鐵皮石斛SERK基因的鑒定與表達分析
2019-04-17 12:09:08李良建馬凌暉劉楚琪王萬軍
生物學雜志 2019年2期
梁 易, 李良建, 馬凌暉, 劉楚琪, 李 暢, 王萬軍
(西南交通大學 生命科學與工程學院,成都 610031)
體細胞胚發生相關類受體蛋白激酶(Somatic embryogenesis receptor-like kinase,SERK)廣泛存在于植物中。1997年Schmidt等首先從胡蘿卜(Daucuscarota)下胚軸發現第一個SERK基因,即DcSERK[1]。隨后,研究人員在擬南芥(Arabidopsisthaliana)[2]、玉米(Zeamays)[3]、水稻(Oryzasativa)[4-5]、苜蓿(Medicagotruncatula)[6]、小麥(Triticumaestivum)[7]、大麥(Hordeumvulgare)[8]、鴨茅 (Dactylisglomerata)[9]、可可(Theobromacacao)[10]、文心蘭(Cyrtochilumloxense)[11]和卡特蘭(Cattleyamaxima)[12]等植物中分別發現了SERK基因。除了少數SERK基因的表達在體細胞胚內檢測不到外,其他已經報道功能的SERK基因一般都參與了體細胞胚的發生[13]。
SERK蛋白屬于富亮氨酸重復序列類受體蛋白激酶(Leucine-rich repeat receptor-like kinases, LRR-RLKs)的第二亞類,具有典型的胞外受體結合域、跨膜結構域和胞內激酶活性結構域;SPP基序(Ser-Pro-Pro,富脯氨酸結構域)是SERKs區別于其他LRR-RLKs蛋白的特征,也是鑒定SERK蛋白的依據[14]。SPP基序位于胞外受體結合域與跨膜結構域之間,可能起鉸鏈作用使胞外信號接收更加靈活。SERK蛋白高度同源,特別是胞內激酶區;但胞外區的同源性稍低,不同的SERK蛋白可能接收不同的刺激信號,因而胞外受體結合域的序列變化更多。此外,SERK基因結構也很保守,一般含有11個外顯子與10個內含子;外顯子與編碼蛋白功能域相對應。以擬南芥AtSERK1為例,信號肽(Signal peptide, SP)由外顯子1編碼,跨膜結構域(Transmembrane region, TM)由外顯子8編碼,外顯子9~11編碼胞內激酶活性結構域[2]。
Schmidt等發現DcSERK只在胡蘿卜胚性細胞內表達且只表達到體細胞胚的球形期[1],蜜柑CiSERK也只在胚性細胞內檢測到[15],擬南芥外植體中過量表達AtSERK1時能提高成胚率3~4倍[2],水稻表達OsSERK1也會誘導體細胞胚發生能力增強[5],金花茶(Camellianitidissima)體胚發生5個不同階段都有SERK表達[16];這些事實都表明SERK基因在體細胞胚發生過程起重要作用。近年來的研究發現SERK基因的表達并不局限于體細胞胚發生過程,它在植物其它生理過程中也起作用。例如擬南芥SERKs蛋白與ERECTA、TMM組成受體復合體共同激活下游MAP激酶級聯反應調控氣孔發育[17];擬南芥serk1、bak1、bkk1三突變體只能長出非常短的根且分生組織非常少[18],促進植物生長的肽類激素Phytosulfokine (PSK)能在根中介導SERK蛋白與PSKR1的異源二聚化并啟動信號轉導[19-20]。SERKs作為共受體還參與包括油菜素內酯信號傳導、花藥絨氈層細胞分化和維管束發育及花器官脫落在內的諸多生長發育過程;還參與細胞死亡調控、植物對病原菌的先天免疫反應等過程[21]。
