慢病毒干擾水通道蛋白4促進脊髓損傷修復與其上調電壓依賴性陰離子通道2相關性的研究*
2019-02-17 07:43:44張華陽黃守娟向軒頤張奇蘭
成都醫(yī)學院學報 2019年6期
呂 芾,張華陽,黃守娟,向軒頤,鄭 莉,張 曉,張奇蘭
成都醫(yī)學院(成都 610500)
脊髓損傷屬于中樞神經(jīng)系統(tǒng)的嚴重創(chuàng)傷,大部分是由于各種外力作用于脊柱,造成脊髓壓迫或斷裂,常導致不可恢復的感覺及運動功能部分或完全喪失[1]。由于脊髓損傷難以恢復,常將之視為一種嚴重的致殘性疾病,其給社會和家庭帶來了很大負擔。脊髓損傷分為原發(fā)性與繼發(fā)性,脊髓水腫是脊髓損傷后的主要并發(fā)癥之一,是其他繼發(fā)性脊髓損傷的病理學基礎,可以極大地影響創(chuàng)傷后的恢復過程。線粒體功能障礙常為導致細胞水腫加劇的關鍵性事件,當線粒體能量的產(chǎn)生和利用受到抑制,將進一步增加細胞內滲透壓負荷,加劇細胞水腫。研究[2-3]發(fā)現(xiàn),脊髓水腫的形成和消退與水通道蛋白(aquaporins,AQP)表達水平有關。而電壓依賴性陰離子通道(voltage-dependent anion-selective channel protein,VDAC)作為線粒體外膜上的一種陰離子通道蛋白,在線粒體能量代謝過程中起重要作用。因此,本課題擬探討AQP與VDAC的相互作用關系,進一步從能量代謝角度研究損傷性脊髓水腫的分子生物學機制。
AQP家族是一類在細胞膜表達的特殊通道蛋白,負責水和其他小分子運輸。目前,AQP已發(fā)現(xiàn)13個亞型,其中AQP4在大腦和脊髓中分布廣泛,是中樞神經(jīng)系統(tǒng)調節(jié)水含量的主要分子途徑[4-5]。AQP4 表達上調與中樞神經(jīng)系統(tǒng)膠質細胞的水腫密切相關,而當其表達下調時可以減輕腦和脊髓水腫的程度,進而促進損傷修復[6-7]。VDAC家族作為一種線粒體外膜上分子和離子交換出入的通道,對線粒體與胞質之間的物質代謝和能量代謝起著十分重要的作用[8]。其中,VDAC2作為其家族成員之一,是線粒體外膜上含量豐富的陰離子通道蛋白,對線粒體和細胞功能的調節(jié)非常重要[9]。當VDAC2表達減少,會使線粒體膜內外代謝物質交換減少,由此導致的ATP產(chǎn)生及利用障礙會影響線粒體發(fā)揮正常功能。
尚未有研究將AQP4與VDAC2對細胞水腫的調節(jié)作用聯(lián)系起來,或探討過二者間的相互作用機制。本實驗制備SD大鼠脊髓鈍挫傷(SCC)模型,采用免疫組織化學(IHC)技術、定量聚合酶鏈式反應(PCR)技術,觀察脊髓損傷后VDAC2表達變化情況;蘇木精-伊紅(HE)染色、BBB 功能評分用于檢測脊髓損傷后組織和細胞的損傷程度以及大鼠運動功能的受損情況;運用生物信息學方法預測AQP4和VDAC2的基因聯(lián)系;構建慢病毒干擾AQP4大鼠模型,觀察AQP4干擾后SCC模型大鼠體內VDAC2的表達變化;并通過以上步驟探討干擾AQP4對脊髓損傷過程中VDAC2表達的影響以及VDAC2的差異性表達在脊髓損傷修復過程中的作用。
1 材料與方法
1.1 實驗動物
健康成年SD大鼠114只,雌雄不限,體重200~240 g。由四川大學實驗動物中心提供。將動物分籠飼養(yǎng),環(huán)境溫度控制在20~25 ℃。隨機把大鼠分為4大組,即假手術(Sham)組(n=11)、SCC組(n=69),取材時間點分為6、12 h和1、3、5、7、14、28 d;空載(Vector)組(n=17)、AQP4慢病毒干擾(AQP4-SH-LV)組(n=17),取材時間點為手術后3、7、28 d。
