多殺性巴氏桿菌OmpH對宿主Fn和Plg粘附作用的初步研究
2019-05-20 08:42:20朱偉峰魏后軍仇汝龍范志宇陳萌萌薛家賓索朗斯珠
中國獸醫雜志 2019年10期
關鍵詞:血清
陳 露,朱偉峰,魏后軍,胡 波,仇汝龍,范志宇,陳萌萌,薛家賓,索朗斯珠,王 芳
(1. 西藏農牧學院動物科學學院,西藏林芝860000 ; 2. 江蘇省農業科學院獸醫研究所,江蘇南京210014 ; 3. 農業部獸用生物制品工程技術重點實驗室,江蘇南京210014 ; 4. 國家獸用生物制品工程技術研究中心,江蘇南京210014)
多殺性巴氏桿菌是引起多種畜禽和野生動物巴氏桿菌病的主要病原菌[1-2]。多殺性巴氏桿菌還參與其他細菌、病毒引發呼吸道病后的繼發感染,進而造成更為復雜的動物呼吸道疾病[3-4]。多殺性巴氏桿菌也是一種人獸共患病病原體。與動物密切接觸的人群的血清中巴氏桿菌抗體陽性率比未接觸者高2倍左右[5]。人類發病主要通過傷口感染。患者創口處腫脹、劇痛、發熱、形成膿腫和周圍淋巴結炎,個別病例出現敗血癥和腦膜炎[1-2]。近年來多殺性巴氏桿菌在多種動物中的臨床分離率或檢出率均位居細菌性病原的前列[6-8]。
目前已經發現了多殺性巴氏桿菌部分毒力因子及可能的致病機制,如莢膜、脂多糖、外膜蛋白等[3]。但是目前尚不能完全清楚地解釋巴氏桿菌病發病的分子過程,也無法完全確定各毒力因子在致病中的地位[9-10]。孔蛋白OmpH是多殺性巴氏桿菌的一種主要的外膜蛋白[11]。以往研究表明,OmpH是保護性抗原并具有粘附宿主細胞的能力,是一種粘附因子[12-13]。很多病原菌的粘附因子除了可以粘附宿主細胞外還可以粘附胞外基質成分(如纖維連接蛋白,Fibronectin,Fn)或者通過結合宿主血漿纖維蛋白溶解酶原(Plasminogen,Plg)促進細菌對宿主細胞和組織的粘附[14-15]。因而OmpH有可能通過結合宿主Fn和Plg促進多殺性巴氏桿菌對宿主的粘附。本文旨在通過研究OmpH對Fn和Plg的粘附作用,初步揭示多殺性巴氏桿菌OmpH分子致病機制。
1 材料與方法
1.1 質粒、菌種及培養 pET28a(+)表達載體、BL21 (DE3)宿主菌、多殺性巴氏桿菌C51-17株為本試驗室保存。多殺性巴氏桿菌使用馬丁肉湯(海博公司)添加10% 新生牛血清(Gbico公司)培養。大腸桿菌使用(LB)(海博公司)液體或固體培養基培養。
