蘋(píng)果酸生物煉制研究進(jìn)展
2019-04-08 07:41:46姜紹通李興江
食品科學(xué)技術(shù)學(xué)報(bào) 2019年2期
姜紹通,李興江
(安徽省農(nóng)產(chǎn)品精深加工重點(diǎn)實(shí)驗(yàn)室/合肥工業(yè)大學(xué) 食品與生物工程學(xué)院, 安徽 合肥 230009)
蘋(píng)果酸最早是由Carl Wilhelm Scheele于1785年在蘋(píng)果汁中分離得到。蘋(píng)果酸結(jié)構(gòu)上類(lèi)似于琥珀酸及富馬酸,其內(nèi)在的四碳二元羧酸分子結(jié)構(gòu)拓展了蘋(píng)果酸的用途[1],如廣泛用于食品添加劑、醫(yī)藥化工領(lǐng)域及作為聚蘋(píng)果酸的前體物質(zhì)等[2]。
目前,大多數(shù)工業(yè)生產(chǎn)的蘋(píng)果酸通過(guò)化學(xué)合成方法制備,酶催化法合成僅占一小部分,直接發(fā)酵法合成更少。化學(xué)合成方法[3]通過(guò)加熱馬來(lái)酸得到蘋(píng)果酸;酶催化法[4]通過(guò)微生物中分離得到的富馬酸酶轉(zhuǎn)化富馬酸得到蘋(píng)果酸,其中富馬酸是由石化資源制得的馬來(lái)酸轉(zhuǎn)化而來(lái)。酶催化法[5]和化學(xué)方法的原料均來(lái)自石化資源。由可再生糖質(zhì)原料直接發(fā)酵轉(zhuǎn)化制備蘋(píng)果酸,雖然尚未在工業(yè)上廣泛應(yīng)用,但顯示出了產(chǎn)品安全特性及原料供應(yīng)豐富的巨大優(yōu)勢(shì),正在獲得越來(lái)越多的關(guān)注和研發(fā)投入。目前,蘋(píng)果酸與琥珀酸、富馬酸共同被美國(guó)能源部列為未來(lái)12種頂級(jí)生物基平臺(tái)化學(xué)品的首位[5],因此,直接發(fā)酵法代表了蘋(píng)果酸生物煉制技術(shù)的未來(lái)。
本文期望在細(xì)菌、酵母和絲狀真菌的轉(zhuǎn)化機(jī)制方面進(jìn)行分析,通過(guò)將細(xì)胞的代謝與發(fā)酵過(guò)程聯(lián)系起來(lái),討論蘋(píng)果酸生物精煉上游原料和下游發(fā)酵過(guò)程的組合。根據(jù)原料及菌種的特性考慮了水解和發(fā)酵的整合(同步糖化發(fā)酵);基于纖維素糖中共存有五碳或六碳糖,討論了五六碳共發(fā)酵;基于發(fā)酵中存在的減緩溫室效應(yīng)特點(diǎn),分析了碳的固定化;基于纖維素糖獨(dú)有的抑制特性,分析菌種對(duì)發(fā)酵抑制的適應(yīng)性;同時(shí),提出了能量調(diào)控的觀(guān)點(diǎn),以促進(jìn)發(fā)酵中的蘋(píng)果酸產(chǎn)物通量的提升。
