家蠶超長鏈脂肪酸延長酶基因Bmelo424的克隆及功能分析
2019-03-27 11:49:42左偉東康寧李春林欒悅童曉玲代方銀魯成
生物工程學報 2019年3期
左偉東,康寧,李春林,欒悅,童曉玲,代方銀,魯成
1 西南大學 家蠶基因組生物學國家重點實驗室 農業(yè)農村部蠶桑生物學與遺傳育種重點實驗室,重慶 400715
2 西南大學 生物技術學院,重慶 400715
超長鏈脂肪酸延長酶 (Elongases of very long chain fatty acid,ELOVL) 是定位于內質網中的微粒體酶,在催化合成長鏈脂肪酸 (C16,C18) 和超長鏈脂肪酸 (≥C20) 的過程中起到限速作用[1]。酵母中首次鑒定出的該酶的編碼基因ELO1參與延伸C14脂肪酸到C16,而ELO2和ELO3則分別對延伸C24和C26脂肪酸必不可少[2-3]。目前,在哺乳動物中已經有 7個家族成員被鑒定(ELOVL1–ELOVL7)。其中,ELOVL1負責延伸C20到C28的脂肪酸,并與C24鞘脂類的合成相關[4-5]。ELOVL2控制精巢中C28:5n-6和C30:5n-6的脂肪酸合成,進而影響小鼠的精子成熟和雄性生殖力[6]。ELOVL3參與C20–C24脂肪酸的合成,并與毛囊皮脂腺的發(fā)育以及飲食性肥胖抵抗(Diet induced obesity resistant,DIO-R) 相關[7-8]。ELOVL4參與合成C28和C30飽和脂肪酸以及C28–C38多不飽和脂肪酸,且與黃斑營養(yǎng)不良、脊髓小腦性共濟失調和可變性紅斑角皮癥相關[9-10]。ELOVL5能夠延伸C18和C20的多不飽和脂肪酸底物,而且和脊髓小腦性共濟失調以及開角型青光眼相關[11-13]。ELOVL6延伸稍短的C12–C16脂肪酸,其基因多態(tài)性和Ⅱ型糖尿病相關聯[14-15]。ELOVL7延伸C16–C22脂肪酸,并參與前列腺癌細胞增殖和巨細胞病毒復制[16-18]。
與哺乳動物不同,昆蟲ELOVL基因數量較多但功能驗證的結果較少。果蠅中第一個被鑒定的ELOVL基因elo68α在雄性生殖系統中特異性表達,其產物能夠延伸C14和C16單不飽和脂肪酸,參與雄性信息素的合成[19]。在雌性果蠅中專一表達的eloF基因,其產物能夠延伸C19-C30脂肪酸,與雌性果蠅信息素合成和求偶行為相關[20]。另外,與ELOVL6基因相似度較高的noa基因,其RNA水平的減少導致果蠅運動障礙和生存能力顯著下降,而針對精巢包囊細胞的RNAi會引起雄性不育[21]。過表達果蠅脂肪酸延長酶基因Bond能使釀酒酵母elo2/elo3雙缺失致死突變菌株恢復活力,突變Bond基因影響精母細胞胞質分裂;而且Bond基因沉默能顯著抑制雄性生育力[22-23]。此外,白紋伊蚊中脂酰輔酶A延長酶 (ELO) 的表達水平是其滯育卵抗干燥能力的重要影響因素[24]。與表皮碳氫化合物生物合成相關的ELOVL基因,在歐洲蜜蜂中也采用RT-qPCR進行了表達譜分析[25]。桔小實蠅被單增李斯特菌、金黃色葡萄球菌和大腸桿菌侵染后,noa基因的表達上調,參與調節(jié)Toll/Imd免疫信號傳導途徑[26]。在黃粉蟲中,TmELO1主要參與C16和C20脂肪酸的合成,且能形成痕量的C22和C24脂肪酸,而TmELO2僅合成C16脂肪酸,這兩個基因的RNAi都沒有改變黃粉蟲的脂肪酸組分,但TmELO1基因的RNAi會導致黃粉蟲死亡率上升[27]。