1.2 OmpH基因的克隆 以多殺性巴氏桿菌 PMTB2.1菌株全基因組序列[16]為參考序列,設計引物OmpH-F:5′-cagcaaatgggtcgcggatccGCAACAGTTT-ACAATCAAGACGGT-3′(小寫字母為與載體相同的同源臂,下同),OmpH-R:5′-gcaagcttgtcgacggagctc-GAAGTGTACGCGTAAACCAACACC-3′。按照文獻[17]提取C51-17菌株基因組并進行PCR擴增。酶切質粒并通過同源重組克隆試劑盒(諾唯贊)將OmpH的PCR擴增片段整合到pET28a(+)載體上,然后將陽性重組質粒轉入BL21。挑取陽性克隆PCR鑒定正確后送北京擎科新業有限公司南京分部測序,鑒定所克隆基因是否完整及是否正確連接到載體上。
1.3 誘導表達并鑒定 對OmpH重組大腸桿菌進行IPTG誘導表達并優化誘導條件,包括比較37 ℃、28 ℃、16 ℃的表達效果,比較0.2 mM、0.4 mM、0.6 mM、0.8 mM、1 mM等終濃度下IPTG的誘導效果。重組菌誘導表達后用超聲波破碎儀器進行破碎,并離心分別收集上清和沉淀。最后進行SDS-PAGE跑膠,并進行考馬斯亮藍染色,檢測表達效果。
1.4 OmpH與多殺性巴氏桿菌感染康復血清的反應 首先進行rOmpH的SDS-PAGE,然后將蛋白轉入NC膜上。經過5%的脫脂乳封閉以后,37 ℃與本實驗室保存的C51-17株的兔感染康復血清[18](1/500)孵育1 h;PBST漂洗5遍以后加入HRP偶聯的羊抗兔IgG作為二抗,37 ℃孵育1 h;最后,漂洗后的膜用ECL顯色試劑盒(諾唯贊公司)在化學發光成像系統(天能公司)中顯色。
1.5 Fn和Plg結合活性檢測 參照文獻[14]進行Far Western Blot試驗。首先進行重組OmpH的SDS-PAGE,同時以已知的能與Fn和Plg結合的紅斑丹毒絲菌SpaA重組蛋白[14]作為對照。然后將蛋白轉到NC膜上。經過5%脫脂乳封閉以后,加入10 μg/mL人源Fn或 Plg(Sigma公司),37 ℃孵育2 h。漂洗5遍后,分別加入1 000倍稀釋的兔抗人纖維連接蛋白抗體和兔抗人血漿纖維蛋白溶解酶原抗體,37 ℃孵育1 h。漂洗后加入HRP偶聯的羊抗兔IgG作為二抗,37 ℃孵育1 h;最后,漂洗后的膜用ECL顯色試劑盒(諾唯贊公司)在化學發光成像系統中顯色(天能公司)。
1.6 OmpH多克隆血清的制備 所有實驗動物的飼養和使用均在江蘇省實驗動物監督委員會的指導下進行。所有的操作規程已經獲得江蘇省農業科學院實驗動物監督委員會的批準。將100 μg/50 μL純化的重組蛋白用等體積弗氏完全佐劑(Sigma公司)乳化后,腹腔注射免疫5只小鼠,每只100 μL,14 d后,以100 μg/ 50 μL純化的重組蛋白用等體積弗氏不完全佐劑(Sigma公司)乳化后,第2次皮下注射免疫小鼠。第2次免疫10 d后采血制備多克隆抗血清,并用蛋白A抗體純化試劑盒(金斯瑞公司)純化出血清中的IgG。
