人參內(nèi)生菌Burkholderia sp. GE 17-7制備人參皂苷Rg3的研究
2019-04-17 12:02:30李俊瑩武倫鵬康辰凱
生物學(xué)雜志 2019年2期
李俊瑩, 武倫鵬, 康辰凱, 付 玉
(1. 鞍山師范學(xué)院 化學(xué)與生命科學(xué)學(xué)院, 鞍山 114007; 2. 國(guó)家參茸產(chǎn)品質(zhì)量監(jiān)督檢驗(yàn)中心, 延吉 133002)
人參系五加科人參屬植物(PanaxginsengC.A. Mayer),被稱為“百草之王”,是中國(guó)東北三寶之一。人參與西洋參、三七等是近親,是我國(guó)傳統(tǒng)的名貴藥材,據(jù)記載已有4000多年的使用歷史。人參皂苷是人參中起主要藥理學(xué)活性的物質(zhì)。不同的人參皂苷,顯示出的性質(zhì)和功能也存在較大差異。研究表明,人參稀有皂苷能夠?qū)?nèi)分泌系統(tǒng)、中樞神經(jīng)系統(tǒng)、免疫系統(tǒng)、心腦血管系統(tǒng)等都有重要的藥理作用[1-7]。其中,人參稀有皂苷Rg3主要作用于細(xì)胞增殖周期的G2/M期,誘導(dǎo)腫瘤細(xì)胞凋亡,抑制腫瘤細(xì)胞浸潤(rùn)和轉(zhuǎn)移,對(duì)肺癌、黑色素瘤和肝癌細(xì)胞等有明顯抑制作用[8-13]。
雖然稀有人參皂苷具有很強(qiáng)的抗腫瘤活性,但這些皂苷天然含量極低,如人參皂苷Rg3在白參中的含量?jī)H為0.000 3%,在紅參中的含量約為0.03%,而Rh2在天然人參中并不存在,僅在紅參中含有約0.001%。因此,從人參中直接提取這些抗腫瘤稀有皂苷顯然不符合實(shí)際。目前,制備稀有人參皂苷的方法主要有化學(xué)法和生物轉(zhuǎn)化法,相對(duì)于污染環(huán)境、專一性差的化學(xué)法,環(huán)境友好、專一性強(qiáng)的生物轉(zhuǎn)化法表現(xiàn)出了更強(qiáng)的競(jìng)爭(zhēng)力。
內(nèi)生菌是一類存在于植物組織中而不引起侵染癥狀的重要微生物資源。在地球已知的接近30種高等植物中,每個(gè)物種都與多種內(nèi)生菌共生[14]。早在100多年前,植物內(nèi)生微生物已經(jīng)被發(fā)現(xiàn)。但長(zhǎng)期以來,內(nèi)生菌的存在一直被忽略。直到20世紀(jì)30年代,由于牲畜食用了感染內(nèi)生真菌的牧草引發(fā)中毒,帶來畜牧業(yè)嚴(yán)重?fù)p失后,植物內(nèi)生菌才作為一種新的微生物受到了國(guó)內(nèi)外學(xué)者的廣泛關(guān)注。內(nèi)生菌不僅在生物防治和促進(jìn)增長(zhǎng)寄主植物等方面顯示很強(qiáng)的生態(tài)功能[15-17],也在分解有機(jī)物方面表現(xiàn)出強(qiáng)大的功能[18]。