1 蘋(píng)果酸生物煉制研究現(xiàn)狀
1.1 發(fā)酵原料的范圍研究
將可再生生物質(zhì)轉(zhuǎn)化為生物燃料、化學(xué)品及食品添加劑是當(dāng)前生物化工領(lǐng)域研究的熱點(diǎn)課題,許多不同類(lèi)型的生物質(zhì)水解產(chǎn)物均可用于制備蘋(píng)果酸。文獻(xiàn)[2,6-8]報(bào)道:用玉米芯水解產(chǎn)物可生產(chǎn)蘋(píng)果酸;Rhizopusdelemar菌株被用于轉(zhuǎn)化玉米秸稈水解物生產(chǎn)蘋(píng)果酸,也具有工業(yè)應(yīng)用意義;其他生物質(zhì)也可被用于蘋(píng)果酸發(fā)酵。這些研究表明,蘋(píng)果酸生物煉制的原料范圍很廣。
1.2 菌種的篩選研究
近年來(lái),有17種微生物被用于蘋(píng)果酸轉(zhuǎn)化研究,這些微生物可分為3種類(lèi)型:細(xì)菌、酵母和絲狀真菌。細(xì)菌具有發(fā)酵周期短、生長(zhǎng)快的優(yōu)點(diǎn),通常用于有機(jī)酸的工業(yè)發(fā)酵;作為模型微生物,酵母廣泛應(yīng)用于酒精發(fā)酵領(lǐng)域,雖然酵母不是最重要的蘋(píng)果酸生產(chǎn)菌種,但由于其代謝能力多樣,受到廣泛關(guān)注;絲狀真菌發(fā)酵蘋(píng)果酸最大的優(yōu)勢(shì)是營(yíng)養(yǎng)需求簡(jiǎn)單,利于產(chǎn)品分離。絲狀真菌是最重要的蘋(píng)果酸產(chǎn)生菌,科研人員也多圍繞該類(lèi)菌種開(kāi)展研究。產(chǎn)蘋(píng)果酸菌株情況見(jiàn)表1。Jantama等[9]發(fā)現(xiàn),大腸桿菌KJ070能夠利用10%的葡萄糖在24 h內(nèi)生產(chǎn)626 mmol的蘋(píng)果酸;Crtereal等[10]研究表明,漢遜酵母具有較好的生產(chǎn)蘋(píng)果酸及其他多種四碳二元羧酸的潛力。

表1 生產(chǎn)蘋(píng)果酸的菌株
1.3 基本代謝路徑研究
類(lèi)似丁二酸及L-乳酸,蘋(píng)果酸的合成路徑引起人們重視,尤其是其工業(yè)化生產(chǎn)調(diào)控[24]。