,采用Primer 5.0 進行引物設計,以β-肌動蛋白(β-actin)為內參基因,委托生工生物科技有限公司合成引物,引物序列如表2所示。
1.5 數據處理與分析
采用SAS 9.2 軟件對數據進行單因素方差分析和Duncan 氏法多重比較,P<0.01 表示差異極顯著,P<0.05 表示差異顯著。
2 結果
2.1 銅對蜜蜂壽命的影響
從蜜蜂存活率曲線圖來看,整個飼養期間,對照組A、處理組B、C 存活率明顯高于處理組D、E、F,10 天內,各處理組的存活率差異很小,自11 天起,處理組B 存活率明顯高于其他組(見圖1)。
2.2 銅對蜜蜂中腸圍食膜的影響
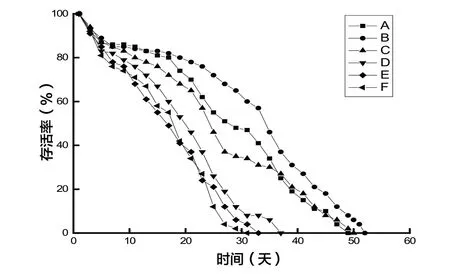
圖1 銅對蜜蜂存活率的影響
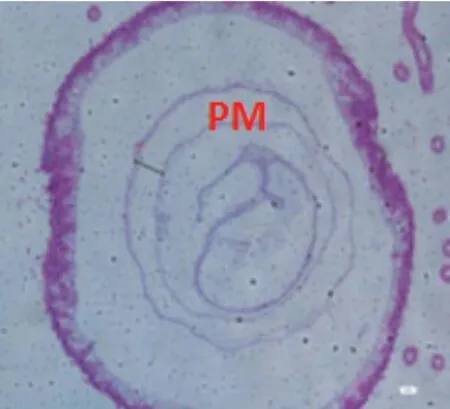
圖2-1 A組蜜蜂中腸組織切片,H&E染色,100×
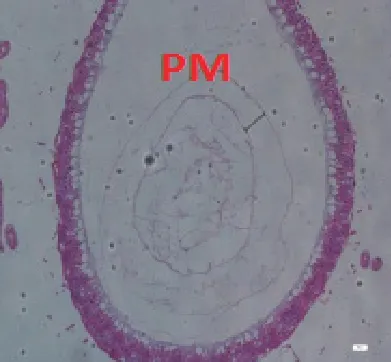
圖2-2 B組蜜蜂中腸組織切片,H&E染色,100×
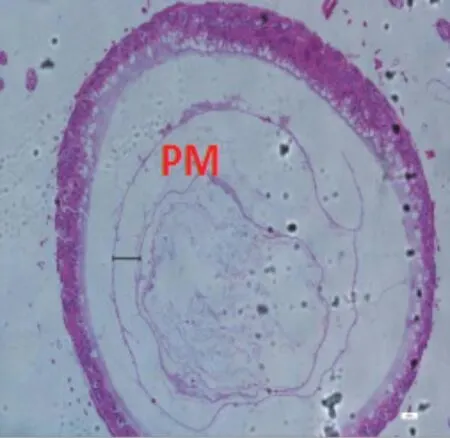
圖2-3 C組蜜蜂中腸組織切片,H&E染色,100×

圖2-4 D組蜜蜂中腸組織切片,H&E染色,100×
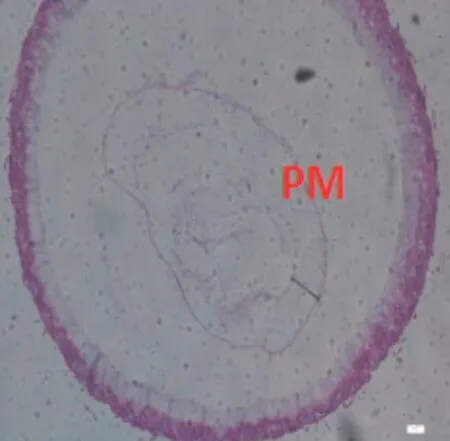
圖2-5 E組蜜蜂中腸組織切片,H&E染色,100×
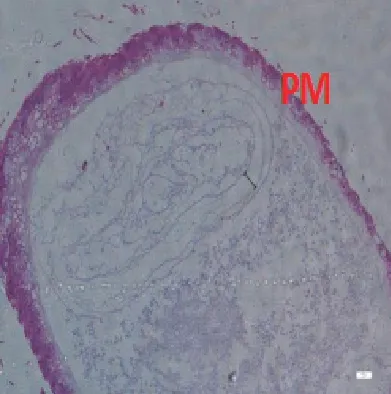
圖2-6 F組蜜蜂中腸組織切片,H&E染色,100×
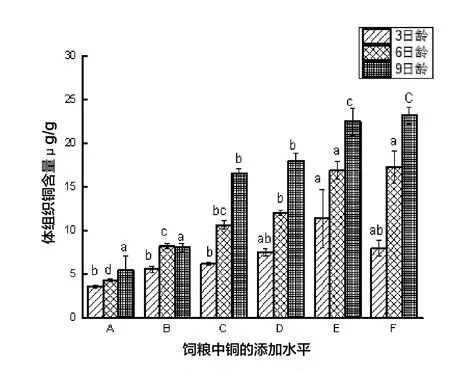