Park等[19-20]從1~4年生人參根部分離鑒定了200多種人參內(nèi)生真菌并對(duì)其多樣性進(jìn)行了分析;Wu等從遼東楤木(Araliaelata)根中分離到能夠產(chǎn)生三萜人參皂苷Rb2和Re的內(nèi)生真菌[21],他們還從15年生人參根部中分離得到產(chǎn)生人參主皂苷Rb2和Rc的內(nèi)生菌[22]。Cho等[23]從5年生園參根中分離出36種人參內(nèi)生細(xì)菌并經(jīng)行了鑒定,其中有3種內(nèi)生菌對(duì)立枯絲核菌有抑制作用。姜云等[24]利用研磨法和組織塊法對(duì)4年生人參根、莖、葉不同器官中共分離獲得152株內(nèi)生細(xì)菌和46株內(nèi)生真菌,發(fā)現(xiàn)有3種菌對(duì)灰霉病有明顯的抑制作用。曲紅光等[25]分離了6年生人參內(nèi)生細(xì)菌18株,其中有3株內(nèi)生細(xì)菌代謝產(chǎn)物粗提物對(duì)人宮頸癌細(xì)胞Hela具有較好的抗腫瘤活性。林星辰等[26]利用人參內(nèi)生菌B69菌株抑制人參根腐病。目前,對(duì)人參內(nèi)生菌的研究主要集中在內(nèi)生菌的分離、代謝產(chǎn)物的抗癌及拮抗作用方面,關(guān)于人參內(nèi)生菌微生物轉(zhuǎn)化制備稀有人參皂苷的研究很少。崔磊等[27]利用黨參內(nèi)生菌D19轉(zhuǎn)化人參根總皂苷為稀有人參皂苷F2和C-K;郭從亮等[28]利用一種植物內(nèi)生菌Coniochaetasp.對(duì)三七總皂苷中人參皂苷Rb1進(jìn)行了特異性轉(zhuǎn)化。本課題組前期從17年生野山參中分離、篩選獲得一株產(chǎn)β-葡萄糖苷酶的內(nèi)生真菌Burkholderiasp. GE 17-7,本研究旨在研究其轉(zhuǎn)化原人參二醇型和原三醇型人參主皂苷,探討其水解人參主皂苷制備稀有皂苷Rg3的特異性。
1材料與方法
1.1實(shí)驗(yàn)材料
菌株Burkholderiasp. GE 17-7由17年生的鮮人參根分離得到,目前由延邊大學(xué)分析測(cè)試中心尹成日課題組保藏并提供,具體分離方法參照文獻(xiàn)[29];人參標(biāo)準(zhǔn)品20(S)-Rb1、20(S)-Rb2、20(S)-Rc、20(S)-Rd、20(S)-Re、20(S)-Rg1、20(S)-Rg2、20(S)-Rg3、20(S)-Rh1、F1和C-K采購自成都曼思特生物科技有限公司;薄層層析板Silica gel60-F254購自德國(guó)Merck公司;3-(4,5-二甲基噻唑-2)-2,5-二苯基四氮唑溴鹽購自美國(guó)Sigma公司;乙腈(色譜純)、甲醇(色譜純)購自美國(guó)TEDIA公司;其他試劑及藥品均為國(guó)產(chǎn)分析純。
1.2菌株Burkholderia sp. GE 17-7的活化
將菌株Burkholderiasp. GE 17-7從凍存管中取出,加入PDB液體培養(yǎng)基中搖培,取100 μL涂布于PDA平板培養(yǎng)基上,放于30℃下生化培養(yǎng)箱中培養(yǎng)3 d,觀察菌落形態(tài)后純化,進(jìn)行下一步實(shí)驗(yàn)。