1.2 AQP4慢病毒質粒載體的制備
1.2.1 慢病毒質粒的制備 將AQP4 siRNA序列(中國廣州,GeneCopoeia)克隆到紅色熒光蛋白(red fluorescent protein, RFP)標記的載體中。再將該載體(5 μg)和慢病毒包裝質粒 (1 μL, 中國廣州,GeneCopoeia)共轉染到293T細胞,生成慢病毒質粒載體。48 h后取含有慢病毒質粒的上清液,置于0.45 μmol/L的醋酸纖維素過濾器中過濾,除去濾液后取5 mL,3 500 r/min,離心半徑28.5 cm,離心25 min,除去上清液,將沉淀重新溶解在500 μL PBS溶液中。最后置于-80 ℃冰箱凍存。
1.2.2 載體注射 使用3.6%水合氯醛對大鼠進行腹腔麻醉(30 mg/kg),備皮后采用俯臥位固定,在T9~T11的位置沿背部正中段縱向依次切開,分離皮膚、筋膜、肌肉;剝除T10的棘突及椎板,充分暴露該節(jié)段脊髓,使用腦立體定位儀(DW-5,成都泰盟)于T10節(jié)段脊髓擬打擊點左右兩側各注射慢病毒質粒載體5 μL(AQP4-SH-LV組)或含有空載體的質粒 5 μL(Vector組)。朝向大鼠尾側進針,進針角度與水平面呈45°,進針深度3~5 μm。注射完畢后依次縫合肌肉、筋膜及皮膚,常規(guī)消毒及抗感染處理后,獨籠飼養(yǎng)。
1.3 動物模型制備
慢病毒載體注射48 h后, AQP4-SH-LV組和Vector組大鼠使用3.6%水合氯醛腹腔注射麻醉(30 mg/kg),常規(guī)消毒。俯臥位固定,依次拆下皮膚、筋膜、肌肉的縫合線;SCC組及Sham組大鼠使用同法麻醉,備皮,常規(guī)消毒,以 T10為中心,縱行切開皮膚及皮下組織,分離椎旁肌肉,暴露棘突與椎板;Sham組大鼠僅咬除T10椎板后,縫合傷口,其余各組大鼠則采用改良Allen′s打擊法,在咬除T10椎板后,用Allen′s打擊儀從30 mm高處自由墜落,打擊對應節(jié)段脊髓,大鼠出現(xiàn)脊髓充血和擺尾,代表損傷成功。打擊成功后,逐層縫合傷口,術后各組大鼠腹腔注射青霉素1 mL(1 600 000 U/mL)、5%葡萄糖氯化鈉溶液2 mL,小心護理,獨籠飼養(yǎng)。每天早晚擠壓大鼠膀胱協(xié)助排尿各1次,至大鼠恢復反射性排尿。護理時注意觀察皮膚有無壓瘡或感染、下肢有無潰爛。室溫維持 25~30 ℃,正常飲食。
1.4 大鼠行為學測試
采用BBB評分體系于術后1、3、5、7、14 d對各組大鼠進行后肢運動功能評分,包括0~21分,功能完全缺失為0分,完全正常為21分。評分由3位以上熟悉評分標準的非本組實驗人員進行,最后選取3個人的有效評分取算數(shù)均值。
1.5 取材和切片制備
各組大鼠在術后根據(jù)需要在不同時間點進行取材。深度麻醉大鼠,若用于qRT-PCR,則自左心室插管后剪開右心耳,快速灌注0.9% NaCl,直接取1 cm損傷段脊髓,放入無RNA酶的EP管中-80 ℃保存。若用于形態(tài)學檢測,深度麻醉大鼠后,自左心室插管,剪開右心耳,快速灌注PBS緩沖液,再灌注4%多聚甲醛。打開椎弓管暴露脊髓段,切取損傷點前后1 cm的脊髓組織,放入4%多聚甲醛固定,48 h后取出固定好的脊髓組織進行常規(guī)脫水、包埋以及切片(切片厚度為5 μm,每切5張選用1張)、攤片、貼片,37 ℃烘片后備用。
