NLRC5對HepG2.2.15細胞中MHC-Ⅰ類分子的調節作用
2019-03-27 08:46:50王文龍盛云建
重慶醫學 2019年6期
關鍵詞:水平
秦 嬌,王文龍,盛云建,強 麗,吳 剛
(西南醫科大學附屬醫院,四川瀘州 646000)
乙型肝炎病毒(HBV)感染后引起的免疫應答是肝細胞損傷及炎癥發生的主要機制,而炎癥反復存在是慢性乙型肝炎進展為肝硬化甚至肝癌的重要因素。在HBV感染期間,HBV特異性T細胞的數量和功能與HBV控制情況有關[1]。因此,HBV特異性細胞毒性T淋巴細胞(CTL)不能有效清除感染HBV的細胞是造成乙型肝炎慢性化甚至肝癌的重要因素。經典的主要組織相容性復合體Ⅰ類分子(MHC-Ⅰ類分子)通過提呈抗原肽而激活T淋巴細胞,CD8+CTL識別MHC-Ⅰ類分子提呈的內源性抗原肽,參與適應性免疫應答。在肝母細胞癌實驗中發現HBV感染可下調MHC-Ⅰ類分子HLA-ABC、HLA-E和MICA的表達[2]。
肽聚糖的核苷酸結合寡聚化結構域樣受體家族(NLRs家族)是細胞內模式識別受體(PRR)中一個很大的蛋白家族。NLRC5蛋白作為NLRs家族成員之一,近年多項研究表明,在天然免疫及適應性免疫起重要作用。NLRC5蛋白可參與MHC-Ⅰ基因及其相關基因的調節,如β2-微球蛋白、Tap1和Lmp2基因的表達[3-4]。而NLRC5對HBV感染后所致的MHC-Ⅰ下調是否起作用,目前尚未見相關報道。Hep G2.2.15細胞是Hep G2細胞的衍生系,可持續、穩定地釋放HBV。筆者將NLRC5能否誘導MHC-Ⅰ類分子表達增加的調控機制應用于Hep G2.2.15細胞,觀察NLRC5在Hep G2.2.15細胞中對MHC-Ⅰ類分子表達的調控作用。
1 材料與方法
1.1細胞及材料 Hep G2細胞株由本實驗室保存,Hep G2.2.15細胞由重慶醫科大學肝研所贈予,復蘇、培養至細胞呈對數期生長用于實驗。 DMEM培養基及胎牛血清(美國Hyclone公司);liprofectamineTM2000(美國Introvigen公司)、Opti-MEM (美國Gibco公司)。pc DNA3.1質粒購自中國質粒菌株基因庫;總RNA提取試劑盒、逆轉錄試劑盒(天根公司);PCR引物由上海Invitrogrn生物公司合成。 Anti-NLRC5(一抗)、山羊抗兔二抗(HRP,英國Abcam公司)。FITC標記鼠抗人HLA-ABC抗體(美國eBioscience公司)。
1.2方法
1.2.1實驗分組 A組:Hep G2組,B組:Hep G2+IFN-γ(5 ng/mL)組,C組:Hep G2.2.15組,D組:Hep G2.2.15+IFN-γ(5 ng/mL)組,E組:Hep G2.2.15+pcDNA3.1-NLRC5-GFP;F組:Hep G2.2.15 + pcDNA3.1組。
1.2.2質粒轉染 待細胞數量達80%時準備轉染,將250 μL Opti-MEM與30 μL的 liprofectamineTM2000混合后靜置20 min,分別將pcDNA3.1-NLRC5-GFP、pcDNA3.1加入混有轉染劑的培養基中靜置10 min。加入3每毫升×106細胞懸液,混勻后置于培養箱中,6 h后換液。
1.2.3RT-PCR 從細胞中提取細胞總RNA。取 0.5 μg RNA按逆轉錄試劑盒制備cDNA,逆轉錄原液稀釋10倍,將試劑及樣本cDNA混勻,放入定量PCR儀,并按下列條件進行cDNA擴增反應:94 ℃預變性10 min,94 ℃變性30 s,58 ℃退火30 s,68 ℃延伸25 s,共40個循環。所有樣本均設置3個平行管。