ELOVL基因在昆蟲中已有影響育性、抗干燥性、免疫信號傳導和致死等方面的功能研究,而家蠶作為重要的經濟昆蟲和鱗翅目昆蟲的典型代表,其ELOVL基因僅有信息分析方面的報道[28]。為了進一步探索ELOVL基因在家蠶中的具體功能,本研究從家蠶幼蟲中克隆獲得Bmelo424的基因序列,分析其組織表達特征及蛋白結構特點,并借助釀酒酵母真核表達系統和GC-MS探究其基因功能,同時對重組酵母細胞的溫度脅迫響應也作了分析。
1 材料與方法
1.1 材料
1.1.1 實驗昆蟲
家蠶品種Dazao取自西南大學家蠶基因資源庫,幼蟲在25 ℃室溫的自然光照條件下以新鮮桑葉飼育。在五齡三天幼蟲期取整蠶和各組織凍存于-80 ℃冰箱備用。
1.1.2 試劑耗材
DNA膠回收試劑盒和小量質粒提取試劑盒購自Omega公司;感受態(tài)細胞Trans1-T1購自北京全式金公司;釀酒酵母INVSc1和表達載體pYES2購自Invitrogen公司;TaqDNA 聚合酶、M-MLV反轉錄試劑盒、pMD19-T載體、DNA Marker、限制性內切酶NotⅠ、BamHⅠ和T4 DNA連接酶等購自TaKaRa公司;酵母抽提物、蛋白胨和葡萄糖等購自生工生物工程 (上海) 股份有限公司;酵母尿嘧啶缺陷型合成培養(yǎng)基 (SC-Ura)購自上海哈靈公司,酵母化學感受態(tài)細胞制備試劑盒購自北京天恩澤公司;RNA提取試劑盒購自BioTeke公司;SYBR Green熒光定量PCR試劑盒購自Bio-Rad公司。
1.2 方法
1.2.1 RNA提取及cDNA合成
將五齡三天家蠶各組織 (頭、體壁、中腸、脂肪體、前中絲、后絲、馬氏管、精巢、卵巢和血液) 及整蠶的凍存樣品取出,分別放入液氮預冷后的研缽中,快速研磨至細滑粉末狀,用無RNase槍頭取適量粉末于Trizol中裂解。通過RNA提取試劑盒獲得家蠶各組織及整蠶的總RNA,并使用分光光度計測定每個樣品的RNA濃度,之后利用反轉錄試劑盒制備cDNA模板。
1.2.2 Bmelo424基因的克隆
依據家蠶基因組數據庫SilkDB中的超長鏈脂肪酸延長酶信息[28],利用Primer Premier 5.0設計Bmelo424基因 (BGIBMGA000424) 的克隆引物 (表1)。以五齡三天Dazao整蠶cDNA為模板進行PCR擴增,然后通過1%瓊脂糖凝膠電泳檢測產物大小。目的DNA片段經切膠回收后,與pMD19-T載體連接,將連接產物轉化到大腸桿菌感受態(tài)細胞中,通過菌液PCR篩選陽性克隆并進行測序驗證。
1.2.3 Bmelo424基因的組織表達特征