1.6 HE染色法
選用制備好的脊髓切片,65 ℃烤片2 h,染色流程:1)二甲苯(Ⅰ)10 min;2)二甲苯(Ⅱ)10 min;3)100% 水化乙醇(Ⅰ)5 min;4) 100% 水化乙醇(Ⅱ)5 min;5)95% 水化乙醇(Ⅰ)5 min;6) 95% 水化乙醇(Ⅱ)5 min;7)80% 水化乙醇5 min;8)RO水5 min;9)蘇木素液染色5 min;10)RO水漂洗5 min;11)0.5% 鹽酸乙醇分色8 s;12)自來水緩水流沖洗15 min返藍;13)伊紅液染色7 min;14)RO水漂洗2 min;15)80% 乙醇3 min;16)95% 乙醇(Ⅰ)1 min;17)95% 乙醇(Ⅱ)3 min;18)100% 無水乙醇3 min;19)二甲苯(Ⅰ)5 min;20)二甲苯(Ⅱ)5 min;21)中性樹膠封片。
1.7 免疫組織化學染色
選用制備好的脊髓切片,65 ℃烤片2 h。用免疫酶組化SP法進行染色:與HE染色同法脫蠟、水化后,放入枸櫞酸鈉溶液中行高壓熱抗原修復,0.01 mol/L PBS漂洗,用3%H2O2,37 ℃,孵育15 min,封閉內源性過氧化酶,PBS漂洗,使用5%正常山羊血清37 ℃孵育15 min封閉非特異性抗原,加一抗(1∶100)4 ℃放置18 h,取出切片,室溫下放置20 min,PBS漂洗,加入二抗工作液,37 ℃孵育15 min,PBS漂洗,加入辣根過氧化物酶,37 ℃孵育15 min,PBS漂洗,配制DAB-H2O2顯色液(1 mL過濾PBS+5滴0.3%H2O2+DAB粉末),室溫下加DAB-H2O2顯色液反應3~5 min,顯微鏡下充分顯色后,及時用RO水終止反應,蘇木素復染5 min,RO水漂洗,1%鹽酸-酒精分色10 s,自來水返藍,梯度酒精逐步脫水,二甲苯透明,最后使用中性樹膠封片。光學顯微鏡400倍下觀察免疫陽性反應物分布情況并拍照。
1.8 熒光定量PCR
把1 cm脊髓損傷節(jié)段從-80 ℃保存的冰箱中取出,用Trizol(中國北京,TransGen Biotech)勻漿破碎脊髓組織細胞,經(jīng)氯仿、異丙醇、75%乙醇處理后得到總RNA。采用試劑盒 (中國南京,Vazyme biotech)將總RNA逆轉錄成cDNA,置于-20 ℃冰箱保存?zhèn)溆谩TO計大鼠內參β激動蛋白(β-actin)、VDAC2的上下游引物序列和探針序列。從冰箱中取出逆轉錄的cDNA,使用通用試劑盒(中國南京,Vazyme biotech公司)根據(jù)說明進行熒光定量PCR,獲得PCR擴增產(chǎn)物。反應條件:95 ℃ 2 min,95 ℃ 15 s,52 ℃ 20 s,60 ℃ 40 s,循環(huán)45次。以β-actin為內參照,得到S形動力學曲線后讀取Ct值,經(jīng)過人工校正后按2-ΔΔCt計算出相對表達量(表1)。

表1 引物序列信息
注:Tm為DNA解鏈溫度
1.9 生物信息學
采用GeneMANIA生物信息學方法預測AQP4和VDAC2兩基因之間的關系,網(wǎng)址:http://www.genemania.org。
1.10 統(tǒng)計學方法
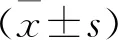
2 結果
2.1 BBB評分