1.3微生物轉(zhuǎn)化人參主皂苷
菌株Burkholderiasp. GE 17-7放到搖瓶中液體培養(yǎng),達(dá)到生長(zhǎng)對(duì)數(shù)期后,分別與人參主皂苷Rb1、Rb2、Rc、Rd、Re和Rg1水溶液按照體積比為1∶1混合,放到30℃的空氣浴振蕩器中150 r/min下培養(yǎng)3 d。每隔5 h取出部分培養(yǎng)液,低溫濃縮后用乙醚溶液萃取,飽和正丁醇處理后旋干,得到了反應(yīng)產(chǎn)物并用薄層色譜法(TLC)、高效液相色譜法(HPLC)和核磁共振法(NMR)進(jìn)行分析。
1.4分析方法
1.4.1TLC色譜條件
材料:硅膠60板;展開劑:氯仿-甲醇-水按體積比為10∶5∶1;顯色劑:10%的硫酸乙醇溶液。通過與標(biāo)準(zhǔn)品的Rf值比對(duì),定性分析樣品中人參皂苷的種類和變化。
1.4.2HPLC色譜條件
色譜柱:BDS HYPERSIL C18柱(250 mm × 4.6 mm,5 μm);流速:1.0 mL/min;柱溫:250℃;洗脫液:水(A)和乙腈(B)進(jìn)行二元洗脫;梯度條件是:0~13 min,A∶B(77∶23,V/V);13~33 min,A∶B(46∶54,V/V);33~45 min,A∶B(32∶68,V/V);55~60 min,A∶B(0∶100,V/V);60~63 min,A∶B(77∶23,V/V)。每一個(gè)人參皂苷都通過與皂苷標(biāo)準(zhǔn)譜圖的保留時(shí)間一一對(duì)應(yīng),其含量主要通過峰面積來計(jì)算。
1.4.3NMR條件
利用瑞士Bruke Av 300核磁共振波譜儀對(duì)單體皂苷進(jìn)行核磁共振檢測(cè)。探頭:5 mm BBO;溶劑:Pyridine-d5 (氘代吡啶)。進(jìn)行包括13C NMR檢測(cè)。
2結(jié)果與分析
2.1TLC色譜分析結(jié)果
利用人參內(nèi)生菌Burkholderiasp. GE 17-7生物轉(zhuǎn)化原人參二醇型主皂苷[包括20(S)-Rb1、20(S)-Rb2、20(S)-Rc和20(S)-Rd]和原三醇型人參主皂苷[包括20(S)-Re和20(S)-Rg1]。從TLC圖中可以看出,菌株GE 17-7能有效轉(zhuǎn)化原人參二醇型皂苷Rb1和Rd,且終產(chǎn)物為Rg3。菌株GE 17-7對(duì)原人參二醇型皂苷20(S)-Rb2和20(S)-Rc、原三醇型人參皂苷20(S)-Re和20(S)-Rg1均無轉(zhuǎn)化效果(圖1)。
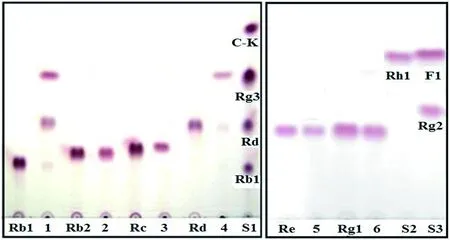
S1、S2、S3:人參皂苷標(biāo)準(zhǔn)品
圖1菌株GE 17-7水解人參皂苷Rb1(1)、Rb2(2)、Rc(3)、Rd(4)、Re(5)和Rg1(6)的TLC圖