基于文獻報道合成Bmelo424基因的熒光定量PCR引物Q-Bmelo424和內參引物sw22934[28],以家蠶五齡三天不同組織的cDNA為模板,檢測其組織表達特征。反應總體系為10 μL (SYBR Green Ⅱ 4.2 μL ,上下游引物各0.3 μL,去離子水4.2 μL,cDNA模板1 μL)。反應程序:95 ℃,30 s;95 ℃,5 s,60 ℃,30 s,共40個循環(huán)。每個樣品3個生物學重復,收集目的基因和內參基因的Cq值計算其相對表達量。
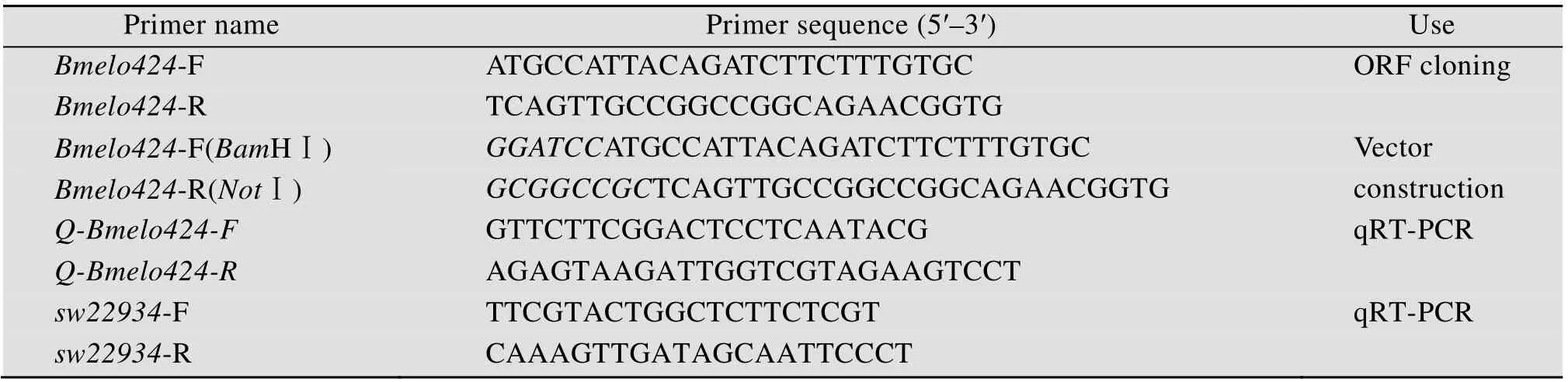
表1 文中所用引物序列Table 1 Primers used in this paper
1.2.4 Bmelo424蛋白序列和結構分析
利用在線工具對Bmelo424的跨膜結構域(http://www.cbs.dtu.dk/services/TMHMM/) 和磷酸化位點 (http://www.cbs.dtu.dk/services/NetPhos/)進行預測,采用Cell-PLoc 2.0[29](http://www.csbio.sjtu.edu.cn/bioinf/euk-multi-2/) 進行其亞細胞定位,通過PSIPRED v3.3 (http://bioinf.cs.ucl.ac.uk/psipred/) 進行其二級結構預測。
1.2.5 Bmelo424表達載體的構建
通過Bmelo424基因的克隆序列設計分別帶有BamHⅠ和NotⅠ的上下游引物 (表1),采用TA克隆的方式首先構建pMD-Bmelo424重組質粒。將pMD-Bmelo424質粒和穿梭載體pYES2同時用BamHⅠ和NotⅠ雙酶切,酶切產物經1%瓊脂糖凝膠電泳后,分別切膠回收Bmelo424和pYES2目的DNA片段。回收產物用T4 DNA連接酶過夜連接,連接反應體系為:Bmelo424片段7 μL,pYES2片段1 μL,10×T4緩沖液1 μL,T4 DNA連接酶1 μL。連接產物轉化大腸桿菌感受態(tài)細胞,采用菌液PCR篩選陽性克隆并進行酶切和測序雙重驗證。
1.2.6 pYES2-Bmelo424的誘導表達
依據酵母化學感受態(tài)細胞制備試劑盒操作說明,將pYES2-Bmelo424和pYES2分別轉化釀酒酵母INVSc1感受態(tài)細胞,把菌液涂布在含2%葡萄糖的SC-Ura固體選擇培養(yǎng)基中,30 ℃恒溫培養(yǎng)48 h,挑取單菌落做菌液PCR進一步驗證陽性轉化子。
分別挑取陽性轉化單菌落接種于10 mL含2%葡萄糖的SC-Ura液體選擇培養(yǎng)基中,30 ℃、200 r/min振蕩培養(yǎng)并測定OD600吸光度。計算并取出相應體積菌液 (參照酵母表達載體pYES2使用手冊),3 000 r/min離心5 min,然后將菌體重懸于50 mL含2%半乳糖和1%棉籽糖的SC-Ura液體誘導培養(yǎng)基中,使得初始菌液OD600為0.4,30 ℃、200 r/min振蕩培養(yǎng)30 h后,3 000 r/min離心5 min收集菌體,無菌水洗滌菌體3次,真空冷凍干燥48 h,置于-80 ℃冰箱備用,每個樣品做3個重復。
1.2.7 重組產物的GC-MS分析