采用BBB評分檢測運動功能,評價大鼠脊髓損傷后的運動功能。Sham組大鼠各時間點評分均達到21分;對于SCC、AQP4-SH-LV、Vector組大鼠,脊髓損傷后1 d其后肢均癱瘓,BBB運動功能評分結果為0,此后隨著后肢功能恢復,其評分逐漸增加,在第14天達到最大值。但在各個時間點(1、3、5、7、14 d),上述3組大鼠BBB評分均低于Sham組(P<0.05); 此外,AQP4-SH-LV組大鼠評分結果在各時間點(1、3、5、7、14 d)均較Vector組高,其中,5、7 d兩組評分差異有統(tǒng)計學意義(P<0.05)(圖1)。

圖1 BBB評分結果
注: 與Sham組比較,*P<0.05;AQP4-SH-LV組與Vector組比較,#P<0.05
2.2 脊髓鈍挫傷后VDAC2的表達變化
2.2.1 SCC后qRT-PCR結果 本實驗采用qRT-PCR技術檢測脊髓損傷后VDAC2的表達變化,結果表明,SCC組和Sham組大鼠在各時間點(6、12 h和1、3、5、7、14、28 d)均能檢測到VDAC2 mRNA轉錄。與Sham組相比,SCC組大鼠各時間點VDAC2的mRNA轉錄量均低于前者(P<0.05)。SCC組大鼠VDAC2 mRNA轉錄水平在脊髓損傷后隨時間的推移而發(fā)生改變,其在脊髓損傷后6 h明顯下降,并在第7天降至最低水平;第7天后其VDAC2 mRNA表達水平緩慢上升,至損傷后第28天,VDAC2的mRNA轉錄量高于第7天,差異有統(tǒng)計學意義(P<0.05)(圖2)。

圖2 qRT-PCR示VDAC2 mRNA的相對表達
注: SCC后VDAC2 mRNA的表達變化;與Sham組比較,*P<0.05;與SCC-7 d組比較,#P<0.05
2.2.2 SCC后IHC結果 采用免疫組織化學技術檢測VDAC2蛋白表達的定位及半定量情況,結果顯示,VDAC2在各組大鼠脊髓灰質神經(jīng)元和神經(jīng)膠質細胞、脊髓白質的星形膠質細胞中有表達,棕褐色免疫陽性產(chǎn)物主要分布于細胞質。SCC術后6 h和1、3、7、14 d,SCC組大鼠免疫陽性細胞數(shù)均低于Sham組(P<0.05),且胞漿內VDAC2陽性反應物明顯減少,提示VDAC2蛋白表達減少;其中,7 d陽性細胞數(shù)最少,14 d較7 d有所增加(P<0.05),但依舊低于Sham組水平(圖3)。

圖3 Sham及SCC組大鼠脊髓組織VDAC2的IHC染色分布(400×)及免疫陽性細胞統(tǒng)計
注: A~F:Sham組及SCC組大鼠脊髓組織VDAC2的IHC染色結果,比例尺=20 μm;黑色箭頭表示免疫反應陽性細胞; G:VDAC2免疫陽性細胞計數(shù)結果;與Sham組相比較,*P<0.05;與SCC-7 d組比較,#P<0.05
2.3 HE染色
Sham組:Sham組大鼠脊髓結構完整,灰白質界限清楚;灰質內神經(jīng)元和神經(jīng)膠質細胞胞體結構完好,分布均勻;血管、中央管形態(tài)正常;白質內神經(jīng)纖維排列整齊而致密。SCC、Vector組:SCC造模后3 d,可見SCC組和Vector組大鼠脊髓組織廣泛出血,如脊髓中央管出血,毛細血管充血,血管周圍有紅細胞;灰質中可見成片壞死區(qū),周圍組織水腫嚴重,細胞間隙和血管間隙明顯增大, 大量神經(jīng)元和膠質細胞腫脹、破裂,細胞核發(fā)生固縮,胞漿紅染加深;有神經(jīng)膠質細胞增生;脊髓灰質和白質界限模糊不清;損傷后7 d: 脊髓組織大部分液化壞死并有空洞形成,脊髓白質破壞嚴重,形成大量囊腔和空泡,神經(jīng)元和膠質細胞腫脹有所減輕, 細胞間隙和血管間隙水腫逐漸減輕,充血消退, 仍有少量增生的膠質細胞。AQP4-SH-LV組:大鼠SCC術后3、7 d,脊髓組織有出血現(xiàn)象,可發(fā)現(xiàn)與Vector組相比,該組大鼠脊髓內血管周圍組織和神經(jīng)元的腫脹程度較輕,水腫體積減小,脊髓灰質和白質的界限較清晰;脊髓中央散在小部分壞死區(qū),程度較輕,白質內大部分髓鞘完整且分布均勻(圖4)。

