苦參雙功能核酸酶1的體外表達及結構性質分析
2019-04-17 12:02:28趙德蕊劉苗苗楊青山吳家文
生物學雜志 2019年2期
趙德蕊, 廖 怡, 劉苗苗, 楊青山, 吳家文
(1. 安徽中醫藥大學 研究生院, 合肥 230012; 2. 安徽中醫藥大學 新安醫學教育部重點實驗室科研實驗中心, 合肥 230038; 3. 安徽中醫藥大學 藥學院, 合肥230012; 4. 安徽道地中藥材品質提升協同創新中心, 合肥 230038; 5. 安徽省中醫藥科學院, 合肥 230038)
核酸酶分為核酸外切酶和核酸內切酶兩大類,限制性核酸內切酶能特異性識別切割雙鏈DNA上的特異核苷酸序列,又分為I、II和III 3種類型;I型和III型核酸內切酶不僅能特異性切割核酸,而且能對特殊堿基進行甲基化修飾,其中I型核酸內切酶切割位點距離識別位點可達數千個堿基之遠;而III型核酸內切酶切割位點距離識別位點只有25個堿基對左右。II型核酸內切酶只具有識別切割的功能,不能對堿基進行修飾,所識別的多為短的回文序列,所切割的堿基序列即為所識別的序列。本實驗克隆表達的苦參雙功能核酸酶1 (Sophoraflavescensbifunctional nuclease 1,SfBFN1) 屬于I型核酸內切酶,具有降解核糖核酸和脫氧核糖核酸的雙重功能[1],故稱為雙功能核酸酶1。
早期研究發現動物的核糖核酸酶具有抗腫瘤活性,如豹蛙的核糖核酸酶可以治療惡性胸膜間皮瘤,對該蛋白的研究已進入III期臨床試驗[2-5]; 牛精液核糖核酸酶具有抗癌細胞增殖和促進癌細胞凋亡的作用[6]。植物核酸酶1也同樣具有抗腫瘤作用,如黑松花粉細胞外核酸酶1[7]和綠豆核酸酶1[8]都能使無胸腺小鼠體內黑色素腫瘤縮小。Matousek等[9]克隆純化了番茄的雙功能核酸酶1,將這種酶及聚乙二醇修飾的化合物注射到無胸腺雌性小鼠體內,并測試其抗癌活性,均可抑制小鼠體內人黑色素瘤或前列腺癌的生長。靜脈注射蛇麻的雙功能核酸酶1到小鼠體內,發現黑色素瘤或前列腺癌的生長也明顯受到抑制[10]。
苦參(SophoraflavescensAit.)為豆科植物,其干燥根具有清熱燥濕等功效,外治滴蟲性陰道炎[11];還具有抗腫瘤、鎮痛等作用[12-14]。近年來,雙功能核酸酶1基因已經從擬南芥[1]、番茄[9]和蛇麻[10]等植物中成功被克隆,但未見苦參雙功能核酸酶1基因的研究報道。本實驗基于轉錄組測序利用苦參的功能基因資源,首次通過RT-PCR(Reverse transcription polymerase chain reaction, RT-PCR)技術擴增了SfBFN1基因的開放閱讀框(Open reading frame, ORF),構建了SfBFN1的克隆及表達載體,篩選了SfBFN1蛋白最適宜的體外表達條件,分析了SfBFN1蛋白的理化性質、結構特點,為其開發成為抗癌蛋白質藥物制劑奠定了實驗基礎。
1 材料與方法
1.1 材料與試劑