脂肪酸提取及甲酯化依據文獻略有調整[30]:稱取50 mg釀酒酵母凍干粉于10 mL離心管,加入3 mL氫氧化鈉甲醇溶液 (1 mol/L),振蕩混勻,80 ℃水浴20 min,水浴期間多次振蕩混勻。甲酯化:往樣品中加入3 mL鹽酸甲醇溶液 (2 mol/L)混勻,然后80 ℃水浴20 min,快速冷卻。萃取凈化:待上述溶液冷卻,加入1 mL正己烷 (含0.01%BHT),充分振蕩萃取。離心,取1 μL正己烷層上機測定,使用儀器為安捷倫氣相色譜質譜聯用儀 (Agilent7890-5975)。
色譜條件:所用色譜柱為安捷倫HP-5MS(60 m×0.25 mm×0.25 μm)。進樣口溫度為280 ℃,進樣量為1 μL,分流比為20∶1,載氣為氦氣,恒線速度流速為1.5 mL/min;升溫程序:初始柱溫為120 ℃,保持1 min;以6 ℃/min的速率升到170 ℃;再以2 ℃/min的速率升到230 ℃,保持12 min;最后以10 ℃/min的速率升到285 ℃,保持10 min。
質譜條件:離子源溫度為200 ℃,四級桿溫度為150 ℃,傳輸線溫度為260 ℃,電子轟擊能量為70 ev,采用全掃描模式,掃描范圍m/z40–550,掃描速率為2.83次/s,溶劑切除時間為4.4 min。
通過檢索NIST11譜圖庫并結合標準品圖譜和各組分的保留時間來定性脂肪酸,再按照峰面積歸一法計算其百分含量。
1.2.8 釀酒酵母溫度脅迫處理
將1.2.6中用誘導培養(yǎng)基振蕩培養(yǎng)30 h后的菌液分別取出5 mL,調整pYES2-Bmelo424和pYES2菌液密度至OD600值相同,分別在4 ℃和40 ℃脅迫處理24 h,然后對處理后的菌液分別作10倍梯度稀釋,最后各取2 μL接種在含2%葡萄糖的SC-Ura固體培養(yǎng)基上,30 ℃培養(yǎng)48 h,觀察釀酒酵母受溫度脅迫后的生長狀況。
2 結果與分析
2.1 Bmelo424基因的克隆與分析
以家蠶品種Dazao五齡三天幼蟲整蠶的cDNA為模板,用Bmelo424基因的克隆引物進行PCR擴增,PCR產物的大小采用1%瓊脂糖凝膠電泳檢測,電泳結果顯示有600 bp左右的特異性條帶,和預期片段大小相同 (圖1A)。對該PCR產物進行切膠回收,連接轉化pMD19-T載體,篩選出陽性克隆進行測序,得到558 bp的開放閱讀框 (ORF) 序列,編碼185個氨基酸,具備高度保守的組氨酸簇HXXHH (圖1B),比對結果表明Bmelo424克隆序列和家蠶SilkDB數據庫預測基因序列完全一致。
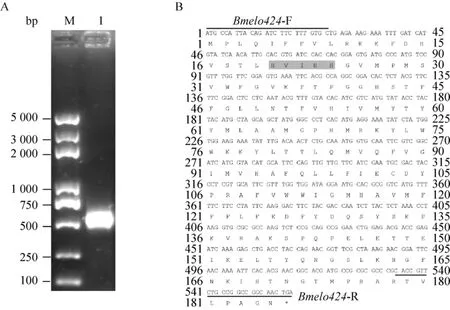
圖1 家蠶Bmelo424基因的克隆 (A) 及分析 (B)Fig.1 Cloning and analysis of Bmelo424 gene.(A) PCR amplification of the Bmelo424 gene.(B) The ORF sequence of Bmelo424 gene and its deduced amino acid, HXXHH motif was highlighted in grey.