圖3 離群蜜蜂體組織銅含量
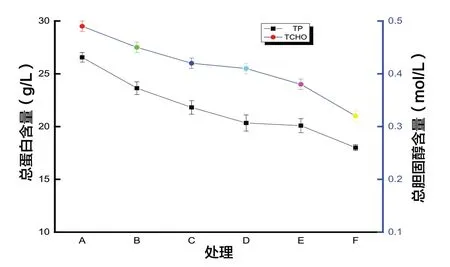
圖4 飼糧中銅水平對9日齡蜜蜂血淋巴生化性能的影響
9日齡中腸切片顯示(100×)(見圖2),飼糧中銅的不同添加水平影響離群蜜蜂中腸圍食膜的結構。對照組A、處理組B、C 蜜蜂中腸的PM 較厚且形態勻稱,而B 處理組PM 最厚;D、E、F 處理組蜜蜂中腸PM 的厚度不一。當飼糧中銅水平為2.00μg/g 時對蜜蜂中腸圍食膜的結構相對來說較為完整。
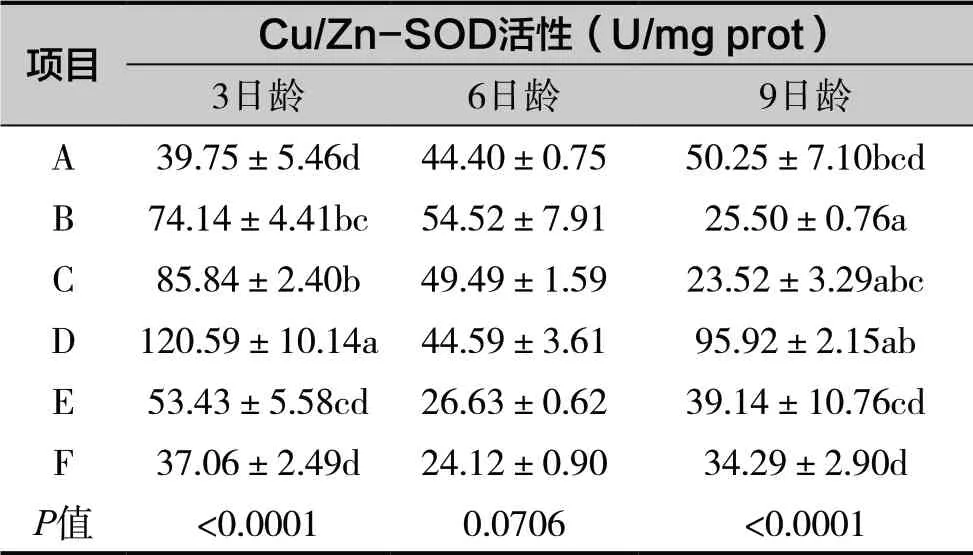
表3 銅對離群蜜蜂Cu/Zn-SOD活性的影響
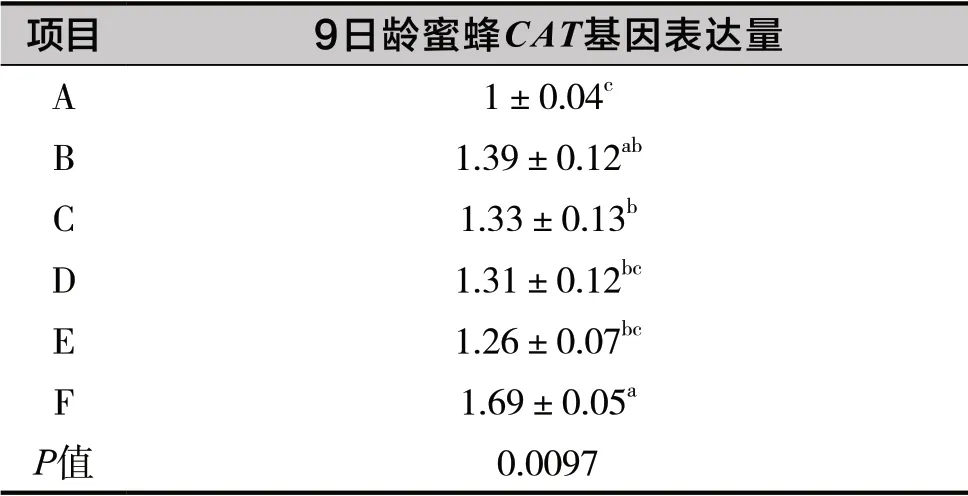
表4 銅對蜜蜂CAT相對表達量的影響
2.3 銅對蜜蜂體組織銅含量的影響
由圖3可知,3、6、9日齡體組織Cu 含量也隨飼糧中Cu 水平不斷增加而顯著上升(P<0.05),對照組蜜蜂體組織Cu 含量顯著小于其他組(P<0.05)。
2.4 銅對蜜蜂血淋巴生化指標的影響
由圖4可知,飼糧中銅的添加可顯著影響蜜蜂體中總蛋白、總膽固醇含量(P<0.05),隨著飼糧中銅添加水平的升高,9日齡蜜蜂體中總蛋白、總膽固醇含量均下降(P<0.05)。
2.5 銅對蜜蜂抗氧化能力及相關基因相對表達量的影響
2.5.1 銅對蜜蜂抗氧化能力的影響
由表3可知,D 處理組3日齡蜜蜂Cu/Zn-SOD活性顯著高于其他組(P<0.05),B 處理組6日齡蜜蜂Cu/Zn-SOD 活性顯著高于對照組A 組以及其他處理組(P<0.05),3、6日齡蜜蜂Cu/Zn-SOD 活性都呈現先升高后下降的趨勢。