作為四碳二羧酸最重要的生產(chǎn)菌,大腸桿菌的代謝途徑被廣泛研究[25],也可通過(guò)琥珀酸產(chǎn)生菌的代謝修飾生產(chǎn)蘋(píng)果酸;酵母由于具有很好的pH值耐受性而被人們用于蘋(píng)果酸生產(chǎn),13C同位素示蹤分析表明,Saccharomycescerevisiae[15]的蘋(píng)果酸累積通過(guò)丙酮酸的羧化、三羧酸循環(huán)及其逆循環(huán)合成而得;絲狀真菌米曲霉的13C同位素示蹤分析[26]表明,線(xiàn)粒體內(nèi)TCA循環(huán)(三羧酸循環(huán))的活性對(duì)蘋(píng)果酸的分泌貢獻(xiàn)較大。
蘋(píng)果酸生物煉制的中心代謝途徑如圖1。分析表明,細(xì)菌中蘋(píng)果酸的通量主要來(lái)源于草酰乙酸的直接代謝;酵母菌中除了草酰乙酸的代謝外,部分蘋(píng)果酸來(lái)自TCA循環(huán);絲狀真菌中除了草酰乙酸代謝和TCA循環(huán)代謝外,蘋(píng)果酸的通量也來(lái)自蘋(píng)果酸酶的直接催化以及乙醛酸循環(huán)的代謝。
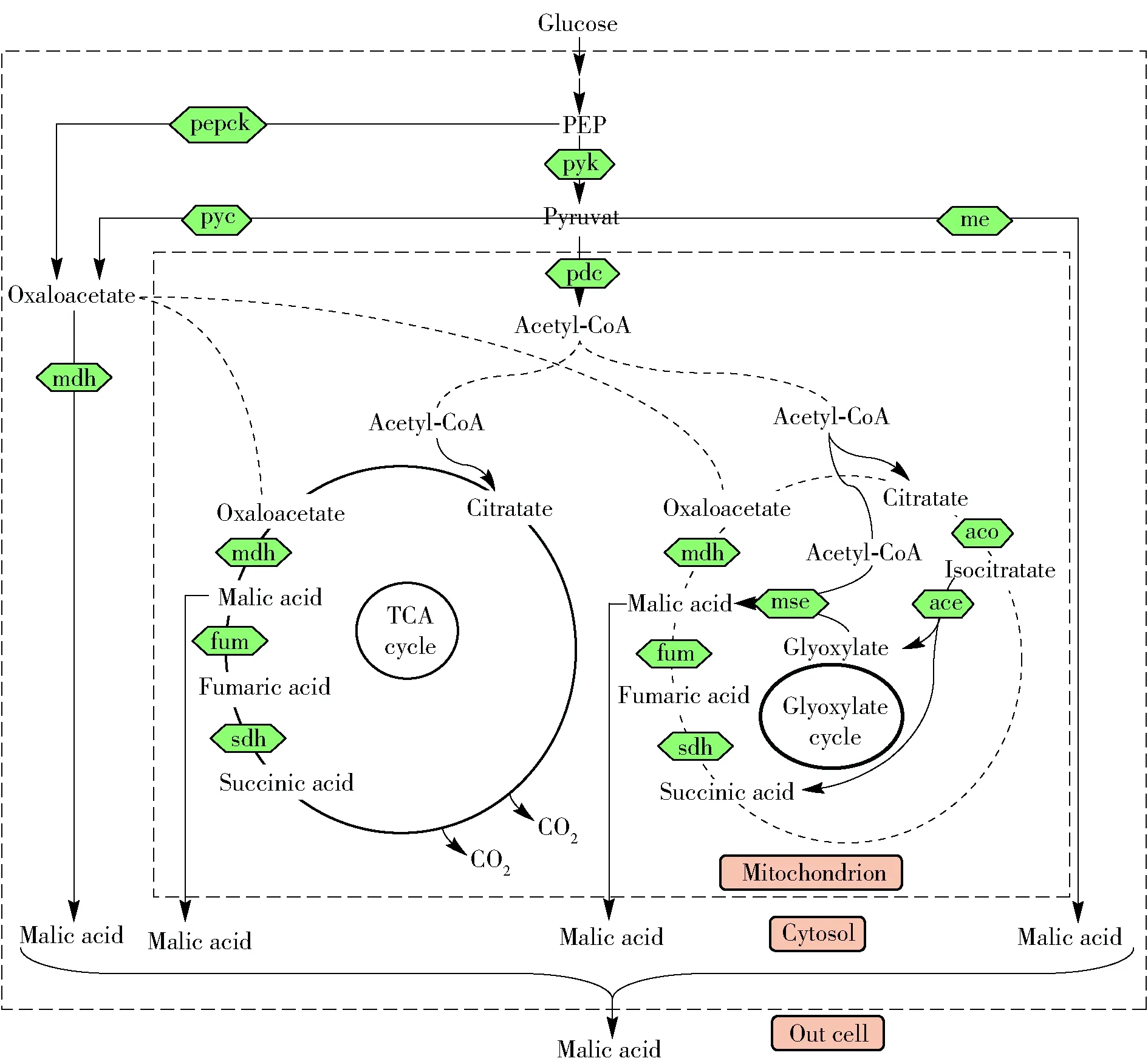
Glucose,葡萄糖;PEP, 磷酸烯醇式丙酮酸;pyk, 丙酮酸激酶;pepck,磷酸烯醇式丙酮酸羧激酶;Pyruvate, 丙酮酸;pyc, 丙酮酸羧化酶; pdc,丙酮酸脫氫酶; Oxaloacetate, 草酰乙酸;mdh, 蘋(píng)果酸脫氫酶;Acetyl-CoA, 乙酰輔酶A; Citratate, 檸檬酸;sdh, 琥珀酸脫氫酶;fum,富馬酸酶;Isocitratate, 異檸檬酸;aco, 烏頭酸酶;mse, 蘋(píng)果酸合酶;ace, 異檸檬酸裂解酶;me,蘋(píng)果酸酶;Mitochondrion, 線(xiàn)粒體;Cytosol,胞液。圖1 蘋(píng)果酸生物精煉的中心代謝途徑Fig.1 Central metabolic pathway for malic acid bio-refinery
1.4 關(guān)鍵酶研究
富馬酸酶是生產(chǎn)蘋(píng)果酸的重要酶,特別是由富馬酸為原料生產(chǎn)蘋(píng)果酸時(shí)。已在釀酒酵母中檢測(cè)到富馬酸酶,并且在面包酵母細(xì)胞中測(cè)量到其高比活性[27]。蘋(píng)果酸酶在大腸桿菌中介導(dǎo)C3代謝物羧化成C4代謝中起重要作用[28]。研究表明,也可以通過(guò)用修飾電極再生輔酶來(lái)生產(chǎn)蘋(píng)果酸[11]。此外,在米曲霉中檢測(cè)到丙酮酸羧化酶和蘋(píng)果酸脫氫酶的活性也是影響蘋(píng)果酸通量高低的主要原因[26]。
蘋(píng)果酸代謝的關(guān)鍵酶見(jiàn)表2。對(duì)這些關(guān)鍵酶的分析表明,產(chǎn)生蘋(píng)果酸的菌株具有固定CO2的能力,并且參與TCA和乙醛酸鹽循環(huán)的合成。
1.5 代謝改造策略研究