2.2 Bmelo424蛋白序列分析
通過TMHMM Server 2.0工具在線預測Bmelo424蛋白的跨膜區(qū)域,結果顯示其有4個跨膜區(qū),預測位置分別在13–35、45–67、79–101、105–124的氨基酸區(qū)域 (圖2A)。利用NetPhos 3.1 Server預測Bmelo424的磷酸化位點有:6個絲氨酸 (Serine) 磷酸化位點 (分別在第30、43、131、133、141和204 位點)、8個蘇氨酸 (Threonine) 磷酸化位點 (第38、44、81、82、148、149、173和179位點)、4個酪氨酸 (Tyrosine) 磷酸化位點(第61、128、132和156位點) (圖2B)。通過PSIPRED v3.3工具在線分析其二級結構,其中,無規(guī)則卷曲所占比例最大,為53.3%,α-螺旋和β-折疊股分別占26.7%和20% (圖2C)。蛋白質亞細胞定位預測結果顯示Bmelo424蛋白位于內質網中。
2.3 Bmelo424基因的組織表達特征
采用熒光定量PCR的方法對Bmelo424基因在五齡三天家蠶幼蟲組織的表達模式進行分析,結果顯示,其在精巢、卵巢、頭、體壁、脂肪體、中腸、血細胞、前中絲、后絲和馬氏管中均有表達 (圖3)。其中,在頭和體壁中表達量較高,在精卵巢中表達量稍低,在血細胞中表達量最低。
2.4 重組質粒的構建和鑒定
用帶有酶切位點的引物PCR擴增Bmelo424基因 (表1),通過TA克隆獲得pMD-Bmelo424重組質粒,將該重組質粒與pYES2質粒用BamHⅠ和NotⅠ分別雙酶切后進行連接和轉化,篩選陽性質粒并進行雙酶切鑒定 (圖4),陽性質粒酶切后出現目的基因大小的片段,同時雙酶切后的載體片段與空質粒pYES2的大小一致,而且陽性質粒測序結果也和克隆序列相同,說明成功構建pYES2-Bmelo424表達載體。
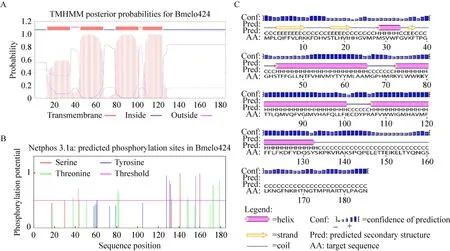
圖2 Bmelo424蛋白序列分析Fig.2 Protein sequence analysis of Bmelo424. (A) Transmembrane domain prediction of Bmelo424.(B)Phosphorylation sites prediction of Bmelo424.(C) Secondary structure prediction of Bmelo424.