1.2.4Western blot 收集細胞進行蛋白提取,配置濃度為0.5 mg/mL標準液,根據BCA法測定各樣品的蛋白濃度,然后進行SDS-PAGE電泳,分別以60 V電泳濃縮膠、120 V電泳分離膠。選用PVDF膜進行轉膜,室溫封閉2 h,一抗4 ℃孵育過夜,TBST洗膜后加入(1∶3 500),室溫下孵育2 h;用ECL化學發光法顯影。
1.2.5流式細胞學檢測 取對數生長的細胞,經胰蛋白酶消化后制成細胞懸液,調定濃度為每毫升1×106個,將細胞用2 μL/mL的CFSE在37 ℃染色30 min,洗滌2次,以 FITC標記的鼠抗人HLA-ABC抗體室溫孵育10 min,使用流式細胞儀進行檢測,每個抗體重復3次,記錄激發波長488 nm處綠色熒光。
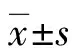
2 結 果
2.1HBV對NLRC5轉錄水平及蛋白的表達影響 與Hep G2組相比,Hep G2.2.15組細胞中的NLRC5轉錄水平及蛋白表達水平均下降(P<0.05),見圖1。

圖1 HBV對NLRC5轉錄水平及蛋白的表達影響
2.2IFN-γ對NLRC5轉錄水平及蛋白的表達影響 Hep G2細胞加入IFN-γ后,NLRC5的轉錄水平及蛋白表達水平均增加(P<0.05),Hep G2.2.15細胞在IFN-γ刺激后不能提高NLRC5的蛋白表達水平,見圖2。

圖2 IFN-γ對NLRC5轉錄水平及蛋白的表達影響
2.3HBV對MHC-Ⅰ類分子表達的影響 與Hep G2細胞相比,Hep G2.2.15細胞中的MHC-Ⅰ類分子的表達水平降低(P<0.05),見圖3。

圖3 HBV對MHC-Ⅰ類分子表達的影響
2.4轉染NLRC5蛋白后Hep G2.2.15細胞的表達情況 轉染NLRC5后的轉錄及蛋白表達水平均增加(P<0.05),見圖4。
2.5MHC-Ⅰ類分子在穩定表達NLCR5的Hep G2.2.15細胞表面的表達情況 C組、E組、F組的MHC-Ⅰ類分子MFI分別為16.17、17.86、16.59。與未轉染組相比,過表達NLRC5蛋白后的Hep G2.2.15細胞表面的MHC-Ⅰ類分子表達上調(P<0.05),見圖5。
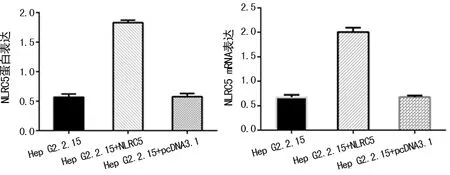
圖4 轉染NLRC5蛋白后Hep G2.2.15細胞的表達情況
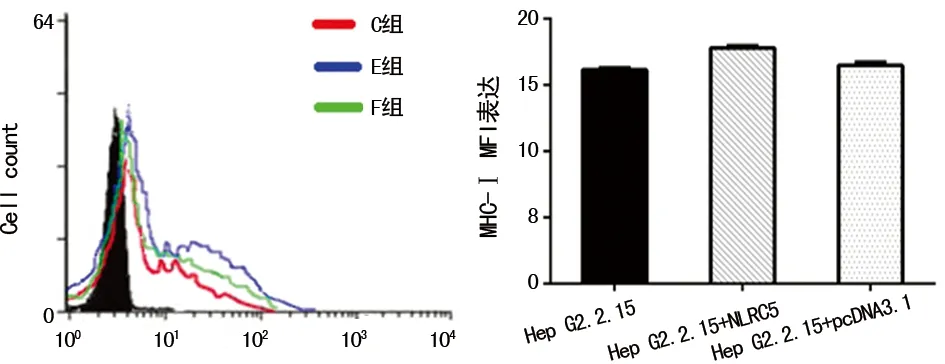
圖5 MHC-Ⅰ類分子在穩定表達NLCR5的Hep G2.2.15細胞表面的表達情況
3 討 論
病原相關分子模式(PAMP)作為先天免疫反應的第一道防御入侵微生物的防線,其識別依賴于幾類PRR。NLRC5蛋白作為NLRs家族成員之一,近年來廣泛在天然免疫及適應性免疫應答、抗感染、腫瘤等領域被研究。NLRC5具有非典型NLRs結構,即位于中心的腺苷酸三磷酸酶結構域、位于N端的效應器控制域(CARD 屬于死亡折疊域),與NLRs其他家族成員的CARD或PYD沒有同源性,為非典型的CARD結構域及位于C端最長的亮氨酸重復序列區域(LRR結構)[5-8]。研究發現,NLRC5廣泛表達于人體多個組織,而免疫相關組織,如骨髓造血組織、淋巴結、脾和外周血淋巴細胞中NLRC5的表達最高[9],表明NLRC5參與免疫應答過程。NLRC5表達被TLR3配體[poly(I∶C)]、病毒感染或IFN-γ觸發[7,10-11]。HBV是一種雙鏈DNA病毒,在本實驗中發現,在HBV感染后的肝細胞中可發現NLRC5表達,表明NLRC5在HBV感染機體后引起的免疫應答過程中起著一定的作用,可能參與了HBV的清除作用。