圖4 HE染色結果顯示各組大鼠脊髓組織情況(400×)
注: A~H:各組大鼠各時間點脊髓組織,比例尺=20 μm
2.4 慢病毒干擾AQP4后VDAC2的表達變化
2.4.1 慢病毒干擾AQP4后qRT-PCR結果 AQP4-RNAi-LV的模式圖如下,qRT-PCR結果顯示,慢病毒干擾后12 h,AQP4-SH-LV組大鼠AQP4的mRNA表達量與Vector組相比下降(P<0.05),證明AQP4干擾模型制備成功;SCC術后7 d, AQP4-SH-LV組大鼠體內VDAC2的mRNA表達量較Vector組大鼠上調,差異有統(tǒng)計學意義(P<0.05)(圖5)。

圖5 慢病毒干擾AQP4后VDAC2的表達變化
注:A:AQP4-RNAi-LV的模式圖,增強型RFP(mCherry FP)作為報告基因被插入到質粒中,該框架也包括抗生素氨芐西林以及作為表達載體啟動子的pUC Ori;B:AQP4-RNAi-LV感染后12 h AQP4 mRNA的表達變化;C:AQP4-RNAi-LV感染后7 d VDAC2 mRNA的表達變化;與Vector組比較,*P<0.05
2.4.2 慢病毒干擾AQP4后IHC結果 Vector組和AQP4-SH-LV組大鼠體內VDAC2免疫陽性產(chǎn)物分布于大鼠脊髓灰質神經(jīng)元和神經(jīng)膠質細胞,脊髓白質的星形膠質細胞胞質中。在SCC術后3、7、28 d,AQP4-SH-LV組大鼠VDAC2的免疫陽性細胞數(shù)相較于Vector組均有所增加,其中,3、7 d兩組差異有統(tǒng)計學意義(P<0.05),28 d時差異無統(tǒng)計學意義(P>0.05)(圖6)。

圖6 AQP4-SH-LV及Vector組大鼠脊髓組織VDAC2的IHC染色分布(400×)及免疫陽性細胞統(tǒng)計
注: A~F:AQP4-SH-LV及Vector組大鼠脊髓組織VDAC2的IHC染色結果,比例尺=20 μm;黑色箭頭表示免疫反應陽性細胞; G:VDAC2免疫陽性細胞計數(shù)結果;與Vector組相比較,*P<0.05
2.5 生物信息學結果
利用GeneMANIA數(shù)據(jù)庫(http://www.genemania.org),檢索時輸入AQP4與VDAC2基因并選擇物種大鼠(rattusnorvegicus),預測AQP4與VDAC2基因在該物種中的聯(lián)系。結果顯示,兩基因在人類和大鼠間均具有共表達(co-expression)、共定位(co-localization)、共享蛋白結構域(shared protein domains)、物理結構互作(physical interactions)、基因互作(genetic interactions)等聯(lián)系。通過蛋白質相互作用預測,結果顯示,VDAC2與AQP4蛋白質均具有底物特異性通道活性、被動跨膜轉運蛋白活性和通道活性等相關作用(圖7)。

圖7 GeneMANIA生物信息學方法檢測VDAC2與AQP4的關系
3 討論