通過(guò)對(duì)蘋(píng)果酸代謝的全面研究,尤其是全細(xì)胞優(yōu)化的研究[31],人們借助代謝途徑改造生產(chǎn)多種有機(jī)酸,包括C4蘋(píng)果酸[32]。Ye等[13]通過(guò)從Thermococcuskodakarensis中提取蘋(píng)果酸酶來(lái)構(gòu)建體外人工蘋(píng)果酸合成途徑,發(fā)現(xiàn)其對(duì)葡萄糖的得率可以達(dá)到60%。
為了提高蘋(píng)果酸的產(chǎn)量,在大腸桿菌中實(shí)施了富馬酸還原酶的敲除以提升產(chǎn)物[12],根據(jù)蘋(píng)果酸的通量來(lái)源,削弱富馬酸酶可以增加蘋(píng)果酸的產(chǎn)量;同時(shí)富馬酸酶的過(guò)度表達(dá)也可以反過(guò)來(lái)從下游改善其代謝[33]。對(duì)于釀酒酵母K-901H[34],Vid24p基因功能喪失導(dǎo)致了蘋(píng)果酸脫氫酶累積,進(jìn)而實(shí)現(xiàn)蘋(píng)果酸產(chǎn)率的提升。所有這些改造策略顯著促進(jìn)產(chǎn)物代謝。蘋(píng)果酸代謝改造策略及效果如表3。
1.6 發(fā)酵工藝研究
對(duì)于發(fā)酵過(guò)程,通常采用3種類(lèi)型的發(fā)酵模型,包括深層發(fā)酵、固體發(fā)酵和酶催化。Zou等[2]探討了深層發(fā)酵過(guò)程并進(jìn)行了總結(jié)。PenicilliumsclerotiorumK302[20]可以在有氧條件下高產(chǎn)蘋(píng)果酸鈣(92.0 g/L),該深層培養(yǎng)工藝具有較大潛力。寄生曲霉CICC40365深層培養(yǎng)8 d可得到55.47 g/L的蘋(píng)果酸,黃曲霉ATCC13697和米曲霉QM No.821[37]則可通過(guò)固體發(fā)酵方法生產(chǎn)蘋(píng)果酸。酶生物催化和膜分離[38]過(guò)程被應(yīng)用于富馬酸和蘋(píng)果酸生物轉(zhuǎn)化。微生物電滲析與細(xì)胞催化相結(jié)合的方法也是一個(gè)亮點(diǎn),該方法具有電耗低和外源電子利用的多種底物的優(yōu)點(diǎn)[39]。
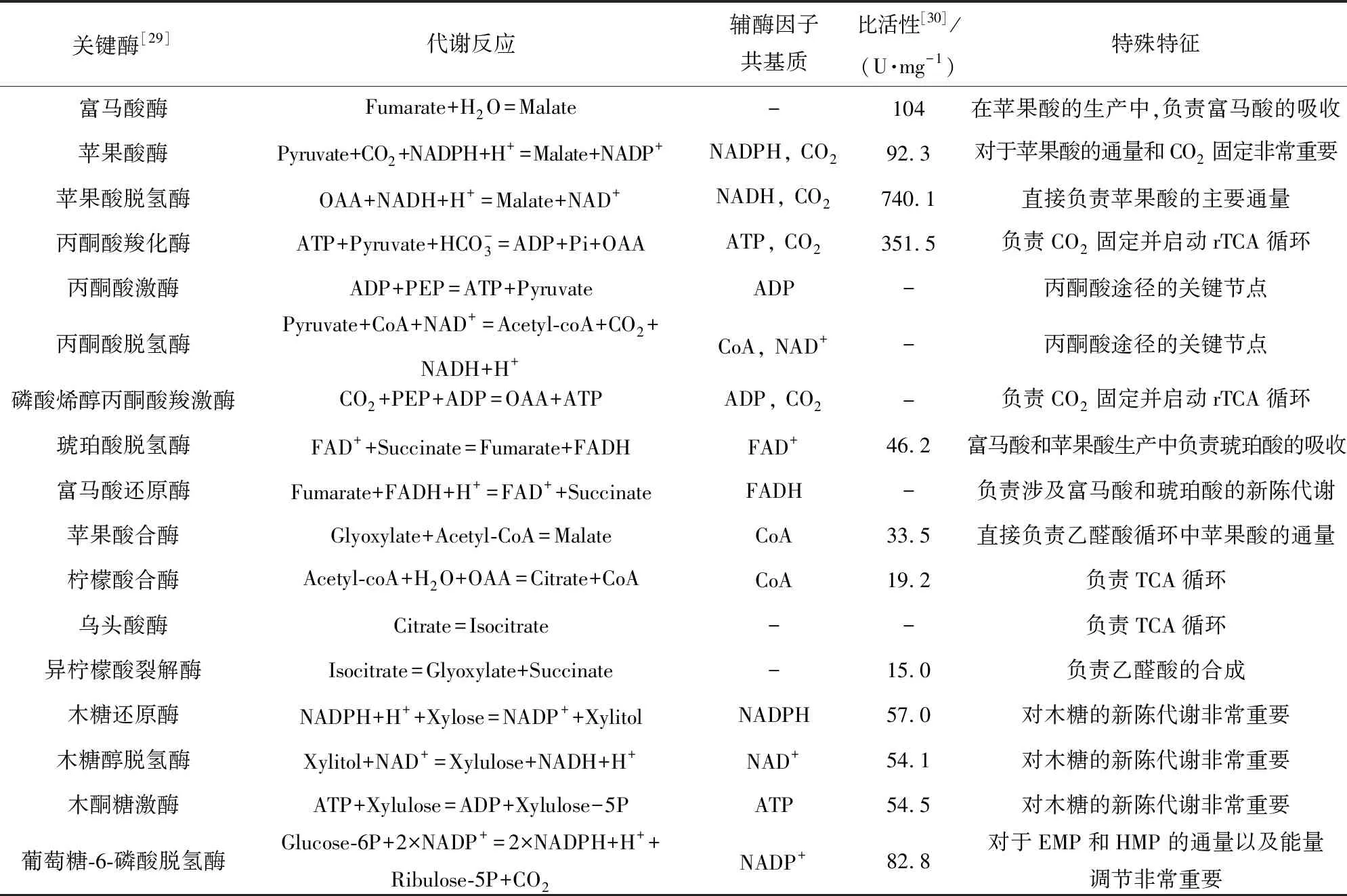
表2 蘋(píng)果酸代謝中的關(guān)鍵酶