2.5.2 銅對蜜蜂抗氧化相關基因相對表達量的影響
由表4可知,銅顯著影響9日齡蜜蜂CAT基因表達量(P<0.05)。隨著幼蟲飼糧中銅水平的不斷升高,9日齡蜜蜂幼蟲CAT基因相對表達量呈現逐漸增加的趨勢。
2.5.3 銅對蜜蜂高親和力銅吸收蛋白基因相對表達量的影響
由圖5可知,9日齡幼蟲F 處理組Ctr1基因的相對表達量顯著高于其他組(P<0.01),隨著幼蟲飼糧中銅水平的不斷增加,9日齡蜜蜂Cu/Zn-SOD基因相對表達量呈現逐漸增加的趨勢(P<0.05)。
3 討論
3.1 飼糧中銅的添加水平影響蜜蜂生長發育
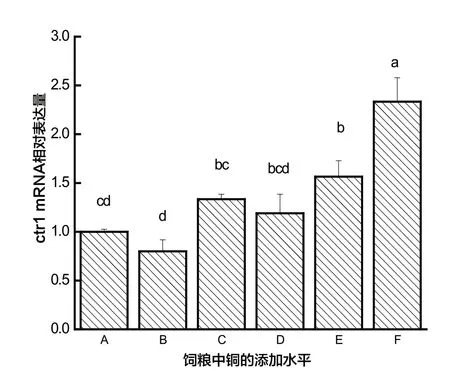
圖5 銅對9日齡蜜蜂Ctr1相對表達量的影響
Adam 等[4]研究表明,Cu2+主要沉積在雙翅目幼蟲中腸上皮細胞的細胞漿內。過量的Cu2+使中腸上皮細胞發生變性、腫脹、破裂[5]。本實驗中,在飼糧中添加銅能夠保護腸道,有助于腸道的生長發育,當蜜蜂飼糧糖漿中銅添加量在2.00μg/g 時能夠使中腸圍食膜的結構較為均勻完整,有利于提高蜜蜂的健康水平。昆蟲取食含Cu2+的飼料后,Cu2+在體內的積累量與飼料中Cu2+濃度存在顯著的劑量效應關系,并隨著取食時間的延長而增加[6]。在昆蟲體內存在毒物興奮效應,即低濃度刺激、高濃度抑制的劑量反應現象[7]。本研究中高劑量的Cu2+脅迫會顯著降低昆蟲的總蛋白和總脂濃度。
3.2 飼糧中銅的添加水平影響蜜蜂的抗氧化能力