鐵皮石斛種子萌發需形成原球莖結構,過程可分為8個時期,分別為種胚活化期、原球莖形成期、原分生組織形成期、頂端分生組織形成期、葉維管系統形成期、根形成期、原球莖退化期、苗形成期,記為P1~P8。本研究通過對鐵皮石斛SERK基因的篩選鑒定,并利用熒光定量PCR方法研究基因在原球莖發育各時期和不同組織的表達情況,同時對基因進行生物信息學分析,為深入研究SERK基因的功能奠定了理論基礎,也為研究鐵皮石斛原球莖發育的分子機理奠定基礎。
1 材料與方法
1.1 材料
鐵皮石斛種子;RNA提取試劑盒購自成都biofite公司;PCR反應體系購自上海生工,引物由上海生工合成;熒光定量試劑盒與逆轉錄體系購自TaKaRa公司。
1.2 方法
1.2.1 原球莖發育各時期與各組織器官材料的獲取
鐵皮石斛種子接種于1/2 MS半固體培養基上,分別取種胚活化期 (P1,~5 d)、原球莖形成期(P2,~15 d)、原分生組織形成期 (P,~28 d)、頂端分生組織形成期 (P4,~37 d)、橢球形原球莖期 (P5,~45 d)、葉原基維管系統形成期 (P6,~60 d)、根端分生組織形成期 (P7,~80 d)、原球莖基部退化期 (P8,~100 d)與幼苗(P9,~120 d)材料。愈傷組織由莖段誘導,根、莖、葉、花等植物組織采集于成熟的鐵皮石斛植株。原材料樣品用液氮速凍后于-80℃保存備用。
1.2.2 鐵皮石斛總RNA提取及逆轉錄
取上述凍存原材料,用植物RNA提取試劑盒(biofite公司)提取總RNA,DNaseⅠ膜上消化去除基因組污染。電泳檢測到28 S與18 S兩條帶,測得OD260/280在1.8~2.1之間,總RNA -80℃保存備用。以上述總RNA為材料,使用逆轉錄酶Reverse Transcriptase M-MLV(RNase H-,TaKaRa)體系和引物Oligo(dT)18逆轉錄合成cDNA,-20℃保存備用。從每份cDNA取少量得混合cDNA,-20℃保存備用。
1.2.3 鐵皮石斛SERK基因的篩選與鑒定
SERK基因的篩選主要通過同源比對方法,用到的數據庫和工具包括鐵皮石斛CDS與基因組數據庫、NCBI、Blast、FGENESH+。首先從NCBI上下載其他物種已鑒定的SERK基因CDS序列,比對鐵皮石斛CDS與基因組數據庫得到目的片段;然后通過蛋白質結構、基因結構等多重特征確定目的片段是否為SERK基因。
采用3′RACE實驗方法鑒定從數據庫中篩選到的SERK基因。設計基因特異引物GSP為上游引物,下游使用通用引物3RNP。以混合cDNA為模板,PCR反應后回收目的片段,連接載體后轉化感受態大腸桿菌DH5α(菌種購自上海生工),藍白斑篩選挑取白斑單克隆,搖菌培養過夜后菌液PCR檢測合格后測序,測序結果與原序列一致,則鑒定成功。