脊髓損傷是一種中樞神經(jīng)系統(tǒng)的嚴重創(chuàng)傷,其導致的脊髓水腫在很大程度上影響脊髓損傷后的修復過程,而脊髓損傷后水腫的發(fā)生發(fā)展過程與線粒體能量代謝息息相關。AQP4和VDAC2作為調節(jié)水代謝和能量代謝的重要因子,對脊髓損傷的修復起著至關重要作用。
3.1 脊髓損傷后VDAC2表達的意義
VDAC2通過在線粒體外膜上形成親水性電壓門控通道,對線粒體外膜透化起門控作用,且因調控物質出入的特性,在能量代謝、維持線粒體穩(wěn)態(tài)等方面也發(fā)揮著不可替代的作用[10]。本實驗成功制備了SD大鼠SCC模型,在脊髓損傷后1 d,SCC組大鼠BBB評分降為0,3 d后隨著運動功能的恢復,評分緩慢增加,但低于Sham組大鼠;脊髓灰質周圍組織水腫嚴重且有大量神經(jīng)元和膠質細胞腫脹、破裂。與此同時,該組大鼠VDAC2的 mRNA表達明顯下調,VDAC2的免疫陽性細胞數(shù)減少,至28 d也未恢復到損傷前水平。上述結果提示,VDAC2的表達水平對維持正常的脊髓功能有重要作用,SCC后其表達下調不利于脊髓損傷恢復。研究[11]發(fā)現(xiàn),VDAC2可能通過凋亡因子影響脊髓損傷后水腫的發(fā)生發(fā)展,Bax、 Bak等是Bcl-2 家族中的促凋亡蛋白。Bax可以透過線粒體外膜使得細胞凋亡,Bax受不斷轉運到線粒體和逆向轉運到胞漿的調節(jié),在VDAC2缺乏的細胞中,Bax和Bak構成的線粒體定位會受到影響,提示VDAC2在Bax和Bak進入線粒體及維持線粒體外膜穩(wěn)定性中起重要作用。 Lauterwasser等[12]進一步發(fā)現(xiàn),線粒體外膜上的孔道蛋白VDAC2為Bax逆向轉運平臺的基本成分,VDAC2具有授予線粒體外膜相關的Bax募集,以及通過逆向轉運來抑制Bax的雙重作用。有研究[13]發(fā)現(xiàn),VDAC2可作為促凋亡蛋白Bak激活的抑制劑。Lazarou等[14]通過實驗發(fā)現(xiàn),在健康細胞中存在內源性依賴VDAC2存在的非活性BAK。在線粒體外膜上,VDAC2與BAK的疏水尾部相互作用,將其隔離在一個非活性狀態(tài),激活的Bak被VDAC2抑制。Pl?tz等[15]結果表明,當Bcl-xs過表達,可干擾VDAC2-Bak的互動,導致釋放Bak,而過表達VDAC2可大幅度降低Bcl-xs導致的細胞凋亡。此外,Wu等[16]發(fā)現(xiàn),VDAC1 和 VDAC2 基因敲除的小鼠,呼吸容量減少,因為線粒體外膜通透性降低,可導致氧化磷酸化所需的代謝物減少。另外,大量研究[17-19]結果顯示,VDAC與鈣離子進出線粒體密切相關,提示VDAC 作為Ca2+進出線粒體的通道,在維持鈣穩(wěn)態(tài)和細胞活性調節(jié)中起關鍵作用。VDAC2還具有胞漿酶的結合位點,包括己糖激酶(HK)和甘油激酶的同工酶,HK是細胞糖酵解過程重要的限速酶之一,VDAC2結合HK-1/HK-2后,可優(yōu)先利用線粒體產(chǎn)生ATP,使糖酵能供順利進行[20]。因為VDAC2控制著多種離子和代謝產(chǎn)物在線粒體內外的轉運,其表達水平可影響許多生理及病理過程。一些實驗[21-23]已表明,VDAC2會影響ATP/ADP進出線粒體,低表達時可抑制線粒體能量的產(chǎn)生和利用,與線粒體的能量代謝密切相關。而細胞線粒體能量代謝障礙時,會發(fā)生ATP生成減少,細胞膜鈉鉀泵功能障礙,細胞內鈉離子和水的過多積聚,導致細胞水腫。無機磷酸鹽、乳酸和嘌呤核苷酸等代謝產(chǎn)物的蓄積,將增加滲透壓負荷,進一步加重細胞水腫,這與本實驗結論相吻合。因此,本實驗認為脊髓損傷后VDAC2表達水平與水腫的發(fā)生發(fā)展直接相關,其表達下調不利于脊髓損傷恢復。
