牛種布魯氏菌2308強毒株dsbG基因缺失突變株的構建與初步評價
2019-05-23 03:57:203
中國人獸共患病學報 2019年5期
3
布魯氏菌病是一種由布魯氏菌(Brucella)引起的流行范圍廣的人獸共患傳染病之一,是《中華人民共和國傳染病防疫法》規定的乙類傳染病[1]。人類感染布魯氏菌病主要表現為波狀熱、盜汗、關節炎,甚至導致不孕不育等;公畜感染易引起睪丸炎,懷孕母畜感染易導致流產[2]。布魯氏菌病在世界各地都有發生,給畜牧業發展和人類健康帶來了嚴重威脅。因此,對該病的研究受世界各國學者的高度重視[3]。
布魯氏菌是兼性胞內寄生的革蘭氏陰性菌,呈球形或短桿狀,菌體長0.6~1.5 μm,寬0.5~0.7 μm,其主要寄生于巨噬細胞和胚胎滋養層細胞[4-5]。近年來,對布魯氏菌毒力因子的研究已成為國內外學者研究的重點,布魯氏菌無經典的毒力因子,其毒力與侵入宿主細胞、抵抗宿主細胞殺傷作用以及胞內生殖有關[6]。
DsbG是最早顯示具有一般分子伴侶活性的周質蛋白之一,這種陪伴蛋白質折疊的能力可能增加DsbG作為蛋白質二硫鍵異構酶的有效性[7]。本研究小組前期利用蛋白質組學研究發現;DsbG在布魯氏菌毒力株2308和疫苗株RB51之間的表達存在差異,因此,本研究以布魯氏菌2308毒力株的dsbG基因為基礎,以布魯氏菌2308作為研究對象,利用抗性替代構建差異表達蛋白DsbG的基因突變株,研究其遺傳穩定性和生長狀態,并研究其在HPT-8細胞中的存活和繁殖能力,為探索布魯氏菌潛在毒力因子奠定基礎,為進一步揭示布魯氏菌感染與懷孕母畜流產之間的分子機制提供理論依據。
1 材料和方法
1.1 材 料
1.1.1菌株和細胞 牛種布魯氏菌2308毒力株購自中國獸醫藥品監察所;RB51疫苗株由中國農業大學吳清民教授惠贈;pUC19K載體由北京軍事醫學科學院惠贈;人胚胎滋養層細胞(HPT-8)由石河子大學動物科技學院人獸共患傳染病實驗室保存;大腸埃希菌(Escherichiacoli, DH5a)、pMD19-T simple載體購自TaKaRa公司。
1.1.2試劑 DMEM培養基、胎牛血清購自Gibco公司;硫酸慶大霉素購自河南輔仁懷慶制藥有限公司;0.22 μmol/L濾器購自美國Milipore公司;細胞培養板購自COSTAR公司;BrucellaBroth培養基和BrucellaAgar培養基購自美國BD公司;質粒小提取試劑盒和dNTP購自Tiangen Biochemical Technology Co., Ltd.;其他試劑均為國產分析純。