1.7 Plg和Fn的招募抑制試驗 參照文獻[19-20]進行試驗。首先用多殺性巴氏桿菌C51-17株的菌體(108CFU/孔)或者重組OmpH(1 μg/孔)包被ELISA板,4 ℃過夜;經過PBST洗滌3次,以5%脫脂乳37 ℃ 封閉1 h。以50 μL 10倍稀釋的抗OmpH IgG 37 ℃孵育0.5 h(孵育10倍稀釋的免疫前血清IgG為對照)。PBST洗滌3遍,分別與50 μL的纖維連接蛋白(100 ng)和血漿纖維蛋白溶解酶原(100 ng),37 ℃ 孵育1 h。洗滌后,每孔加入1 000倍稀釋的兔源抗血漿纖維蛋白溶解酶原 IgG或者1 000 倍稀釋的兔源抗纖維連接蛋白 IgG,37 ℃孵育0.5 h;洗滌3次,加入5 000倍稀釋的HRP偶聯的羊抗兔二抗,37 ℃孵育0.5 h。洗滌后,每孔加入100 μL顯色液(1 mg/mL TMB和0.03%H2O2),室溫避光反應10 min后,加入終止液(2M H2SO4)。在酶標儀讀取450 nm的吸光度。以抗OmpH.IgG處理組的OD值除以免疫前血清IgG處理組的OD值計算多殺性巴氏桿菌、OmpH分別對Fn和Plg的相對結合活性。試驗重復3次,用t檢驗檢測試驗組和對照組的統計學差異。
2 結果
2.1 OmpH重組蛋白的表達和純化 將擴增的OmpH基因連接到pET28a(+)載體上,陽性菌株測序結果顯示,C51-17株的OmpH基因序列與PMTB2.1的OmpH基因序列完全相同并正確連接到載體上。將重組質粒轉入BL21工程菌后,通過優化誘導條件,選用37 ℃、0.6 mM IPTG誘導表達,獲得較多的重組蛋白rOmpH(43 kDa),主要以包涵體形式表達(圖1A)。
2.2 OmpH與多殺性巴氏桿菌康復血清的反應 Western Blot結果顯示,孵育多殺性巴氏桿菌感染OmpH能夠與多殺性巴氏桿菌康復血清發生特異性反應,OmpH泳道存在1條約43 kDa大小的條帶,與rOmpH大小一致(圖1B)。
2.3 OmpH與Fn和Plg的結合活性 針對Fn和Plg結合作用檢測的Far Western Blot結果顯示,rOmpH泳道上均存在1條約43 kDa的條帶,與rOmpH大小一致(圖2),即OmpH具有結合Fn和Plg的能力。

圖1 重組OmpH蛋白的表達及與康復血清的反應
A:重組OmpH的表達與純化 ; M:分子大小標識 ; 1:無載體的BL21菌體粗蛋白 ; 2:未誘導的OmpH重組菌粗蛋白 ; 3:37 ℃誘導過夜的OmpH重組菌粗蛋白 ; 4:重組OmpH包涵體蛋白 ; B:Western Blot檢測OmpH與康復血清的反應