Figure 1 TLC chromatograms of metabolites of ginsenoside Rb1(1), Rb2(2), Rc(3), Rd(4), Re(5), and Rg1(6) converted by the strain GE 17-7
2.2HPLC色譜分析結(jié)果
從HPLC圖可知,菌株GE 17-7在與人參皂苷Rb1反應(yīng)5 h后,人參皂苷Rb1的峰面積明顯減少,產(chǎn)生與標(biāo)準(zhǔn)品保留時(shí)間相同的Rd和Rg3峰(圖2-c)。反應(yīng)15 h后,人參皂苷Rb1和Rd的峰完全消失(圖2-d)。產(chǎn)生的單峰(圖2-c)和人參皂苷標(biāo)準(zhǔn)品Rg3的HPLC譜圖(圖2-a)比對(duì),出峰保留時(shí)間相同。說明終產(chǎn)物為人參稀有皂苷Rg3。
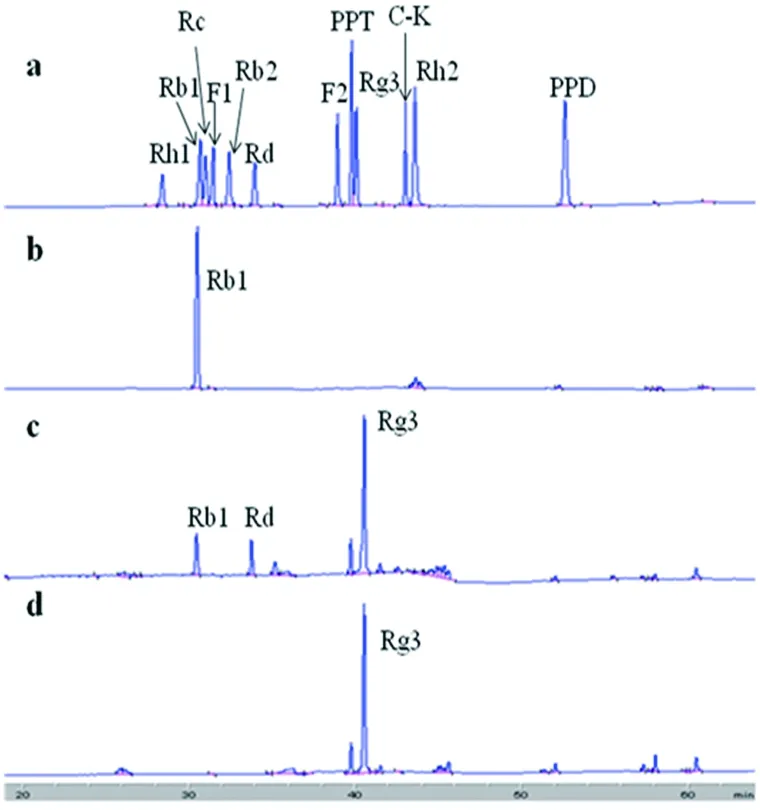
a:人參皂苷標(biāo)準(zhǔn)品;b:反應(yīng)底物;c:轉(zhuǎn)化5 h后的產(chǎn)物;d:轉(zhuǎn)化15 h后的產(chǎn)物
圖2菌株GE 17-7轉(zhuǎn)化人參皂苷Rb1的HPLC的圖
Figure 2 HPLC analysis of metabolites of ginsenoside Rb1 converted by strain GE 17-7
2.3 NMR分析結(jié)果
將HPLC分離出的最終轉(zhuǎn)化產(chǎn)物采用瑞士Bruke Av 300核磁共振波譜儀進(jìn)行檢測(cè),共出現(xiàn)42個(gè)碳信號(hào)峰,其中104.9和105.9為兩個(gè)糖端基碳信號(hào)峰,δ126.2和δ130.6分別為C-24和C-25的雙鍵碳信號(hào)峰,與手性碳原子C-20相連接的C-17,C-21和C-22分別出現(xiàn)在δ54.6,26.9和35.7,而其他13C-NMR數(shù)據(jù)與文獻(xiàn)[30]基本一致,故終產(chǎn)物為人參稀有皂苷20(S)-Rg3。
3討論
人參皂苷是人參的主要活性成分,總皂苷含量約為4%。人參皂苷屬于三萜類皂苷,按其皂苷元的結(jié)構(gòu)不同可分為3種類型:一類是齊墩果烷型,在自然界含量最少;另兩類是達(dá)瑪烷型的人參二醇型皂苷(PPD)和人參三醇型皂苷(PPT)。一般人參中人參二醇型皂苷占總皂苷的45%~60%,人參三醇型皂苷占人參皂苷總量的12%~20%,齊墩果酸型皂苷占7%~10%。由于人參二醇型和人參三醇型皂苷占人參皂苷的大多數(shù),并且人參二醇型皂苷和人參三醇型皂苷內(nèi)含有的皂苷種類比較多,因此目前關(guān)于這兩種類型皂苷的報(bào)道很多。