1.2.4 qRT-PCR
根據基因的CDS序列設計qRT-PCR上游引物DoSERK2-U與下游引物DoSERK2-D,見表1。引物驗證合格后,使用SYBR Green I(購自TaKaRa)體系檢測相對表達量。
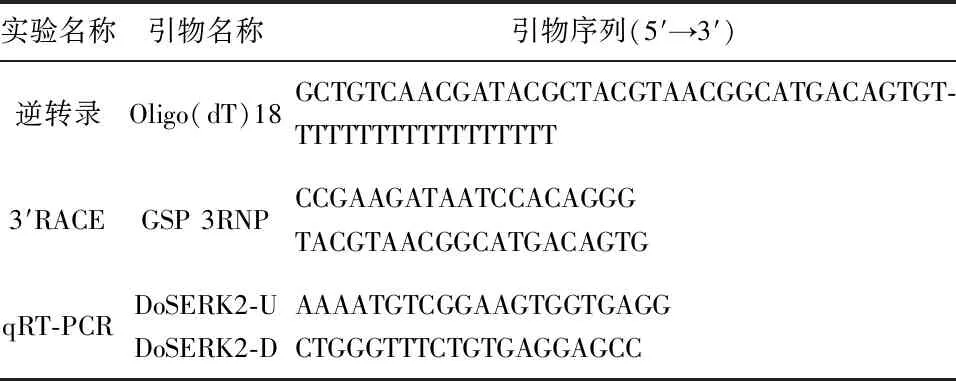
表1 研究中所用到的引物
2 結果與分析
2.1 鐵皮石斛SERK基因篩選與克隆鑒定
利用同源比對方法從鐵皮石斛CDS和基因組數據中獲得5個片段,分別位于scaffold2481、4966、1126、7408、12 804上,通過FGENESH+在線工具預測得5個基因及其編碼的蛋白質序列以及這5個基因的基因結構,將這5個基因記為gene1、gene2、gene3、gene4和gene5。基因結構圖顯示gene1、gene2、gene3和gene4含有11個外顯子,而gene5則只有8個外顯子且基因結構不全。
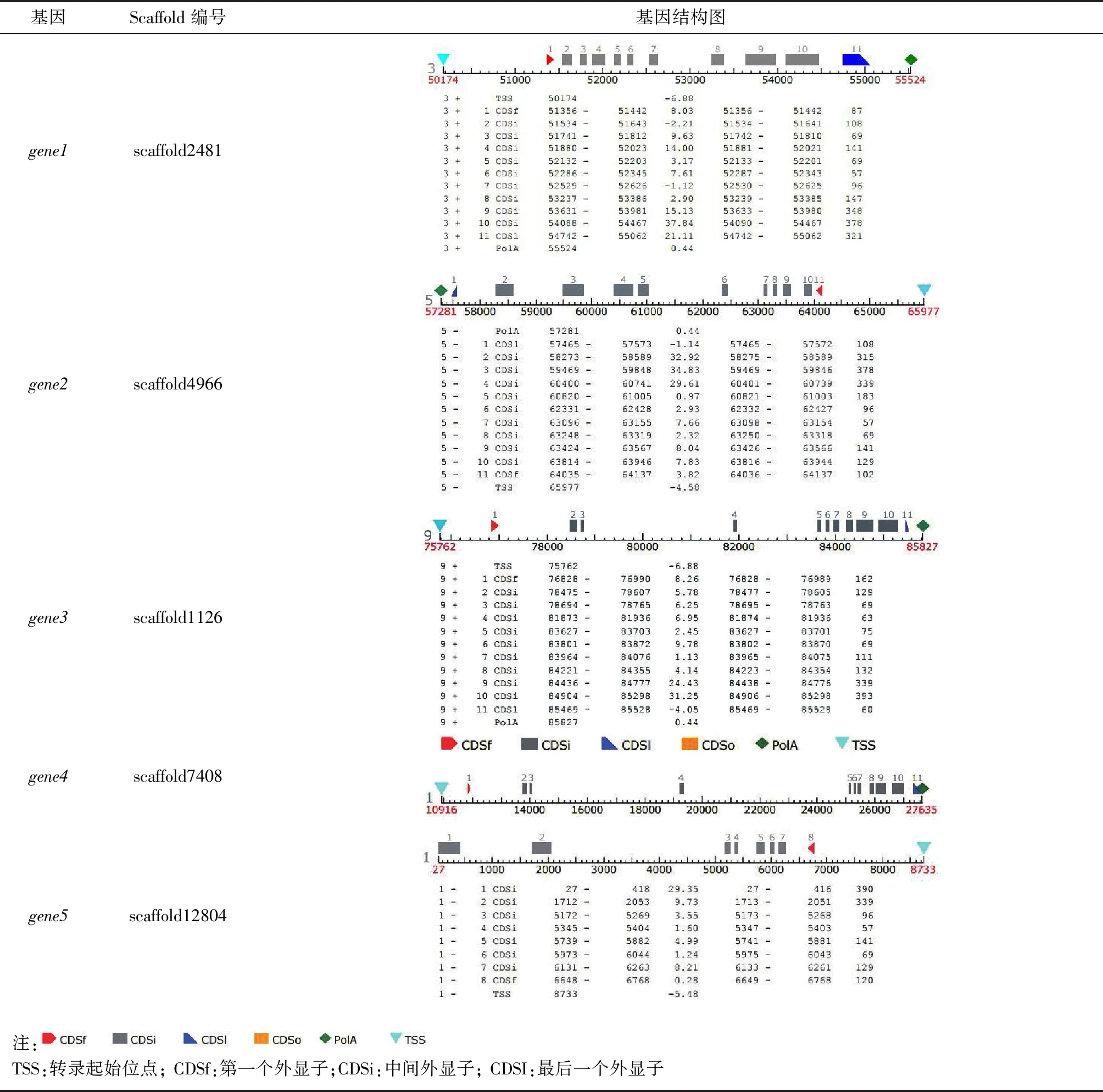
表2 基因結構
這5個基因編碼的蛋白質分別由614aa、647aa、633aa、628aa與454aa組成。序列比對結果發現這些蛋白都具有激酶結構域,其中蛋白質1、2可歸為一類,蛋白質3、4歸為一類;gene3、gene4編碼的蛋白與SERK蛋白的長度和功能域符合,且含有特征序列SPP基序(圖1,紅框)。最終,確定gene3和gene4為鐵皮石斛SERK基因,分別命名為DoSERK1和DoSERK2。查閱文獻發現DoSERK1已被王曉娟等研究報道過[22]。因此,接下來的研究以DoSERK2為對象,Blast比對發現其在NCBI上被注釋為BAK-like基因(XM_020843144)。
設計3′RACE引物GSP,PCR產物電泳檢測得到長度為750 bp左右的片段(圖2),回收、克隆與測序后得到一條長790 bp的核酸序列,比對后該片段與DoSERK2的CDS 3′端一致,含CDS區612 bp,其余178 bp為3′-UTR區。
2.2 DoSERK2的表達分析
提取鐵皮石斛種胚發育各時期和不同組織器官的總RNA,逆轉錄成cDNA;采用實時熒光定量PCR分析DoSERK2在各個時期和不同組織的相對表達量,結果顯示DoSERK2在原球莖發育各時期和不同組織器官中廣泛表達。原球莖發育過程中在P1時期(種胚活化期)表達量最高,P2~P9時期表達量低且差別不大(圖3);在花中表達量最高,根、莖、葉和愈傷組織中表達量低且差別不大。

圖1 gene1、gene2、gene3、gene4和gene5編碼蛋白與DoSERK序列比對

圖2 PCR產物凝膠電泳
2.3 DoSERK2生物信息學分析
2.3.1DoSERK2基因結構
DoSERK2基因全長15 724 bp,CDS長1877 bp,5′-UTR區長1285 bp,3′-UTR長1204 bp,其基因結構與AtSERK1基因相似,含11個外顯子和10個內含子(表2,gene4)。各外顯子與功能域基本對應(圖4),外顯子1編碼信號肽(signal peptide, SP);外顯子2編碼鋅指結構和LRR1;外顯子4編碼LRR2與LRR3;外顯子5和6分別編碼LRR4和LRR5;外顯子7編碼SPP基序,該基序是SERK蛋白的特征序列;外顯子8編碼跨膜結構域(transmembrane region, TM);外顯子9~11編碼胞內激酶活性結構域。