苦參(SophoraflavescensAit.)植株采自安徽中醫藥大學藥物園,由楊青山副教授鑒定。DNA凝膠提取試劑盒、瓊脂糖凝膠回收試劑盒購自天根生化科技(北京)有限公司;RevertAid First Strand cDNA Synthesis Kit和E.Z.N.A Plant RNA Kit購自德國OMEGA公司;酶制劑購自日本TaKaRa公司;引物合成及測序服務由上海生工完成。
1.2 菌株和載體質粒
大腸桿菌DH5α,pET22b(+)和pMD19T載體質粒均購自TaKaRa生物公司; 表達菌株BL21購自北京索寶生物科技有限公司。
1.3 方法
1.3.1 苦參總 RNA 的提取及 cDNA 合成
取苦參適量葉片,液氮速凍后研磨成粉,利用RNA提取試劑盒提取苦參總RNA。以Random為引物,通過逆轉錄試劑盒合成cDNA。
1.3.2SfBFN1的克隆
利用軟件Primer 5.0設計引物(SfBFN1-F:5′-ATGGGTTCTCTGAAAGGACCAATTGTTTG-3′,SfBFN1-R:5′-AAGTGTCCAGGATCTGTTCACGTTTTTTCC-3′)。25.0 μL RT-PCR反應體系中16.0 μL ddH2O,2.5 μL 10×PCR buffer(Mg2+),1.0 μL dNTP mixture(10 mmol/L),1.0 μL 5.0 μmol/μL上下游引物,2.0 μL cDNA和1.5 μL rTaq酶。PCR 擴增程序為:94℃預變性3 min;94℃變性30 s,40℃退火30 s,72℃延伸1 min,共10個循環;隨后94℃變性30 s,50℃退火30 s,72℃延伸1 min,共25個循環;最后72℃再延伸10 min。
1.3.3SfBFN1克隆重組質粒構建
將純化的SfBFN1基因片段連接到pMD19T載體,利用菌液PCR鑒定陽性克隆,NdeI /XhoI酶切鑒定質粒。
1.3.4SfBFN1表達重組質粒構建
將酶切后純化的SfBFN1基因片段連接到pET22b (+) 空載體,鑒定方法同上。
1.3.5重組質粒 pET22b-SfBFN1在大腸桿菌內誘導表達
將重組質粒pET22b-SfBFN1擴大培養,37℃培養至OD600為0.6,再分別加入不同濃度的IPTG(IPTG濃度分別為0、0.1、0.5和1.0 mmol/L),在不同溫度及時間下分別誘導蛋白表達(16℃誘導24 h、25℃誘導8 h和37℃誘導6 h)。
1.3.6 蛋白Western Blot檢測
SDS-PAGE電泳結束后,采用200 mA恒流轉膜2 h及37℃封閉(2 g脫脂奶粉和40 mL PBST)1.5 h,經過4℃一抗(1∶100)孵育過夜和37℃二抗(1∶2500)孵育1 h,最后暗室曝光。
1.3.7 蛋白生物信息學分析