NADPH,煙酰胺腺嘌呤二核苷酸磷酸還原態(tài);NADP+,煙酰胺腺嘌呤二核苷酸磷酸氧化態(tài);OAA,草酰乙酸;NADH,煙酰胺腺嘌呤二核苷酸還原態(tài);NAD+,煙酰胺腺嘌呤二核苷酸氧化態(tài);ATP,三磷酸腺苷;ADP,二磷酸腺苷;PEP,磷酸烯醇式丙酮酸;Pi,磷酸鹽;FADH,還原型黃素腺嘌呤二核苷酸;FAD+,黃素腺嘌呤二核苷酸;Acetyl-CoA,乙酰輔酶A;CoA,輔酶A;Xylulose-5P,5-磷酸木酮糖;Glucose-6P,6-磷酸葡萄糖;Ribulose-5P,5-磷酸核酮糖。
1.7 影響發(fā)酵的關(guān)鍵因子研究
影響發(fā)酵的主要因素包括溶解氧、中和劑、微生物形態(tài)、二氧化碳及細(xì)胞狀態(tài)等。
微生物在有氧(偶爾厭氧)條件下產(chǎn)生蘋(píng)果酸,其中溶解氧在細(xì)胞生長(zhǎng)中起關(guān)鍵作用。中和劑如Ca(OH)2、Na2CO3和CaCO3通常用于控制有機(jī)酸發(fā)酵過(guò)程中的pH值。添加50 g/L的CaCO3確保了釀酒酵母在整個(gè)蘋(píng)果酸發(fā)酵過(guò)程中保持很好的發(fā)酵指數(shù)[15],其中CaCO3通過(guò)保持恒定的pH值在蘋(píng)果酸生物合成中起重要作用。顯著影響氧轉(zhuǎn)移速率的另一種情況是真菌的形態(tài),當(dāng)絲狀真菌在深層培養(yǎng)基中生長(zhǎng)時(shí),在不同的發(fā)酵過(guò)程中形成不同類(lèi)型的形態(tài)。代謝網(wǎng)絡(luò)顯示蘋(píng)果酸積累的幾種途徑與CO2固定密切相關(guān),因此CO2影響著蘋(píng)果酸的發(fā)酵。對(duì)于蘋(píng)果酸生產(chǎn)菌大腸桿菌[40]而言,CO2參與磷酸烯醇式丙酮酸及丙酮酸的羧化。在氮饑餓條件下對(duì)米曲霉中蘋(píng)果酸生產(chǎn)狀況研究表明[21],氮源亞適量反而會(huì)增加蘋(píng)果酸的代謝,這可能與TCA循環(huán)的代謝、細(xì)胞呼吸和溶解氧吸收間接相關(guān)。
2 蘋(píng)果酸生物煉制研究發(fā)展趨勢(shì)
2.1 蘋(píng)果酸煉制各階段研究發(fā)展趨勢(shì)
2.1.1同步糖化發(fā)酵研究
纖維素原料轉(zhuǎn)化為高附加值化學(xué)品(如蘋(píng)果酸)主要包括4個(gè)加工步驟:原料預(yù)處理、水解、發(fā)酵和產(chǎn)品回收[41]。由于同步糖化發(fā)酵提高了產(chǎn)品的整體轉(zhuǎn)化效率,是未來(lái)蘋(píng)果酸生物煉制研究的重要發(fā)展趨勢(shì)。

表3 蘋(píng)果酸代謝重建策略及效果