P1:種胚活化期;P2:原球莖形成期;P3:原分生組織形成期;P4:頂端分生組織形成期;P5:橢球形原球莖期;P6:葉原基維管系統形成期;P7:根端分生組織形成期;P8:原球莖基部退化期;P9:幼苗
圖3原球莖發育各時期和不同組織器官中DoSERK2相對表達量
Figure 3 Expression ofDoSERK2 in the development of protocorm, and in different tissues and organs

SP信號肽;ZIP鋅指結構;LRR富亮氨酸重復序列;SPP富脯氨酸基序;TM跨膜區;Kinase激酶區
圖4DoSERK2各外顯子對應的功能域
Figure 4 The functional domain corresponding to the exons ofDoSERK2
2.3.2DoSERK2調控元件分析
PlantCARE分析DoSERK2基因TSS上游1500 bp核苷酸序列,得調控元件分析表(表3)。TATA-Box位于TSS上游-35~30 bp,除含有TATA-box與CAAT-box基本元件外,還有4個元件涉及光響應,2個元件與茉莉酸甲酯(MeJA)應答有關,4個元件可能為MYB轉錄因子結合位點。
2.3.3 DoSERK2蛋白分析
DoSERK2基因CDS長1877 bp,編碼628個氨基酸。DoSERK2蛋白分子量為69.4 ku,等電點pI為5.64,是一個親水性蛋白;SignalP預測其1~28位氨基酸為信號肽,28~29位為切割位點;WoLFPSORT與TMHMM預測其為質膜蛋白,1~241位氨基酸位于細胞質膜外,242~264位氨基酸為跨膜螺旋區,265~628位氨基酸位于胞內。

表3 DoSERK2 調控元件分析
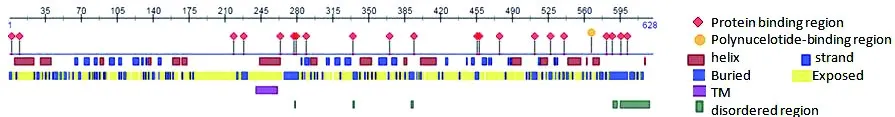
圖5 PreditProtein預測DoSERK2蛋白二級結構
Figure 5 Predict Protein predicts the secondary structure of DoSERK2
1)二級結構。利用PreditProtein與SOPMA網站預測DoSERK2的二級結構(圖5),DoSERK2蛋白具有 21個蛋白質結合位點,1個 DNA 結合位點,二級結構由40.61%α-螺旋、16.56%延伸鏈、7.48% β-轉角和35.35%無規卷曲組成,第242~263個氨基酸為跨膜區域,無序區域較短。
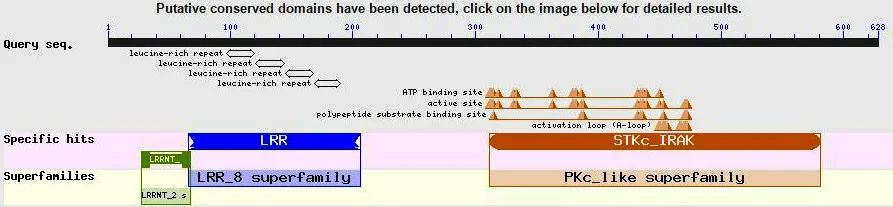
圖6 NCBI Conserved domains在線分析
2)三級結構與功能域。SWISS-MODEL預測DoSERK2胞內激酶區三級結構(圖7),有一個明顯的凹陷區(圖7,白色箭頭);NCBI Conserved domains在線分析結果(圖6)顯示了DoSERK2的功能域(圖7),ATP結合位點、活性位點、多肽底物結合位點與活性環都位于胞內激酶區,將這些氨基酸在三維結構中標示出來,發現它們都位于凹陷區附近(圖7-B,彩色)。