1.2 方 法
1.2.1引物設計與合成 利用Primer Premier 5.0軟件設計相應引物,送華大基因合成,引物序列見表1。
表1 引物和序列Tab.1 Primer and sequences
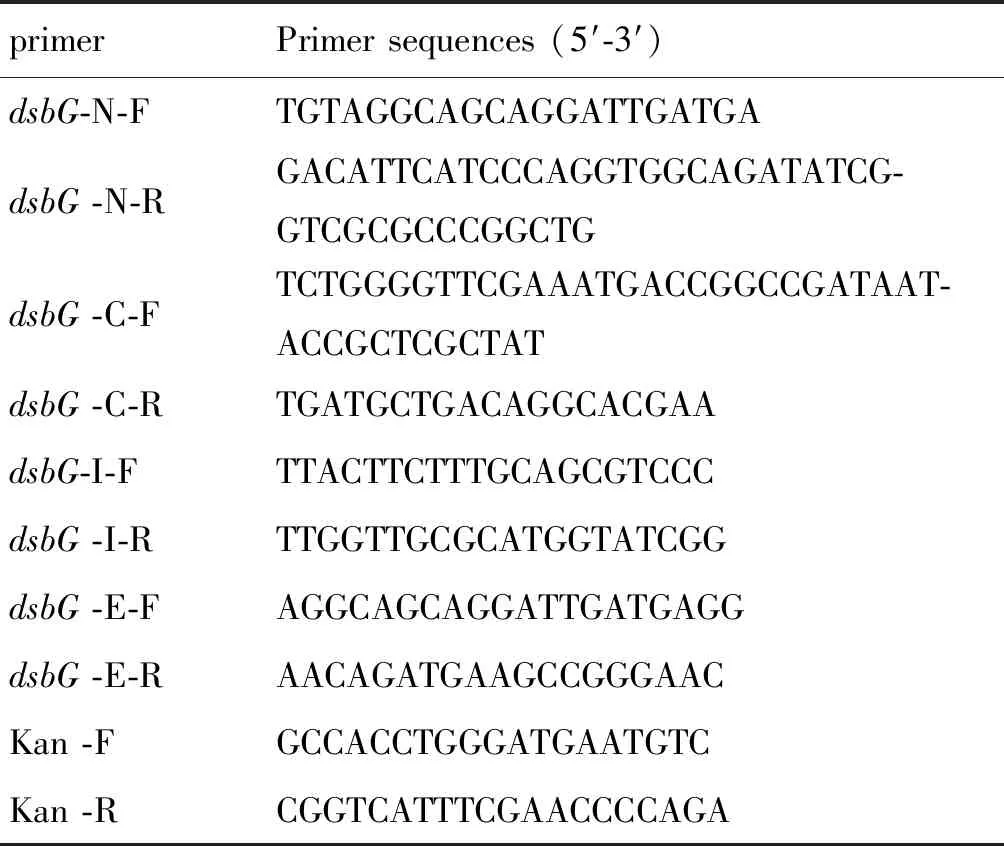
primerPrimer sequences (5′-3′)dsbG-N-FTGTAGGCAGCAGGATTGATGAdsbG -N-RGACATTCATCCCAGGTGGCAGATATCG-GTCGCGCCCGGCTGdsbG -C-FTCTGGGGTTCGAAATGACCGGCCGATAAT-ACCGCTCGCTATdsbG -C-RTGATGCTGACAGGCACGAAdsbG-I-FTTACTTCTTTGCAGCGTCCCdsbG -I-RTTGGTTGCGCATGGTATCGGdsbG -E-FAGGCAGCAGGATTGATGAGGdsbG -E-RAACAGATGAAGCCGGGAACKan -FGCCACCTGGGATGAATGTCKan -RCGGTCATTTCGAACCCCAGA
1.2.2布魯氏菌dsbG突變株自殺載體的構建 以牛種布魯氏菌2308的DNA為模板,以dsbG-N-F和dsbG-N-R為引物擴增dsbG基因的N端同源臂和以dsbG-C-F和dsbG-C-R為引物擴增其C端同源臂,以pUC19K質粒為模板,以Kan-F和Kan-R作為引物擴增卡那霉素抗性片段。通過融合PCR技術,對dsbG基因的N端同源臂、卡那霉素抗性片段和C端同源臂進行融合擴增。回收融合片段產物并連接到pMD19-T載體上,轉化到DH5a感受態細胞中,通過PCR鑒定,并送生工生物工程股份有限公司進行測序。
1.2.3布魯氏菌dsbG突變株的構建 將質粒pMD-DK-Kan電轉化到2308感受態細胞中,活化后涂于帶卡那霉素抗性的布魯氏菌固體培養基,于37 ℃恒溫培養箱中倒置培養48~72 h,挑取單個細菌,分別用卡那霉素替換目的基因的內、外部鑒定引物進行PCR鑒定,獲得2308ΔdsbG突變株。