3.2 慢病毒干擾AQP4后可上調VDAC2的表達
AQP是一族構成跨膜的水通道蛋白,主要功能是參與生物體內細胞水分的調節(jié),對維持組織內水平衡具有重要作用。在中樞神經(jīng)系統(tǒng)中廣泛分布的是AQP4,其主要表達在腦和脊髓中的膠質細胞膜或足突和室管膜細胞上,在神經(jīng)元中尚未發(fā)現(xiàn)其蛋白表達。在脊髓損傷后,AQP4表達增加,提示 AQP4高表達提供途徑,使得過量的水分進入脊髓損傷區(qū), 從而引起脊髓水腫、脊髓壓力升高[24]。本項目的前期實驗中,成功建立了AQP4的慢病毒干擾模型,行Allen′s法脊髓損傷后,qRT-PCR和IHC結果顯示,AQP4干擾后VDAC2表達水平上調,差異有統(tǒng)計學意義(P<0.05);本實驗結果顯示,脊髓損傷后手術組大鼠體內VDAC2的表達水平下降,與Sham組比較,差異有統(tǒng)計學意義(P<0.05);AQP4慢病毒干擾組大鼠體內VDAC2的表達相對于Vector組大鼠明顯升高。生物信息學分析結果表明,AQP4與VDAC2具有共表達、共定位和共享蛋白質結構域等關系。因此,本實驗推測AQP4在脊髓損傷后的高表達可能對VDAC2表達有抑制作用,這不僅使得更多水分進入細胞,線粒體腫脹,也使線粒體外膜上的VDAC2表達下降,結果導致線粒體能量的產(chǎn)生和利用受到抑制,進一步增加了胞內滲透壓負荷,加劇細胞水腫,對脊髓損傷后的修復過程起負面影響。
3.3 干擾AQP4促進脊髓損傷的修復與上調VDAC2有關
AQP4廣泛表達于水分子流動活躍的部位,其存在可能起著調節(jié)水代謝的作用,并參與多部位水腫的發(fā)生和發(fā)展。Hiroaki等[25]實驗顯示,AQP4參與腦出血后期(3 d以后) 細胞毒性腦水腫的形成, 增加了細胞膜對分子的通透性, 導致胞內水腫。在外傷性腦水腫大鼠腦中,受傷部位AQP 4的 mRNA 表達明顯高于腦部其他地方[26]。Yang 等[27]通過實驗發(fā)現(xiàn),在腹膜注射去氨加壓素的水中毒模型中, 觀察到AQP4過表達的轉基因小鼠細胞毒性腦腫脹更為快速。此外,F(xiàn)u等[28]通過制備低氧處理大鼠星形膠質細胞模擬低氧腦缺血的模型,也發(fā)現(xiàn)相比于AQP4 敲除型星形膠質細胞,野生型星形膠質細胞會更快速腫脹,并且細胞體積更大。另有一些實驗研究[29]發(fā)現(xiàn),AQP4的表達變化在脊髓損傷時脊髓水腫的形成和消退過程中起重要作用。本實驗結果也顯示,脊髓損傷后,AQP4-SH-LV組大鼠體內VDAC2 mRNA的轉錄水平相較于Vector組大鼠有所上調,并伴隨有大鼠脊髓細胞水腫減輕,運動功能恢復較好的現(xiàn)象。
綜上所述,線粒體能量代謝功能障礙是導致細胞水腫加劇的關鍵性事件。脊髓損傷后,AQP4通過高表達導致細胞水腫,并下調VDAC2的表達水平,干擾了線粒體能量的產(chǎn)生和利用,進一步增加胞內的滲透壓負荷,加劇SCC后的脊髓水腫。人為干擾AQP4的表達,其對VDAC2的抑制作用減弱,VDAC2的表達得以上調,這使得線粒體能量代謝可維持在一定水平,從而減輕損傷后的脊髓水腫,有利于脊髓損傷的恢復。本研究從能量代謝的角度進一步探討了損傷性脊髓水腫發(fā)生發(fā)展的分子生物學機制,為臨床上脊髓損傷的基因療法提供了新的治療靶點。