利用序列處理在線工具包The Sequence Manipulation Suite(http://www.bio-soft.net/sms/index.html)將SfBFN1的堿基序列翻譯為氨基酸序列。通過ExPASy Protparam[15](http://web.expasy.org/protparam/)和Motif Scan(http://hits.isb-sib.ch/cgi-bin/motif_scan/)分析SfBFN1蛋白的理化特性及結構域。使用MEGA 5.0[16]軟件建立SfBFN1蛋白系統發育進化關系。利用PHYRE2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)軟件預測了SfBFN1蛋白的三級結構,利用PyMOL[17-18]軟件對SfBFN1蛋白的三級結構進行繪制。
2 結果
2.1 SfBFN1的克隆及重組質粒的構建
利用RNA提取試劑盒獲得苦參總RNA,結果如圖1-A;mRNA逆轉錄合成cDNA,利用SfBFN1的引物對其ORF片段進行克隆,結果如圖1-B。
將PCR產物與pMD19T載體連接獲得重組質粒,菌液PCR及酶切鑒定如圖1-C、D。酶切pMD19T-SfBFN1質粒與同樣酶切的pET22b(+) 載體連接獲得重組質粒,菌液PCR及酶切鑒定如圖1-E、F。SfBFN1基因序列及其蛋白質序列信息已投遞到GenBank數據庫,登錄號為MG891836。
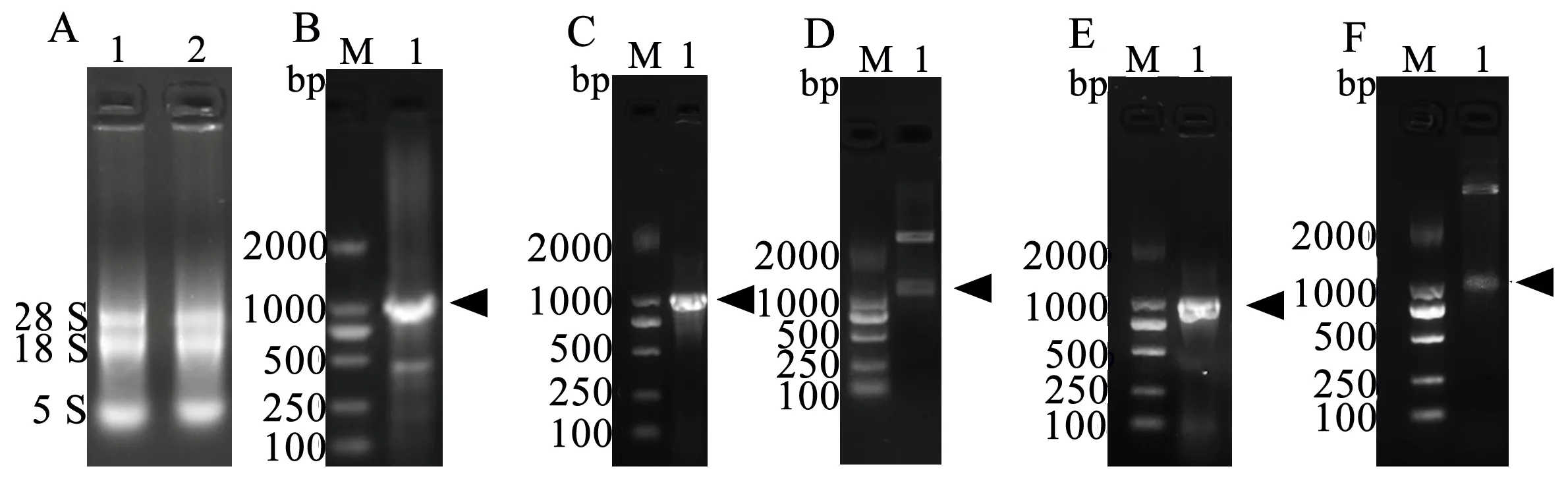
A:苦參總RNA 提取,1、2為平行樣品;B:SfBFN1基因讀碼框的克隆;C :重組質粒pMD19T-SfBFN1菌液PCR鑒定圖;D: 重組質粒pMD19T-SfBFN1酶切鑒定圖;E:重組質粒pET22b-SfBFN1菌液PCR鑒定圖;F:重組質粒pET22b-SfBFN1酶切鑒定圖。箭頭指示目的條帶
圖1SfBFN1基因讀碼框序列的克隆及重組質粒鑒定
Figure 1 The ORF cloning ofSfBFN1 and identification of recombinant plasmids
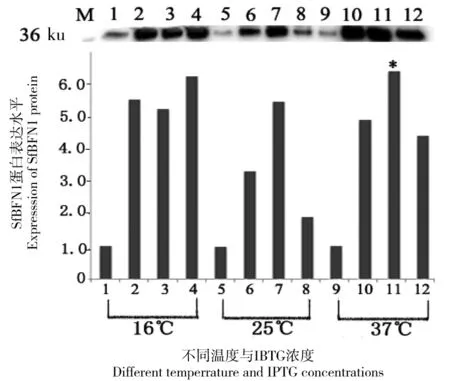
M: Marker; 1~4 分別代表在 16℃下培養24 h, IPTG 終濃度為0、0.1、0.5和1.0 mmol/L 時的蛋白表達情況;5~8 分別代表在 25℃下培養8 h, IPTG 終濃度為0、0.1、0.5及1.0 mmol/L時的蛋白表達情況;9~12 分別代表在 37℃下培養6 h, IPTG 終濃度為 0、0.1、0.5及1.0 mmol/L時的蛋白質表達情況
圖2SfBFN1的誘導表達情況
Figure 2 Expression ofSfBFN1
2.2 重組質粒 pET22b-SfBFN1的誘導表達
將pET22b-SfBFN1轉化到BL21感受態細胞中,以不同的IPTG 濃度分別在16℃、25℃和37℃誘導表達,利用Western Blot鑒定目的蛋白的表達情況,結果(如圖2)顯示:在37℃下,當IPTG的終濃度為0.5 mmol/L時目的蛋白的表達量最多(*標示)。
2.3 生物信息學分析結果
2.3.1SfBFN1序列分析
將SfBFN1的cDNA 序列翻譯成蛋白質序列,如圖 3 所示:讀碼框長度為984 bp,編碼327個氨基酸,TAA為終止密碼子。

圖3 SfBFN1的開放讀碼框序列和氨基酸序列Figure 3 ORF and amino acid sequence of SfBFN1