1.2.4布魯氏菌2308ΔdsbG突變株的遺傳穩定性檢測 將初步篩選獲得的布魯氏菌2308ΔdsbG突變株反復傳代培養10代后,通過內、外部檢測引物對2308ΔdsbG突變株進行PCR鑒定和遺傳穩定性檢測。回收外部引物獲得的基因片段送至華大基因測序。
1.2.5布魯氏菌親本株和突變株生長曲線測定 布魯氏菌2308、RB51和2308ΔdsbG突變株在相同起始濃度下,于37 ℃,170 r/min條件下培養。間隔2 h取樣1次,檢測OD600的變化,直到細菌進入平臺期,并繪制2308、RB51和2308ΔdsbG突變株的生長曲線。
1.2.6布魯氏菌侵染HPT-8細胞形態學觀察 分別用對數生長期的布魯氏菌2308、RB51和2308ΔdsbG突變株按200∶1感染復數侵染HPT-8細胞,24 h后棄去培養液,用PBS漂洗3次,并與含有50 μg/mL慶大霉素的液體培養基共孵育60 min,以殺死胞外的布魯氏菌,用倒置顯微鏡觀察細胞的形態學變化。
1.2.7布魯氏菌粘附侵襲實驗 分別使用布魯氏菌2308、RB51和2308ΔdsbG突變株按200∶1感染復數侵染HPT-8細胞,并在5% CO237 ℃環境中共孵育10 min、20 min、40 min、60 min、90 min和120 min后,用PBS漂洗3次,除去未粘附的細菌,再與含有50 μg/mL慶大霉素的DMEM培養液共孵育60 min以殺死胞外菌。用0.2% Tritonx-100裂解細胞,稀釋后涂于布魯氏菌固體培養基。3~5 d后,通過計算CFU,比較2308、RB51和2308ΔdsbG突變株的粘附侵襲能力。
? E.H.Gombrich,“ An early seventeenth-century canon of artistic excellence:Pierleone Casella's elogia illustrium artificum of 1606”,Journal of the Warburg and Courtauld Institutes,vol.50,1987,pp.224-32.
1.2.8布魯氏菌胞內繁殖實驗 分別用布魯氏菌2308、RB51和2308ΔdsbG突變株按200∶1的感染復數侵染HPT-8細胞,60 min后,用慶大霉素殺死胞外菌,分別在0 h、4 h、8 h、12 h和24 h棄去培養液,PBS漂洗3次。用0.2% Tritonx-100裂解細胞,稀釋后涂于布魯氏菌固體培養基,3~5 d后,通過計算CFU,比較并分析布魯氏菌2308、RB51和2308ΔdsbG突變株的胞內繁殖能力。