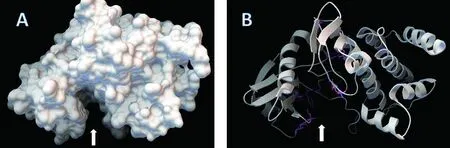
A:molecular surface; B:ribbon;sticks and balls顯示位于ATP結合位點、活性位點、多肽底物結合位點與活性環的氨基酸
圖7 SWISS-MODEL預測DoSERK2蛋白三級結構
Figure 7 SWISS-MODEL predicts the tertiary structure of DoSERK2
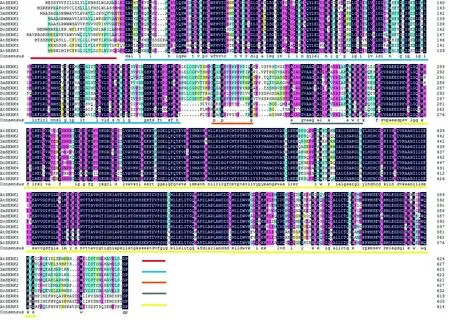
圖中功能域分別為:信號肽(紅色);LRR(亮氨酸重復序列,藍色);SPP(富脯氨酸基序,橙色);TM(跨膜區,灰色);激酶(黃色)圖8 序列比對Figure 8 Sequence alignment
2.3.4 DoSERK2蛋白序列比對與分析
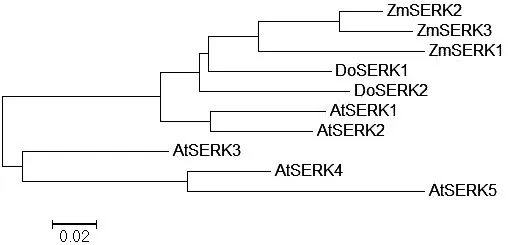
將DoSERK2與DoSERK1(AKN89445.1),玉米ZmSERK1(CAC37638.1)、ZmSERK2(CAC37639.1)、ZmSERK3(NP_001306698.1),擬南芥AtSERK1(ACN59271.1)、AtSERK2(CAF33246.1)、AtSERK3(AAK68074.1)、AtSERK4(NP_178999.2)和AtSERK5(NP_179000.3)共10條蛋白進行多序列比對,發現這些序列同源性達83.72%,都具有SERK蛋白特征功能域(圖8)。采用MEGA5.0 UPGMA法構建系統發育樹(圖9),發現DoSERK2蛋白與DoSERK1同源性最高,玉米3個SERK蛋白最接近,擬南芥AtSERK1與AtSERK2最接近;而AtSERK3、AtSERK4和AtSERK5與其他SERK蛋白同源性較低,特別是富脯氨酸區域和C-端與其他7個蛋白差別較大。這說明SERK蛋白在同一植物中既有相似性又有差異性。
The ID of corresponding mRNA in NCBI: DoSERK1 (AKN89445.1), DoSERK2 (XM_020843144), ZmSERK1(CAC37638.1), ZmSERK2 (CAC37639.1), ZmSERK3 (NP_001306698.1), AtSERK1 (ACN59271.1), AtSERK2 (CAF33246.1), AtSERK3 (AAK68074.1), AtSERK4 (NP_178999.2), AtSERK5 (NP_179000.3)
圖9系統發育樹
Figure 9 Phylogenetic tree
3 討論