ldhA,乳酸脫氫酶基因;adhE,乙醇脫氫酶基因;ackA,乙酸激酶基因;focA,甲酸轉(zhuǎn)運(yùn)蛋白基因;pflB,丙酮酸甲酸裂解酶基因;mgsA,甲基乙二醛合酶;pc,丙酮酸羧化酶基因;mdh,蘋(píng)果酸脫氫酶基因;fum,富馬酸酶基因;mae,蘋(píng)果酸通透酶;frdBC,富馬酸還原酶;sfcA,NAD+依賴(lài)的蘋(píng)果酸酶基因;maeB,NADPH連接的蘋(píng)果酸酶基因;fumB和fumAC,富馬酸酶的幾個(gè)基因;pck,磷酸烯醇丙酮酸羧激酶;vid24,別名gid4,葡萄糖誘導(dǎo)的降解缺陷復(fù)合物;HPr,含組氨酸蛋白。
2.1.2提高細(xì)胞耐受性的改善策略研究
當(dāng)碳源從葡萄糖擴(kuò)展到生物質(zhì)糖時(shí),必須考慮一系列抑制劑和所產(chǎn)生的細(xì)胞耐受性或適應(yīng)性。纖維素糖降解過(guò)程中形成或釋放多種細(xì)胞抑制劑,特別是在預(yù)處理過(guò)程中[42]。這些抑制劑對(duì)微生物有一定的毒性和負(fù)面影響,例如形成的糠醛會(huì)導(dǎo)致細(xì)胞損傷[43]。研究發(fā)現(xiàn),參與硫同化途徑的4種基因轉(zhuǎn)錄水平能顯著保護(hù)細(xì)胞免受糠醛氧化應(yīng)激的影響,也可通過(guò)與質(zhì)膜密切相關(guān)的化學(xué)和遺傳擾動(dòng)來(lái)改善細(xì)胞對(duì)乙酸抑制劑的耐受性[44]。如果高濃度抑制劑在發(fā)酵系統(tǒng)中積累,則需要脫毒[42]。
2.1.3五六碳糖共發(fā)酵研究
拓寬生物質(zhì)糖生物轉(zhuǎn)化蘋(píng)果酸的基質(zhì)范圍對(duì)其未來(lái)成功應(yīng)用至關(guān)重要。EMP途徑的葡萄糖代謝已經(jīng)在大多數(shù)生物體中得到很好的應(yīng)用,六碳糖(低聚糖、纖維二糖、淀粉、蔗糖和麥芽糖)都通過(guò)EMP途徑引入或參與葡萄糖代謝。五碳糖(木糖和阿拉伯糖)是來(lái)自生物質(zhì)的水解糖中的重要糖成分,含量?jī)H次于葡萄糖。葡萄糖和木糖共同發(fā)酵蘋(píng)果酸[7]對(duì)工業(yè)制備蘋(píng)果酸具有重要意義。六碳糖和五碳糖的共代謝途徑見(jiàn)圖2。
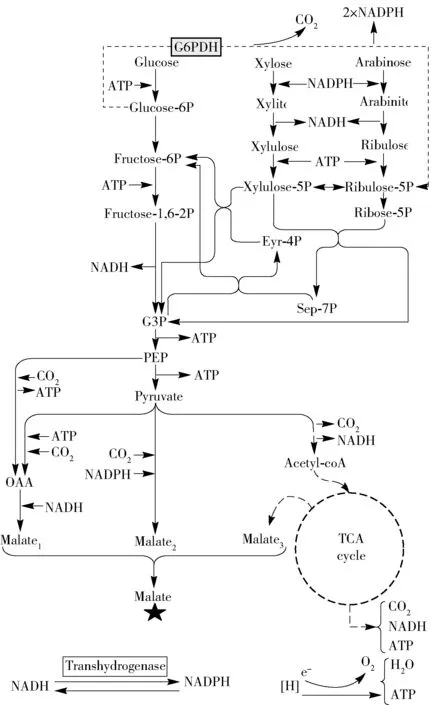
Glucose-6P,6-磷酸葡萄糖;Fructose-6P,6-磷酸果糖;Fructose-1,6-2P,1,6-二磷酸果糖;G3P,3-磷酸甘油醛;PEP,磷酸烯醇式丙酮酸;Pyruvate,丙酮酸;OAA,草酰乙酸;Xylose,木糖;Xylitol,木糖醇;Xylulose,木酮糖;Xylulose-5P,5-磷酸木酮糖;Eyr-4P,4-磷酸赤蘚糖;Sep-7P,7-景天磷酸庚酮糖;Arabinose,阿拉伯糖;Arabinitol,阿拉伯糖醇;Ribulose,核酮糖;Ribulose-5P,5-磷酸核酮糖;Ribose-5P,5-磷酸核糖;Acetyl-coA,乙酰輔酶A。圖2 六碳糖和五碳糖的共代謝途徑Fig.2 Co-metabolism pathway of hexose and pentose