1.2.9統計學分析 利用SPSS Statistics 17.0分析軟件對實驗數據進行統計學分析,P<0.05為有統計學意義。
2 結 果
2.1布魯氏菌dsbG基因突變株的構建
2.1.1dsbG基因同源臂及卡那霉素抗性基因的擴增 以金屬浴滅活的布魯氏菌2308為模板,PCR擴增dsbG基因的上、下游同源臂序列,分別獲得約為500 bp(圖1A)和399 bp(圖1B)的片段;以pUC19K質粒為模板,擴增卡那霉素抗性片段,獲得大小約為1 093 bp的片段(圖1C);獲得條帶大小與預期值相符。
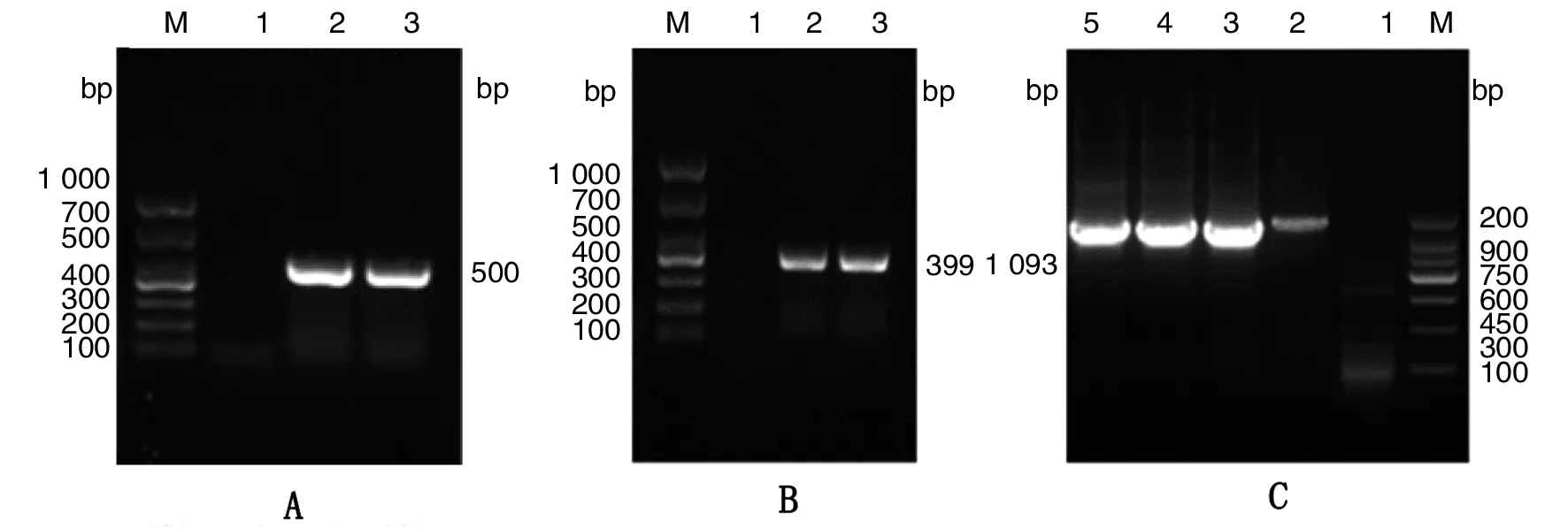
A: M,DNA marker;1,陰性對照; 2~3,dsbG-N PCR產物B: M,DNA marker;1,陰性對照; 2~3,dsbG-N PCR產物C: M,DNA marker;1,陰性對照; 2~5,Kan PCR產物圖1 dsbG基因同源臂及卡那霉素抗性基因的PCR擴增Fig.1 PCR amplification of the dsbG gene homology arm and kanamycin resistance gene
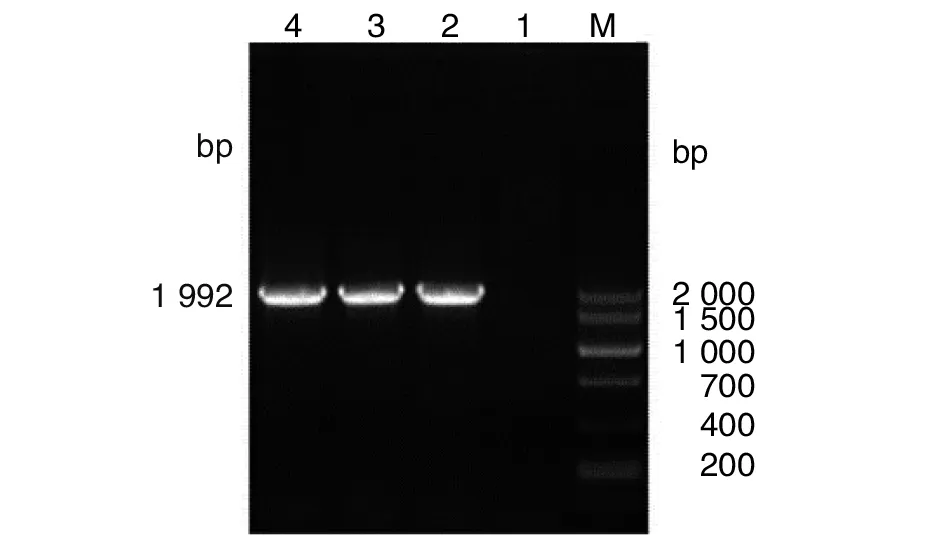
2.1.2dsbG基因同源臂和卡那霉素抗性基因的融合 通過融合PCR技術將上述3個基因片段進行融合擴增,獲得大小約為1 992 bp的基因片段,其與dsbG基因上下游同源臂片段和卡那霉素抗性基因片段預期疊加相符(圖2)。
2.1.3布魯氏菌dsbG突變株載體的構建及鑒定 將pMD19-T載體連接到融合片段上,并轉化至DH5a感受態細胞中,獲得同源重組載體pMD19-T-ΔdsbG-K(1 992 bp), PCR驗證結果與疊加片段大小相同(圖3)。

M: DNA marker; 1~3: dsbG-N-K-C 融合PCR產物圖2 dsbG-N-K-C融合PCR擴增 Fig.2 dsbG-N-K-C fusion PCR amplification