2.3.2 SfBFN1蛋白理化性質
利用Protparam 在線軟件分析SfBFN1蛋白,其分子式C1585H2573N447O466S26,相對分子質量約為36.2 ku,理論等電點8.9,280 nm處的摩爾消光系數為29 045 M-1·cm-1。該蛋白含有30個纈氨酸(Val),其含量為9.2%;含有41個正電荷氨基酸和33個負電荷氨基酸。
2.3.3 SfBFN1基元及結構域分析
Motif Scan分析顯示SfBFN1編碼蛋白的基元和結構域信息見表1,主要含有4個N-糖基化位點,4個酪蛋白激酶II磷酸化位點和2個酪氨酸激酶磷酸化位點等。
2.3.4 SfBFN1分子系統進化分析
利用 NCBI中SmartBlast在線軟件對SfBFN1編碼蛋白進行序列比對;通過MEGA5.0軟件對不同物種雙功能核酸酶1進行同源進化樹分析(圖4),SfBFN1的蛋白序列與羽扇豆、花生及木豆等豆科植物的雙功能核酸酶1蛋白序列的同源性約為80%,說明該酶在豆科植物之間具有保守性;其中,SfBFN1與羽扇豆雙功能核酸酶1蛋白序列的同源性高達84%,它們之間的遺傳距離最短,其親緣關系最近。
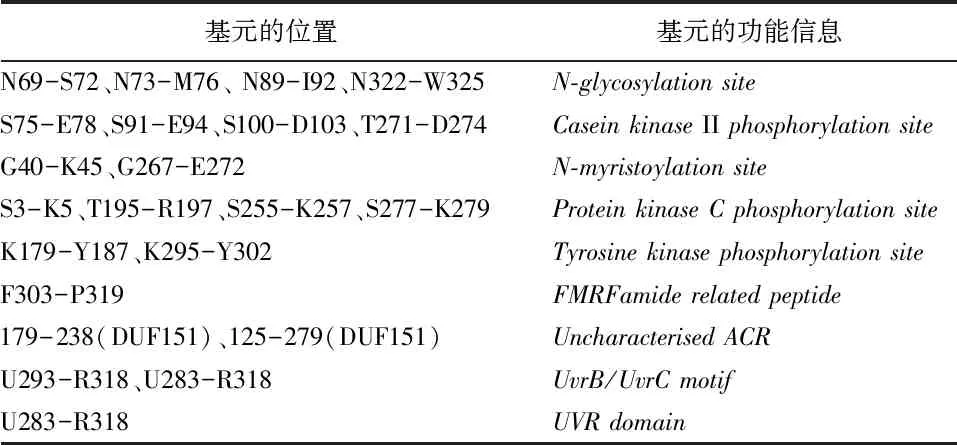
表1 SfBFN1蛋白的基元及功能信息

Lupinusangustifolius(XP_019416686.1);Arachisipaensis(XP_016167659.1);Cajanuscajan(XP_020219569.1);Vignasinensis(XP_014504471.1);Vignaangularis(XP_017428013.1);Malusdomestica(XP_008359765.1);Pyrusbretschneideri(XP_009335956.1);Arabidopsislyrata(XP_020891185.1);Brassicaoleracea(XP_013621486.1);Raphanussativus(XP_018472402.1);Brassicarapa(XP_009128096.1);Camelinasativa(XP_010416415.1)
圖4 SfBFN1蛋白的系統進化樹
Figure 4 Phylogenetic analysis of SfBFN1 protein
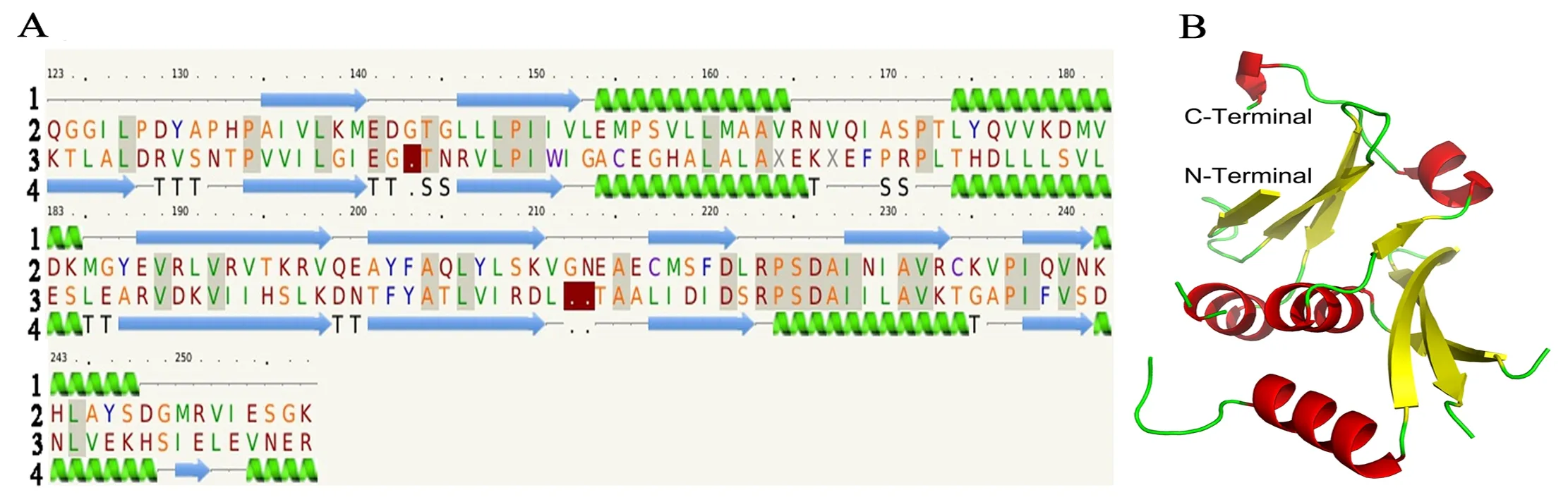
圖A 中1為二級結構示意圖(綠色示意α螺旋,藍色示意β折疊);2 為SfBFN1蛋白氨基酸序列;3 為模板TMO160 蛋白(PDB ID: 1sj5)氨基酸序列;4 為模板TMO160 蛋白的二級結構。圖B為 SfBFN1的三級結構模型(紅色示意α螺旋,黃色示意β折疊)
圖5 SfBFN1蛋白結構模型
Figure 5 The structure model of SfBFN1 protein
2.3.5 SfBFN1蛋白結構