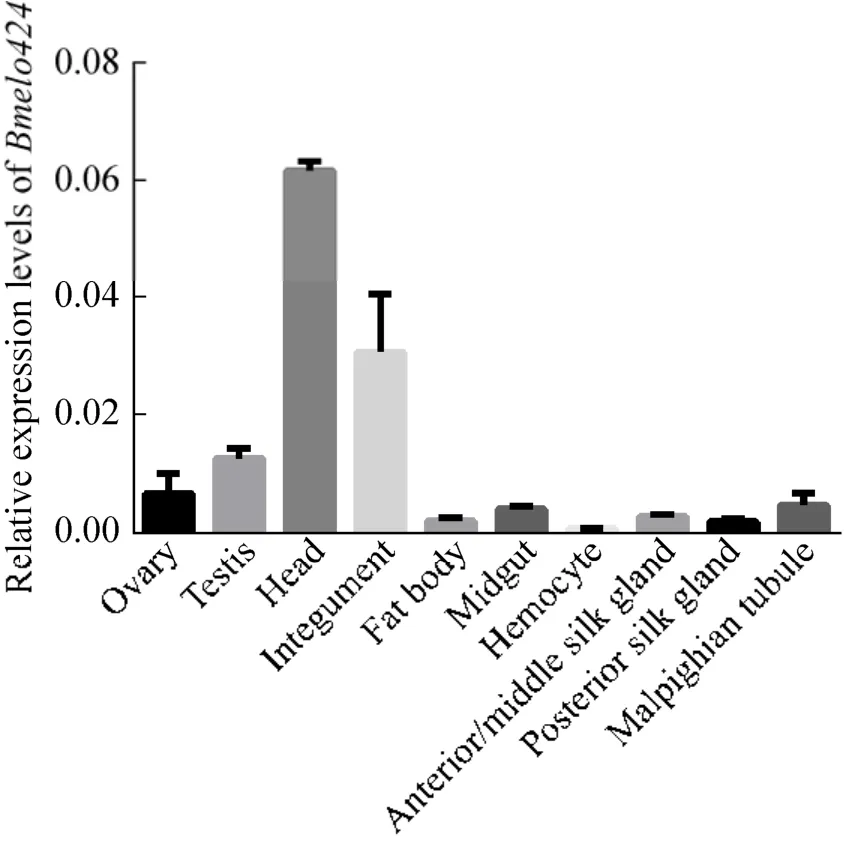
圖3 Bmelo424基因的組織表達譜Fig.3 Expression profiles of Bmelo424 gene in different tissues.
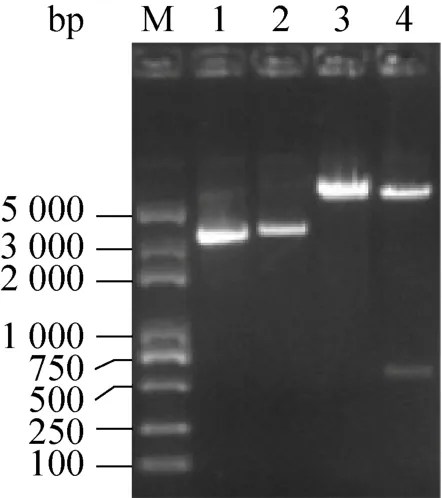
圖4 pYES2-Bmelo424重組質粒的雙酶切鑒定Fig.4 Restriction enzyme digestion of the recombinant pYES2-Bmelo424.M: marker; 1: pYES2; 2:pYES2-Bmelo424; 3: pYES2 plasmid digested with BamHⅠand NotⅠ; 4: pYES2-Bmelo424 plasmid digested with BamHⅠand NotⅠ.
2.5 重組質粒轉化釀酒酵母細胞
釀酒酵母INVSc1屬于營養(yǎng)缺陷型菌株,只有在pYES2存在的情況下,才能在尿嘧啶缺陷型培養(yǎng)基(SC-Ura) 上生長。提取構建好的pYES2-Bmelo424重組質粒和pYES2空質粒,轉化釀酒酵母INVSc1感受態(tài)細胞,涂布培養(yǎng)后在SC-Ura培養(yǎng)基上長出單菌落。挑取單菌落進行菌液PCR反應,得到與目的基因大小相符的片段,進一步驗證重組質粒成功轉化至釀酒酵母INVSc1細胞中 (圖5)。
2.6 重組質粒誘導產物的鑒定
將含有空質粒和重組質粒的釀酒酵母分別接種SC-Ura液體培養(yǎng)基,過夜培養(yǎng)后收集適量菌體,再把菌體重懸于誘導培養(yǎng)基中振蕩培養(yǎng)30 h,離心收集菌體后冷凍干燥,并對培養(yǎng)物進行脂肪酸提取和甲酯化,利用GC-MS檢測其脂肪酸組分。結果顯示,攜帶空質粒pYES2的釀酒酵母細胞中主要含有棕櫚酸 (C16:0)、棕櫚油酸 (C16:1n-7)、硬脂酸 (C18:0) 和油酸 (C18:1n-9) 4種脂肪酸。而攜帶pYES2-Bmelo424重組質粒的酵母細胞中,主要脂肪酸含量發(fā)生變化,除了棕櫚油酸含量顯著上升外,其余3種脂肪酸的含量都有所下降 (圖6)。