SERK基因在植物中廣泛存在且在同一植物基因組中往往具有多個成員,如擬南芥有5個SERK基因,分別為AtSERK1、AtSERK2、AtSERK3、AtSERK4和AtSERK5[2];玉米有3個,分別為ZmSERK1、ZmSERK2和ZmSERK3[3];大麥有3個,為HvSERK1、HvSERK2與HvSERK3[8]。通過對鐵皮石斛基因組進行同源搜索后共發現5個目的片段,翻譯成蛋白后鑒定含SPP基序的基因有2個且最終被確定為SERK基因,分別命名為DoSERK1與DoSERK2。DoSERK1即為DoSERK,已被王曉娟等研究報道過,編碼633個氨基酸,其氨基酸序列與文心蘭(Cyrtochilumloxense)和卡特蘭(Cattleyamaxima)SERK親緣關系最近[22]。DoSERK2是篩選鑒定的另一個鐵皮石斛SERK基因,編碼628個氨基酸,序列與小蘭嶼蝴蝶蘭(Phalaenopsisequestris)BAK1-like蛋白(XP_020586612.1)、深圳擬蘭(Apostasiashenzhenica)SERK蛋白(PKA59550.1)同源性最高,分別為96%和93%。DoSERK1信號肽比DoSERK2多5個氨基酸,DoSERK2的信號肽切割位點在28~29位而DoSERK1則在33~34位[22]。DoSERK1與DoSERK2都是親水性蛋白,它們的分子量、等電點等理化特征與其他SERKs蛋白一致;功能域分析顯示DoSERK1和DoSERK2都具有SERK蛋白特征功能域。
DoSERK1在鐵皮石斛不同組織器官中均有表達,在幼嫩小苗根部表達量最高[22];說明它可能參與多種生長發育過程,但在根的發育過程中發揮重要作用。DoSERK2也在鐵皮石斛中廣泛表達,組織表達分析顯示它在花中表達量最高,而原球莖發育各時期表達分析顯示它在P1種胚活化期表達量最高。花是植物的生殖器官,其生物學功能是產生雌、雄配子體使之結合形成合子并最終發育成種子。這一進程包括雌蕊與雄蕊的發育、授粉作用、受精作用和合子的發育,可推測DoSERK2在鐵皮石斛生殖過程中發揮重要作用,但具體在哪個步驟起作用尚不清楚。鐵皮石斛胚胎發育為紫菀型,基細胞與頂細胞都參與胚的發育,種子成熟時胚胎發育停止在早期,這時種胚尚未分化,需經過原球莖這一中間結構才能形成植株。因此,原球莖發育是胚胎發育與種子萌發過程的整合,原球莖發育早期可視為胚胎發育,后期可視為種子萌發形成植株的過程。原球莖發育第一階段屬于鐵皮石斛早期胚胎發育,可推測DoSERK2在鐵皮石斛早期胚胎發育過程中起重要作用。
DoSERK1與DoSERK2作為受體跨膜激酶參與鐵皮石斛生長發育過程,其作用機制還有待研究。一般來說,SERKs與其它LRR-RLKs相互作為共受體通過磷酸化與去磷酸化接收并傳導信號,擬南芥AtSERK3與BRI1(Brassinosteroid-intensitive1)作為共受體識別蕓苔素(Brassinosteroid, BR)信號并采用可逆的磷酸化途徑引發下游基因表達,由此產生BR生理效應[23-27]。PlantCARE分析顯示DoSERK2可能參與光響應過程,茉莉酸甲酯與MYB轉錄因子可能調控DoSERK2的表達。茉莉酸甲酯是一種與損傷相關的植物激素和信號分子,能夠激發植物防御基因的表達,誘導植物的化學防御。植物MYB轉錄因子參與眾多生物學過程,主要包括初生和次生代謝反應、細胞形態與模式建成、植物生長發育、對生物和非生物脅迫的應答等。這表明DoSERK2基因可能具有多重功能,參與多種生物學過程。