IFN-γ是一種主要由Th1細胞、CD8+T淋巴細胞、NK細胞、巨噬細胞等免疫細胞分泌的細胞因子,IFN-γ可通過刺激JAK-STAT信號、途徑激活NLRC5基因表達[12]。研究發現,免疫系統的IFN-γ-NLRC5-MHC-Ⅰ軸對于CD8+T細胞應答和細胞內病原體的有效殺傷至關重要[13]。本實驗用等量的IFN-γ分別刺激Hep G2細胞和Hep G2.2.15細胞后得到NLRC5在轉錄水平及蛋白水平均呈上升趨勢。
筆者發現經典MHC-Ⅰ類分子表達在Hep G2.2.15細胞中低于Hep G2細胞,Hep G2.2.15細胞不僅具有Hep G2細胞生物學活性,同時又能夠持續、穩定地分泌HBV,表明HBV感染機體情況下可下調MHC-Ⅰ類分子表達,這與文獻一致[2]。MHC-Ⅰ類分子分布于有核細胞表面,通過提呈抗原肽而激活CD8+T淋巴細胞,參與適應性免疫應答。HBV感染宿主細胞后,其表面MHC-Ⅰ類分子表達下調或丟失,因此在宿主細胞表面不易形成足夠的抗原肽-MHC分子復合物,導致機體出現免疫耐受。目前認為,HBV感染機體后誘導機體出現免疫耐受是導致乙型肝炎慢性化的重要因素[14]。HBV特異性免疫應答在HBV清除過程中起重要作用,MHC-Ⅰ類分子限制性的CD8+CTL可誘導肝細胞凋亡,也可分泌IFN-γ,以非細胞裂解機制抑制其他肝細胞內HBV基因復制和表達。慢性HBV感染患者的CTL免疫應答能力減弱是HBV感染趨于慢性化的主要原因[15]。
盡管NLRC5在天然免疫應答中的作用仍有爭議,但最近發現的MHC-Ⅰ類反式激活因子(CITA)也屬于NLR蛋白家族,并構成MHC-Ⅰ類基因轉錄激活的關鍵調節因子[16]。NLRC5作為MHC-Ⅰ類分子的反式激活因子,誘導經典MHC-Ⅰ的表達(即HLA-A、HLA-B、HLA-C),非經典的Ⅰ類(即HLA-E、HLA-F,HLA-G),β亞基(B2M),免疫蛋白酶體組分(PSMB9,即LMP2)和肽轉運蛋白(TAP1)[17]。有研究團隊發現,流感病毒感染NLRC5缺陷型小鼠,在體內降低了MHC-Ⅰ類分子表達,CD8+T細胞功能受損及病毒滴度增加[18]。NLR家族成員NLRC5在各種類型的感染過程中調節MHC-Ⅰ的表達,但其在對HBV免疫中的作用尚未得到充分研究。為探求HBV感染機體時NLRC5能否刺激MHC-Ⅰ類分子表達,筆者將外源性NLRC5轉染到Hep G2.2.15細胞,經RT-PCR及流式細胞術分別得到上調的MHC-Ⅰ類分子表達。表明在Hep G2.2.15細胞中,NLRC5可上調MHC-Ⅰ類分子表達,提出NLRC5對HBV感染的免疫應答中起重要作用。
上述研究發現,HBV感染后NLRC5蛋白和MHC-Ⅰ類分子表達均下調。HBV感染人體后可降低其抗原呈遞能力,從而逃逸免疫監督,導致特異性CTL不能有效清除HBV,機體呈持續HBsAg、HBeAg陽性表現。HBV特異性CTL免疫殺傷效應的降低可能與HBV感染后導致的NLRC5表達下調有關。在HepG2.2.15細胞中過表達NLRC5,證實了NLRC5可以上調HepG2.2.15細胞MHC-Ⅰ類分子表達,而MHC-Ⅰ類分子表達增加促進HBV特異性CTL更好地發揮免疫殺傷效應,這對揭示乙型肝炎免疫耐受機制具有重要意義。因此,NLRC5是否通過MHC-Ⅰ類分子增強HBV特異性CD8+T淋巴細胞的免疫殺傷作用可進一步證實,從而為打破乙型肝炎免疫耐受提供理論依據。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