2.7 轉基因釀酒酵母的溫度脅迫響應
釀酒酵母在正常溫度 (30 ℃) 培養(yǎng)時,攜帶pYES2-Bmelo424重組質粒和pYES2空質粒的細胞的生長速度和菌落形態(tài)沒有明顯差別。而不同溫度脅迫處理后的兩種釀酒酵母在細胞生長活性上表現出差異,其中,低溫 (4 ℃) 處理后,攜帶pYES2-Bmelo424重組質粒的釀酒酵母細胞比攜帶pYES2空質粒的釀酒酵母細胞生長狀況要好,但是高溫(40 ℃) 處理后,其生長狀況卻截然相反 (圖7)。
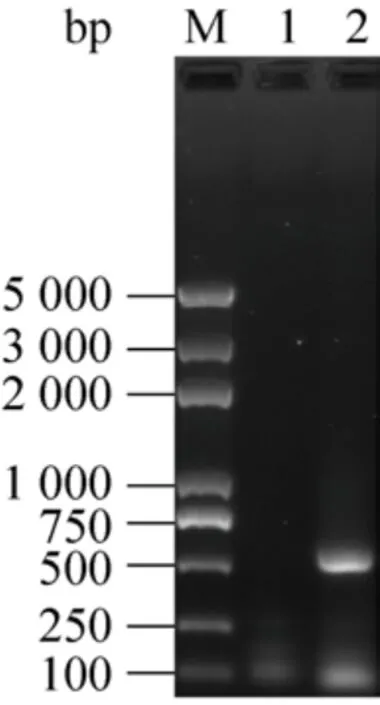
圖5 酵母轉化子的PCR鑒定Fig.5 Identification of yeast transformants by PCR.M:marker; 1: pYES2; 2: pYES2-Bmelo424.
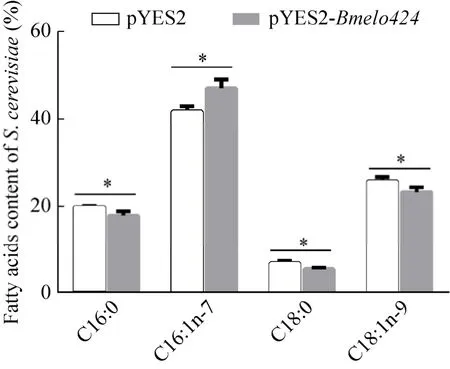
圖6 pYES2酵母細胞和pYES2-Bmelo424重組酵母細胞的脂肪酸含量比較Fig.6 Comparison of fatty acids content between pYES2 and pYES2-Bmelo424 of yeast cells.*P<0.05.
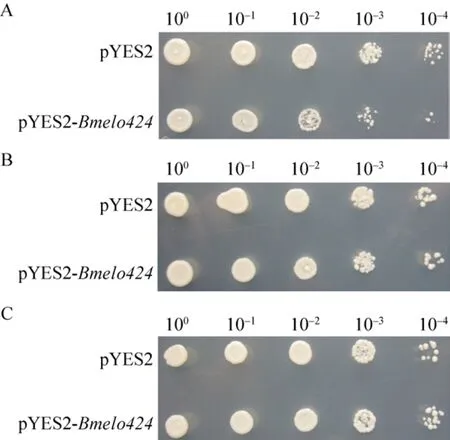
圖7 兩種酵母細胞在不同溫度脅迫處理后的生長狀況比較Fig.7 Comparison of the growth of two kinds of yeast cells after different temperature stress treatments.(A)40 °C.(B) 30 °C (Control).(C) 4 °C.