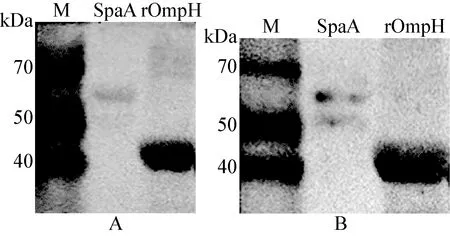
圖2 Far Western Blot檢測OmpH對宿主Fn和Plg分子的結合作用
A:OmpH對Fn的結合作用 ; B:OmpH對Plg的結合作用;M:分子大小標識;SpaA:已知的Fn和Plg結合蛋白—紅斑丹毒絲菌SpaA蛋白作為陽性對照
2.4 OmpH多克隆抗體的Fn和Plg結合抑制作用 ELISA結果如圖3所示,抗OmpH.IgG能夠顯著降低OmpH及多殺性巴氏桿菌對于Fn和Plg的結合能力。與對照組相比,經過抗OmpH.IgG孵育后OmpH對Fn和Plg的結合作用分別下降了約20%和50%,而經過抗OmpH.IgG孵育后多殺性巴氏桿菌菌體對Fn和Plg的結合作用下降了30%左右,均差異顯著(P<0.05)。
3 討論
據報道,病原菌的免疫原性較好的分子在其致病過程中發揮作用[21-22]。本研究表達了重組多殺性巴氏桿菌OmpH,Western Blot表明OmpH與多殺性巴氏桿菌高免血清存在明顯的反應。該結果提示,OmpH可能在多殺性巴氏桿菌感染與免疫反應中發揮重要作用。
Fn是細胞外基質的重要組成成分。很多病原菌通過結合該分子實現對宿主細胞的粘附進而發揮致病作用[23]。結合Fn同樣也是多殺性巴氏桿菌粘附宿主細胞的重要途徑[24]。但是Fn促進多殺性巴氏桿菌粘附的分子機制還不完全清楚。本研究發現OmpH能夠結合Fn,而且OmpH多克隆抗體能夠顯著抑制多殺性巴氏桿菌對Fn的結合。因此,多殺性巴氏桿菌可能通過OmpH結合細胞外基質進而粘附宿主細胞。
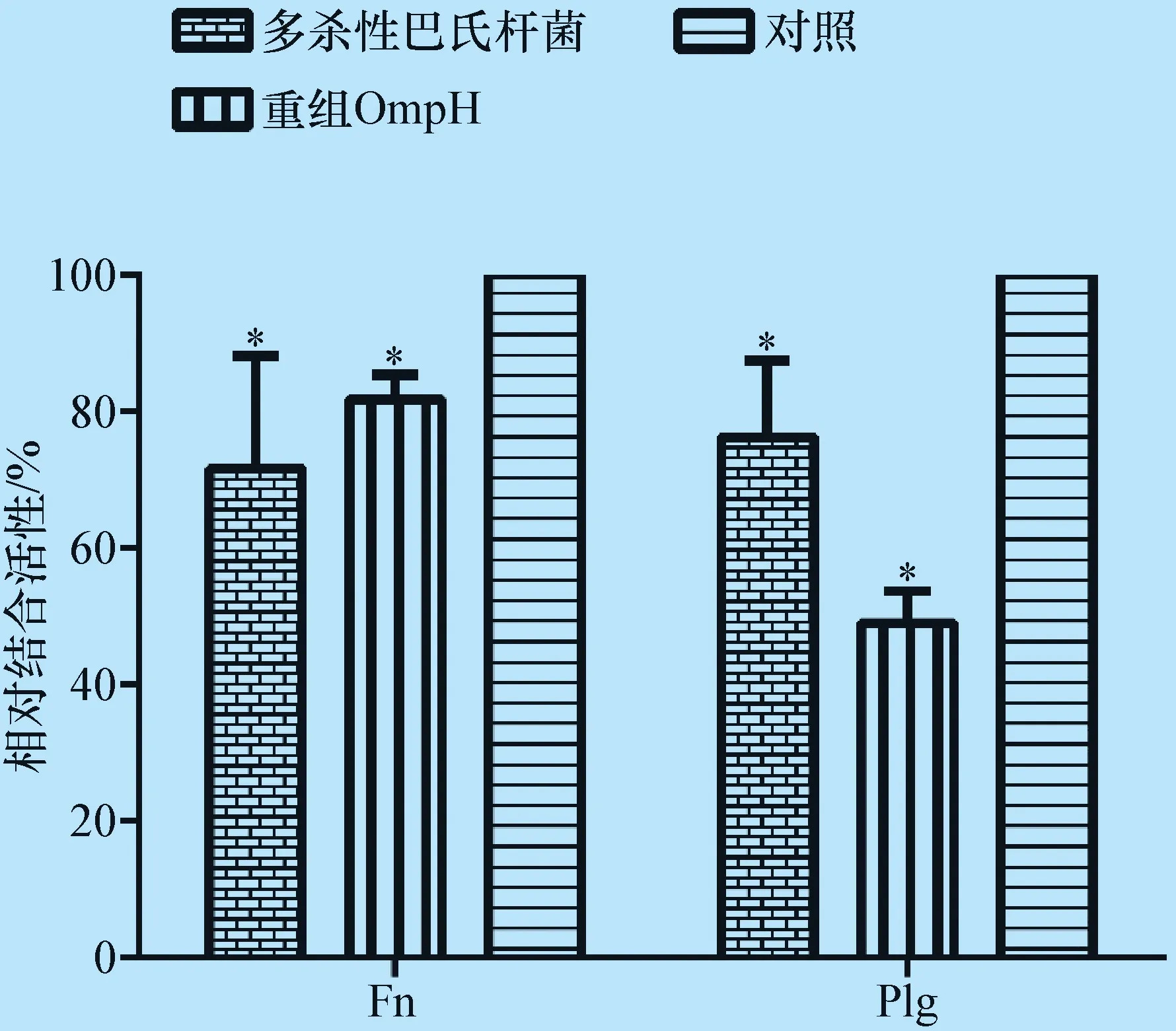
圖3 OmpH多克隆抗體對OmpH及多殺性巴氏桿菌結合宿主Fn和Plg的抑制作用
相對結合活性=抗OmpH抗體孵育組OD值/免疫前血清抗體孵育組OD值;*:P<0.05
Plg是動物體內重要的粘附分子,有多個可以與其他分子發生結合的結構域。很多病原菌通過結合Plg促進對宿主細胞的粘附作用[21, 25-26]。本實驗室已經報道多殺性巴氏桿菌能夠利用Plg[27]。本研究顯示,OmpH能夠結合Plg且OmpH多克隆抗體顯著抑制多殺性巴氏桿菌結合Plg。OmpH很可能在多殺性巴氏桿菌結合宿主Plg過程中發揮作用,最終促進多殺性巴氏桿菌的粘附作用。
綜上所述,多殺性巴氏桿菌OmpH能夠結合Fn和Plg,進而促進多殺性巴氏桿菌對宿主細胞的粘附作用。本研究增加了OmpH致病作用的認識,加深了對多殺性巴氏桿菌致病機制的理解。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