在昆蟲體內存在抗氧化酶系統,如超氧化物歧化酶(SOD)、過氧化氫酶(CAT)和過氧化物酶(POD)等[8]。重金屬銅可與酶分子中的其他金屬離子發生競爭性替代作用而抑制酶活力,使昆蟲體內生化反應緩慢,并減弱昆蟲清除氧自由基能力,最終抑制昆蟲代謝和生長發育,延長其發育歷期。綜合本試驗結果表明,飼糧中銅的添加水平為2.00~4.00μg/g時,意大利蜜蜂工蜂的抗氧化狀態最好。
3.3 飼糧中銅的添加水平影響銅離子轉運蛋白基因相對表達量
銅離子轉運蛋白家族主要包括銅離子轉運蛋白(Ctr)和銅離子轉運磷酸化ATP 酶(Cu-ATPase)。Ctr1的表達水平與細胞內銅離子濃度有密切關系,其表達量的多少直接對機體銅代謝產生影響,銅缺乏時可促進該蛋白的表達,過量則抑制其表達。本研究表明銅的添加對于蜂體Ctr1的表達有顯著影響,其相對表達量在銅的添加水平0.00~10.00μg/g 的范圍內有升高的趨勢,然而目前關于飼糧中銅水平對于其代謝相關酶的影響尚未見報道。
4 結論
(1)當飼糧中銅的添加水平為2.00μg/g 時能顯著提高意大利蜜蜂工蜂的壽命,且對蜜蜂中腸食膜的結構最為有利。
(2)當飼糧中銅的添加水平在0.00~10.00μg/g 的范圍內不斷升高時,9日齡蜜蜂體組織銅含量顯著升高,銅對9日齡意大利蜜蜂過氧化氫酶基因相對表達量、高親和力銅轉運蛋白基因相對表達量顯著增加,但蜜蜂血淋巴中總蛋白和總膽固醇含量顯著降低。
(3) 6日齡和9日齡蜜蜂蜂體超氧化物歧化酶(Cu/Zn-SOD)活性隨著銅的添加水平的提高出現先升高后降低的趨勢,都在2.00μg/g 時活性最高。
(4) 綜合考慮以上指標,推薦意大利蜜蜂工蜂飼糧的適宜銅的添加水平為2.00~4.00μg/g。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國火炬(2010年8期)2010-07-25 11:34:30