M: DNA marker;1: 陰性對照;2~4:pMD19-T-ΔdsbG-Kan PCR產物圖3 同源重組載體pMD19-T-ΔdsbG-Kan 的菌液PCR鑒定Fig.3 Identification of pMD19-T-ΔdsbG-Kan was amplified via PCR
2.2布魯氏菌2308ΔdsbG基因突變株的初步篩選及鑒定 使用dsbG-N-F和Kan-R鑒定引物分別對2308ΔdsbG突變株和2308親本株進行擴增,結果顯示: 2308ΔdsbG突變株可擴增出預期大小相符(1 593 bp)的特異片段,而2308親本株擴增為陰性,如圖4所示:表明抗性基因正確替代了dsbG基因。
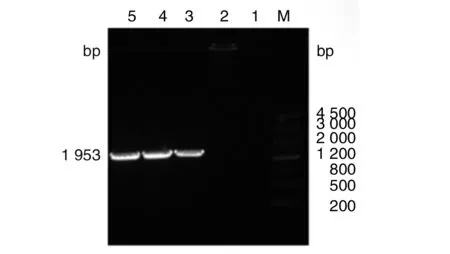
M: DNA marker;1: 陰性對照;2: 2308對照;3~5:檢測菌液圖4 2308ΔdsbG的篩選鑒定Fig.4 Screening and identification of 2308ΔdsbG
表2 檢測引物及片段長度
Tab.2 Detection primer and fragment length

檢測引物敲除前大小/bp敲除后大小/bpdsbG internal detec-tion primerdsbG-I-FdsbG-I-R7530dsbG external detec-tion primerdsbG-E-FdsbG-E-R1 4001 740
結果顯示:構建的基因缺失株在缺失前后所擴增的片段與理論值相符,外部引物獲得基因經測序后比對序列,同源性為100%,表明成功獲得了2308ΔdsbG基因缺失株,并且在連續培養10代后未觀察到回復現象,即2308ΔdsbG突變株可進行穩定遺傳,如圖所示(5A、B)。
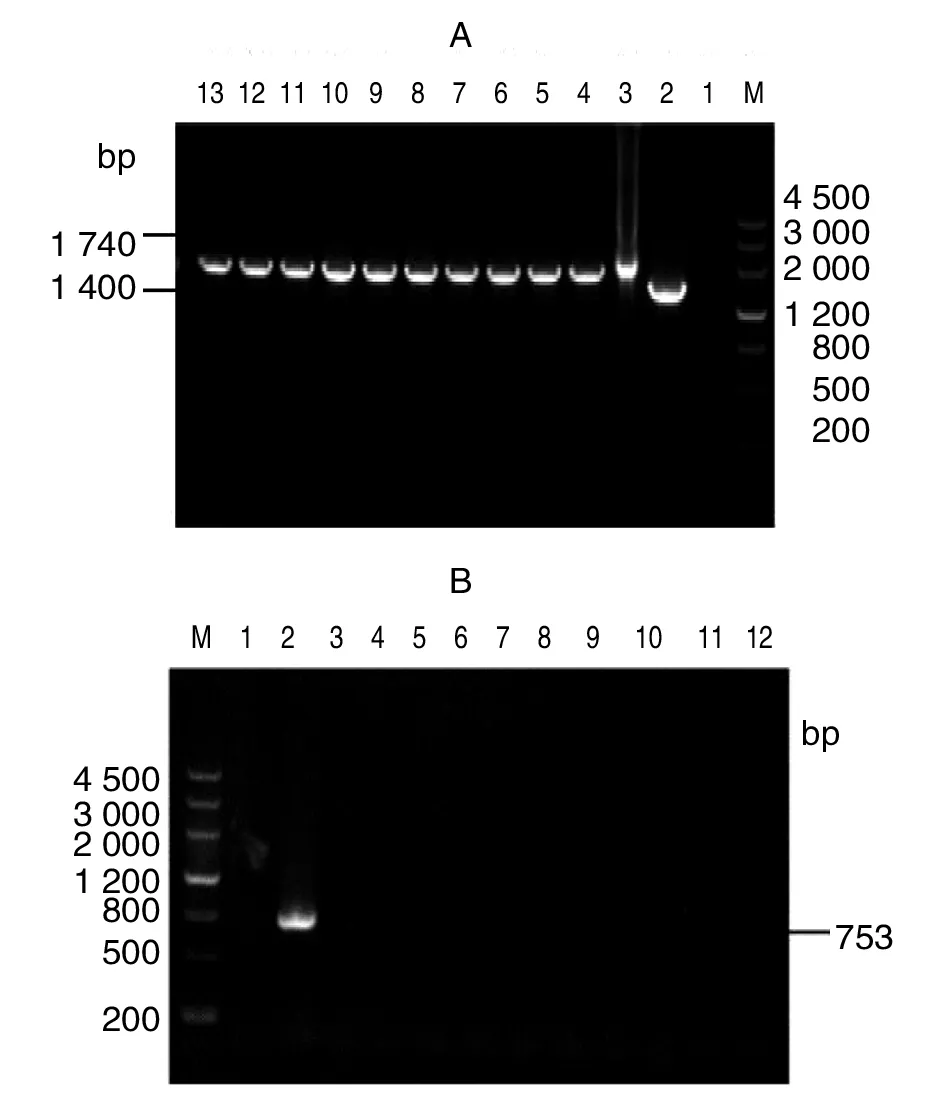
A:M,DNA maker; 1,陰性對照; 2,S2308; 3,PMD19-T-ΔdsbG-Kan 質粒對照;4~13,S2308ΔdsbG基因突變株B:1,陰性對照; 2,S2308; M,DNA maker; 3~12,S2308ΔdsbG基因突變株圖5 布魯氏菌S2308ΔdsbG的遺傳穩定性檢測Fig.5 Genetic stability detection of Brucella S2308ΔdsbG