2.1.4碳固定化的強(qiáng)化研究
CO2是溫室氣體的主要成分,在蘋(píng)果酸[45]發(fā)酵過(guò)程中將CO2固定在磷酸烯醇丙酮酸或丙酮酸中會(huì)產(chǎn)生較大的益處。CO2的固定效率對(duì)蘋(píng)果酸的生產(chǎn)有積極作用[7],碳固定的關(guān)鍵酶主要涉及磷酸烯醇式丙酮酸羧激酶、丙酮酸羧化酶和蘋(píng)果酸酶[15]。Rhizopusdelemar[7]胞內(nèi)的蘋(píng)果酸酶活性非常高,在蘋(píng)果酸的總產(chǎn)量中,突變菌株中檢測(cè)到20%的通量來(lái)自蘋(píng)果酸酶對(duì)CO2的直接固定化。
2.1.5能量平衡改善代謝研究
對(duì)于蘋(píng)果酸合成的基本代謝,磷酸烯醇丙酮酸羧激酶[15]和丙酮酸羧化酶合成途徑差異顯著,丙酮酸被PC(丙酮酸羧化酶)催化為OAA(草酰乙酸)的過(guò)程凈消耗ATP。當(dāng)在產(chǎn)生蘋(píng)果酸的菌株代謝中檢測(cè)到PEPCK和PC時(shí),細(xì)胞的ATP能量平衡將起決定性作用;因此,在了解細(xì)胞能量供應(yīng)時(shí),分析能量平衡很關(guān)鍵。類(lèi)似琥珀酸等四碳二羧酸的發(fā)酵[46],能量不平衡也是菌種高產(chǎn)的瓶頸。當(dāng)NADH(煙酰胺腺嘌呤二核苷酸-還原態(tài),還原型輔酶Ⅰ)細(xì)胞供應(yīng)不足時(shí),可以通過(guò)氧化還原平衡和調(diào)節(jié)NADH/NAD+(煙酰胺腺嘌呤二核苷酸-氧化態(tài))比例[47]提升系統(tǒng)能量的穩(wěn)定;同時(shí)通過(guò)乙酰輔酶A代謝,可以另外獲得NADH,因此可以通過(guò)控制電子傳遞鏈來(lái)產(chǎn)生ATP。NADPH(還原型煙酰胺腺嘌呤二核苷酸磷酸,還原型輔酶Ⅱ)則在水解糖(包括六碳糖和五碳糖)的蘋(píng)果酸生物轉(zhuǎn)化中起重要作用。如果一個(gè)菌株中葡萄糖-6-磷酸脫氫酶[7]的代謝成功啟動(dòng),這個(gè)關(guān)鍵途徑將產(chǎn)生雙倍還原力(NADPH),以補(bǔ)償能量供應(yīng),并進(jìn)而緩解整體代謝還原力[H]的短缺(NADH和NADPH),這是一種有效改善NADHP平衡的方法。
2.2 蘋(píng)果酸生物精煉研究的總體趨勢(shì)