原人參二醇型[20(S)-protopanaxadio1,包括人參皂苷Rb1、Rb2、Rc、Rd、Rg3、Rh2等]和原三醇型人參皂苷[20(S)-protopanaxatrio1,包括人參皂苷Re、Rg1、Rg2、Rh1等]都屬于四環(huán)三萜達(dá)瑪烷型皂苷,不同之處在于二醇型皂苷母核上有3個(gè)羥基取代(C3、C12和C20),成苷的位置主要在C3和C20,糖基多為葡萄糖、阿拉伯糖和木糖;而三醇型皂苷母核上有4個(gè)羥基取代(C3、C6、C12和C20),糖苷鍵主要在C6和C20,糖基多為葡萄糖、鼠李糖和木糖。人參皂苷的結(jié)構(gòu)因糖基側(cè)鏈的不同,顯示出的性質(zhì)和活性也有很大差異[31-35]。

表1 菌株GE 17-7轉(zhuǎn)化人參皂苷Rb1的代謝產(chǎn)物人參皂苷20(S)-Rg3的13C-NMR數(shù)據(jù)(75 MHz, 溶劑: 氘代吡啶)
結(jié)合圖3原人參二醇型皂苷結(jié)構(gòu),可以很明確地看出人參內(nèi)生菌Burkholderiasp. GE 17-7未選取底物母核結(jié)構(gòu)為4個(gè)羥基取代(C3、C6、C12和C20)的原人參三醇型,而是選取了母核上有3個(gè)羥基取代(C3、C12和C20)的原人參二醇型,同時(shí)通過菌株GE 17-7對(duì)人參皂苷Rb2和Rc的水解結(jié)果,我們就可以看出其只對(duì)C-20位上O-β-D-吡喃葡萄糖基-(1→6)-β-D-吡喃葡萄糖具有水解專一性。菌株GE 17-7將人參皂苷Rb1先水解到C-20位O-β-D-吡喃葡萄糖基-(1→6)-β-D-吡喃葡萄糖苷鍵,轉(zhuǎn)化為人參皂苷Rd。再水解到C-20位O-β-D-吡喃葡萄糖基-(1→6)-β-D-吡喃葡萄糖苷鍵,形成終產(chǎn)物人參皂苷Rg3。轉(zhuǎn)化路徑:人參皂苷Rb1→人參皂苷Rd→人參稀有皂苷Rg3。
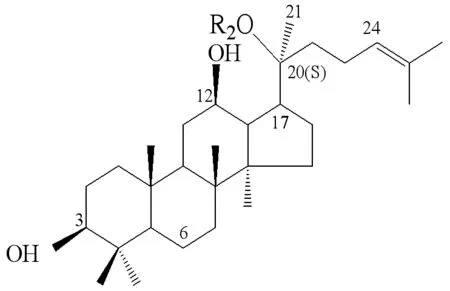

GinsenosideR1R2R3Rb1-Glc2-1Glc-Glc6-1Glc-HRb2-Glc2-1Glc-Glc6-1Arap-HRc-Glc2-1Glc-Glc6-1Araf-HRd-Glc2-1Glc-Glc-H20(S)-Rg3-Glc2-1Glc-H-HRh2-Glc-H-HCompound K-H-Glc-HCompoud Y(C-Y)-H-Glc6-1Arap-HM12(PPD)-H-H-HF2-Glc-Glc-H
圖3 20(S)-人參二醇型皂苷的結(jié)構(gòu)
Figure 3 The structure of 20 (S)-panaxadiol ginsenosides
4 結(jié)論
本研究利用從17年生野山參中分離、篩選獲得一株人參內(nèi)生真菌Burkholderiasp. GE 17-7生物轉(zhuǎn)化人參主皂苷。通過薄層色譜法、高效液相色譜法等方法對(duì)人參主皂苷(Rb1、Rb2、Rc、Rd、Re和Rg1)的轉(zhuǎn)化產(chǎn)物進(jìn)行分離純化,采用波譜解析及理化常數(shù)對(duì)其進(jìn)行結(jié)構(gòu)鑒定。結(jié)果表明人參內(nèi)生真菌Burkholderiasp. GE 17-7能夠特異性制備人參稀有皂苷20(S)-Rg3,為工業(yè)制備人參稀有皂苷提供了新的微生物資源。