2.4突變株和親本株的生長曲線分析 繪制布魯氏菌2308、RB51和2308ΔdsbG突變株的生長曲線(圖6)。從生長曲線可以看出,這些菌株的生長趨勢相似,并且同一時間點菌液濃度僅存在細微差異。
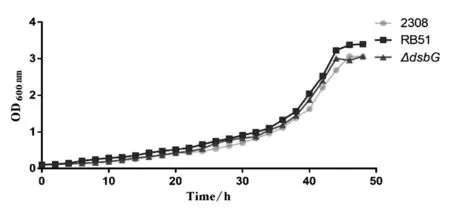
圖6 布魯氏菌生長曲線Fig.6 Growth curve of Brucella strains
2.5胚胎滋養層細胞形態學觀察 布魯氏菌2308、 RB51和2308ΔdsbG突變株侵染HTP-8細胞后,使用倒置顯微鏡觀察侵染前后的細胞形態變化。如圖7所示:我們觀察到正常細胞呈多邊形,培養48 h后成片狀或簇狀生長,用布魯氏菌2308侵染后的細胞呈圓形、皺縮現象較為嚴重,間隙變寬,有少量的細胞脫落和碎片。然而用RB51和2308ΔdsbG突變株侵染后的細胞易出現聚合現象,邊界模糊,并伴有輕微脫落。
2.6布魯氏菌的粘附侵襲能力 不同時間點的布魯氏菌2308、RB51和2308ΔdsbG突變株侵入HPT-8細胞的數量有所差異。如圖8所示:隨著時間延長,進入細胞的布魯氏菌數量逐漸增加,各菌株約在60 min后侵入細胞的數量接近飽和不再增加。2308ΔdsbG突變株在每個時間點的粘附侵襲能力顯著低于親本株2308和疫苗株RB51(F=1.201,P<0.05)。

A:2308組;B:RB51組;C:2308ΔdsbG組;D:PBS組圖7 胚胎滋養層細胞侵染前后的變化Fig.7 Change of the trophoblast cell infected with different Brucella

注:*代表P<0.05圖8 布魯氏菌對胚胎滋養層細胞的粘附侵襲能力Fig.8 Invasive ability of Brucella in HPT-8
2.7布魯氏菌的胞內繁殖能力 布魯氏菌2308、RB51和2308ΔdsbG突變株在HPT-8細胞內具有不同的生存復制能力。如圖9所示:侵染后各時間點2308ΔdsbG突變株的胞內存活能力顯著低于親本株2308和疫苗株RB51(F=8.627,P<0.05)。
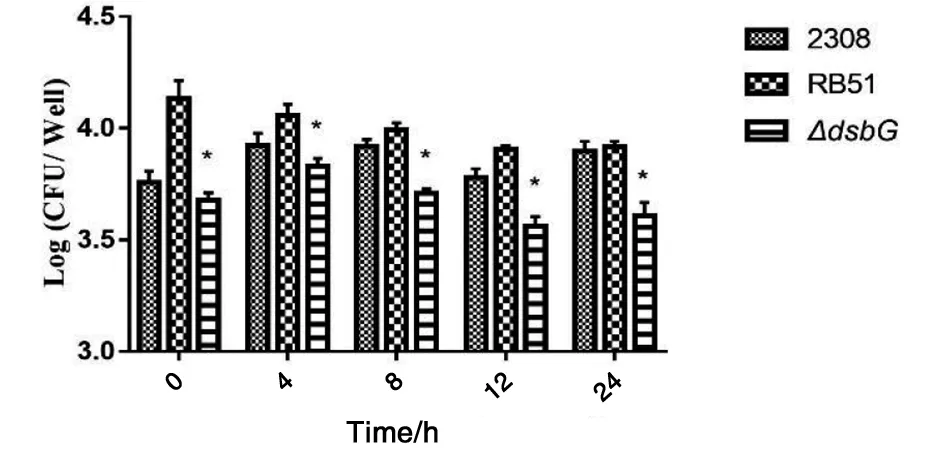
注:*代表P<0.05圖9 布魯氏菌的胞內存活能力Fig.9 Intracellular survival ability of Brucella
3 討 論