本研究利用PHYRE2在線軟件預測了SfBFN1蛋白的部分三級結構,氨基酸序列覆蓋率為40%(覆蓋SfBFN1蛋白132個氨基酸殘基,分別為Q123-D142、T144-V211和E214-K257,序列一致性為23%,可信度為100%)。結果顯示SfBFN1酶蛋白中央保守區域 (Q123-K257) 富含α螺旋和β折疊,其中含5個α螺旋 (E154-A162、L174-M185、P223-C234、K242-D248和F254-G256) 和7個β折疊(A135-M140、C146-V152、E188-V198、A201-K210、C217-D211、I228-R233和I238-N241),如圖5-A所示。
利用PyMOL軟件對SfBFN1的三級結構進行繪制(圖5-B,結構區域為Q123-K257)。SfBFN1酶蛋白的三級結構中7個β折疊被5個α 螺旋包裹在內部,構成了蛋白質立體結構的穩定的內部框架。
3 結論與展望
本實驗成功克隆了SfBFN1基因的開放閱讀框,篩選了SfBFN1蛋白的體外表達條件,分析了其理化性質、同源關系及結構特點。研究發現SfBFN1蛋白與羽扇豆的雙功能核酸酶1蛋白之間的遺傳距離最短,同源性高達84%,這與苦參是豆科植物是相符的。
植物雙功能核酸酶1具有重要的生物學功能[19-20],如氧響應因子1通過與雙功能核酸酶1的啟動子結合從而影響擬南芥葉片的衰老[21],雙功能核酸酶1的啟動子不僅在衰老的器官中有活性,在其他器官中(如正在發育的花、種子,以及受精后的花等)也有活性,說明該酶還參與擬南芥不同的生長發育過程[22]。
動物核酸酶能抑制和殺死腫瘤細胞,如牛胰腺核糖核酸酶、牛精液核糖核酸酶和豹蛙的核糖核酸酶等,這些酶與細胞的相互作用大多是非特異性的,所以它們的應用會引起較強的毒副作用,例如代謝變化、免疫反應或干擾精子生長等[23-24]。雖然植物核酸酶的功能尚未被廣泛發掘,但已證實它具有抗腫瘤作用及較小的毒副反應,其原因可能是動植物具有不同的糖基化模式[9,25],故植物雙功能核酸酶1蛋白作為潛在的抗腫瘤抑制劑引起了越來越多的關注。
通過X射線熒光光譜法發現番茄的雙功能核酸酶1 蛋白中鋅離子有3種空間排列方式,且該蛋白通過二硫鍵保持結構穩定,這點與牛精液核糖核酸酶的結構類似[26]。研究還發現番茄、蛇麻及蕓苔的雙功能核酸酶1蛋白具有桔青霉核酸酶1相似的結構特征,且這三種雙功能核酸酶經過N-糖基化修飾后切割雙鏈DNA,該過程由Zn2+催化發生[27-30]。SfBFN1蛋白的三級結構富含α螺旋與β折疊,5個α 螺旋包裹著7個β折疊,構成了穩定的立體結構,且蛋白中含有4個N-糖基化位點(表1),蛋白質序列中含有10個半胱氨酸,推測此蛋白酶也是通過二硫鍵保持結構穩定,由Zn2+催化此酶經過N-糖基化修飾后切割雙鏈DNA。本研究為進一步純化SfBFN1蛋白,將其開發成抗癌藥物奠定了實驗基礎。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現代企業(2015年9期)2015-02-28 18:56:50
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