2004年,美國(guó)能源部將蘋(píng)果酸列為未來(lái)高附加值生物基化學(xué)品的首位,蘋(píng)果酸將廣泛應(yīng)用于化工、食品和制藥等各領(lǐng)域。由于人體只能代謝L-型蘋(píng)果酸,研究人員已經(jīng)提出逐漸使用L-蘋(píng)果酸代替混合型DL-蘋(píng)果酸,特別是在食品和醫(yī)藥應(yīng)用中,歐美已經(jīng)禁止混合型產(chǎn)品的亂用。由于大多數(shù)蘋(píng)果酸產(chǎn)品是通過(guò)化學(xué)方法合成的,而富馬酸酶的催化不是真正的一步發(fā)酵,并且反應(yīng)中存在其他化學(xué)殘留的可能,因此,蘋(píng)果酸的一步法生物精煉技術(shù)具有重要發(fā)展前景。
未來(lái)蘋(píng)果酸生物煉制研究總體趨勢(shì)為,在原料利用上實(shí)現(xiàn)同步糖化發(fā)酵,利用細(xì)胞耐受性的改善提高其發(fā)酵適應(yīng)性,并進(jìn)而提升碳糖的共發(fā)酵、碳固定化及能量平衡能力,以提升蘋(píng)果酸產(chǎn)量。
3 結(jié)束語(yǔ)
利用生物煉制手段轉(zhuǎn)化可再生糖制備蘋(píng)果酸具有重要意義,其未來(lái)的發(fā)展定位是將原來(lái)的淀粉質(zhì)糖水解系統(tǒng)逐漸過(guò)渡到纖維質(zhì)水解糖系統(tǒng),而如何克服其生物轉(zhuǎn)化過(guò)程中的諸多限制因素至關(guān)重要。本文介紹和總結(jié)了蘋(píng)果酸生物煉制流程,包括原料選擇、菌種篩選、基本代謝路徑研究、關(guān)鍵酶研究、發(fā)酵工藝研究以及關(guān)鍵影響因子研究;同時(shí),統(tǒng)籌分析了轉(zhuǎn)化生物體的內(nèi)部代謝與外在條件之間的關(guān)聯(lián),進(jìn)而提出蘋(píng)果酸生物轉(zhuǎn)化研究的發(fā)展趨勢(shì),期望為蘋(píng)果酸生物精煉提供一定參考。
猜你喜歡
天天愛(ài)科學(xué)(2022年9期)2022-09-15 01:12:54
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天天愛(ài)科學(xué)(2022年4期)2022-05-23 12:41:48
當(dāng)代水產(chǎn)(2022年3期)2022-04-26 14:26:56
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科學(xué)大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