布魯氏菌雖無典型的毒力因子,如外毒素、質粒、菌毛、莢膜等,但它具有較強的致病性,特別是對懷孕母畜,常引起懷孕母畜流產[8]。主要原因是布魯氏菌對懷孕母畜胎盤具有較強的親嗜性,胎盤中的赤蘚醇有助于布魯氏菌生長,而布魯氏菌能夠侵入胚胎滋養層細胞,胚胎滋養層細胞是母體和胎兒聯系的紐帶,一旦受損,就會導致懷孕母畜流產, 這可能是布魯氏菌能夠引起懷孕母畜流產的發病機制之一,其具體分子機制還需做進一步研究[9]。前期本課題組對2308強毒株和RB51疫苗株的差異蛋白組學進行研究,發現了多個差異表達蛋白,如DsbG等,RB51疫苗是通過2308強毒株體外傳代篩選后獲得的弱毒疫苗株,其與2308強毒株的差異表達蛋白對毒力因子的挖掘具有重要意義。
已有研究表明,氧化還原物是布魯氏菌的關鍵毒力因子之一,如cydB、narG和norE等;Dsb蛋白家族是可溶性的巰基二硫鍵氧化還原酶[10]。Dsb蛋白質不僅在調制細菌分泌中發揮作用,而且影響著膜蛋白質折疊過程中二硫鍵的形成和重排[11]。DsbG是Dsb蛋白家族的成員之一,有二硫鍵異構酶和分子伴侶活性。有研究表明DsbG在催化折疊或部分折疊蛋白質的二硫鍵重排中占優勢[11]。在細胞周質中,DsbG在新易位蛋白的氧化方面優先于DsbA或DsbC的氧化。因此對布魯氏菌dsbG基因功能的探索有助于明確其在毒力和致病性中的作用。
在本研究中,我們成功構建了2308ΔdsbG突變株,發現dsbG基因的缺失影響布魯氏菌對宿主細胞的致病性。布魯氏菌可侵入胚胎滋養層細胞,并引起其形態發生改變;對不同時間段進入胚胎滋養層細胞內的布魯氏菌數量進行分析發現,dsbG基因缺失后,進入細胞內的布魯氏菌數量顯著低于親本株,說明該基因可能是調控布魯氏菌侵入胚胎滋養層細胞的關鍵分子之一,如需確定該基因所發揮的具體功能,還需在細胞實驗(如小鼠巨噬細胞)和動物實驗進行進一步的探索和研究。本研究選用胚胎滋養層細胞對布魯氏菌dsbG基因進行研究,有利于布魯氏菌流產機制的闡述,也為進一步解析布魯氏菌的致病機制和新型疫苗的開發奠定了基礎。
利益沖突:無
引用本文格式:楊寧寧,徐明國,張歡,等.牛種布魯氏菌2308強毒株dsbG基因缺失突變株的構建與初步評價[J].中國人獸共患學報,2019,35(5):370-375.DOI:10.3969/j.issn.1002-2694.2019.00.047