3 討論
ELOVL基因突變會引起諸如魚鱗癬、黃斑退化、肌病、智力缺陷和髓鞘脫失等遺傳性疾病[31],昆蟲中ELOVL基因具有影響信息素合成、雄性生育力和Toll/Imd免疫信號傳導調節(jié)等功能[19,23-24]。家蠶是典型的鱗翅目經濟昆蟲,其ELOVL基因多達13個,但還沒有功能鑒定報道[28]。本文以家蠶五齡三天全蠶cDNA為模板克隆獲得Bmelo424基因558 bp的ORF序列,其推導氨基酸序列具有4個潛在的跨膜結構域,高度保守的組氨酸簇HXXHH,蛋白質亞細胞定位預測顯示其位于內質網中,表明Bmelo424和其他延長酶一樣屬于膜結合蛋白,但其主要功能發(fā)揮區(qū)域同樣局限于內質網中[32],另外,其具備發(fā)揮活性所必需的HXXHH基序,并且其活性位點和賴氨酸殘基之間的距離決定超長鏈脂肪酸的碳鏈合成長度[33]。Bmelo424有6個絲氨酸磷酸化位點、8個蘇氨酸磷酸化位點和4個酪氨酸磷酸化位點,這些磷酸化位點可能對其生物學功能行使有重要影響。蛋白質二級結構預測結果顯示α-螺旋和β-折疊股占46.7%,二者常位于內部且不易變形,可穩(wěn)定維持蛋白質的高級結構。Bmelo424基因在每個組織中均有表達,這與芯片數據基本一致[28]。在昆蟲類超長鏈脂肪酸延長酶家族成員中,既有在各組織中廣泛表達的成員[23,26],也有組織特異性表達的成員[19],Bmelo424基因在家蠶各組織均有表達,而其表達水平的高低暗示該基因在不同組織中行使的功能可能有所差異。此外,鑒于昆蟲表皮脂類在失水屏障、影響化學農藥吸收、阻止病原入侵以及信息素組成等方面的作用[34],結合Bmelo424基因主要在頭和體壁高表達的結果,我們推測其可能與家蠶的環(huán)境適應性相關。
Bmelo424基因的異源表達會引起釀酒酵母細胞的脂肪酸組分含量的變化,pYES2-Bmelo424重組酵母比pYES2酵母的C16:1n-7脂肪酸含量有顯著提高,表明外源Bmelo424基因在釀酒酵母細胞中對催化生成C16:1n-7脂肪酸有積極作用。家蠶Bmelo424基因僅影響長鏈脂肪酸的合成,值得一提的是,ELOVL參與合成不同碳鏈長度脂肪酸的功能在其他物種中也有報道,諸如酵母中ELO1限于延伸C14脂肪酸到C16[2];錐蟲中ELO1延伸C4脂肪酸到C10,ELO2延伸C10脂肪酸到C14,ELO3延伸C14脂肪酸到C18[35];小鼠中ELOVL6參與延伸C12–C16脂肪酸[14];果蠅中ELO68α可以合成C16和C18脂肪酸[19];黃粉蟲中TmELO2可以合成C16脂肪酸[27]。這些研究結果都說明ELOVL除了主要負責超長鏈脂肪酸的延伸外,也參與其他碳鏈長度的脂肪酸延伸。同時,我們將昆蟲中已涉及功能實驗的ELOVL成員作了比較,發(fā)現Bmelo424與黃粉蟲的TmELO2親緣關系最近,且黃粉蟲的TmELO2也僅影響C16脂肪酸合成[27],因此,家蠶Bmelo424與黃粉蟲中已報道的同系物功能基本吻合。
在果蠅中,其耐冷性增強后會導致單烯酸比例增加而二烯酸比例降低,同時伴有C16脂肪酸增加而C18脂肪酸減少的現象[36],Bmelo424基因引起酵母C16:1n-7脂肪酸含量提高可以增加酵母細胞的冷脅迫后的適應能力,但對熱脅迫后的適應能力卻明顯減弱,這也間接暗示Bmelo424基因可能影響家蠶的抗逆性。另外,白紋伊蚊中ELO表達水平的上調和滯育卵抗干燥能力的提高相關,也說明ELOVL家族的部分基因影響其抗逆性[24]。這也提示我們可以通過改變ELOVL抗逆相關基因的表達,以提高經濟昆蟲適應逆境的能力,或用類似方式降低害蟲適應逆境的能力以達到控害目標。
