結腸癌組織中E2F7蛋白表達變化及意義
2019-02-16 01:22:38陳震龍支巧明匡玉庭
山東醫藥 2019年1期
陳震龍,支巧明,匡玉庭
(蘇州大學附屬第一醫院,江蘇蘇州215000)
結腸癌的發生發展主要與遺傳、炎性腸病、低纖維飲食、大量飲酒、吸煙和肥胖等因素有關[1]。因為環境污染、飲食結構不合理、生活壓力大等,近年來結腸癌的發病率呈逐年上升趨勢,全球每年新發病例超過100萬[2]。晚期結腸癌患者5年生存率較低[3],腫瘤復發、遠處轉移是導致患者死亡的主要原因[4]。因此探討結腸癌的轉移、侵襲機制并尋找新的治療靶標非常必要。E2F7是近年來新發現的非典型E2F轉錄因子家族(E2Fs)成員,擁有2個獨特的DNA結合域,而缺乏典型的RB蛋白結合區域[5]。研究發現,E2F7在子宮內膜癌[6]、卵巢癌[7]、皮膚鱗狀細胞癌[8]等多種惡性腫瘤組織中表達上調。miRNA-26a可靶向抑制E2F7表達,以促進急性髓性白血病腫瘤細胞增殖并抑制單核細胞分化[9]。目前E2F7在結腸癌中的相關研究甚少,Liu等[10]發現敲低E2F7表達可以逆轉全反式維甲酸ATRA和miR-3666抑制劑對結直腸癌HCT116細胞惡性行為的聯合作用,抑制腫瘤的生長、侵襲、轉移。本研究探討結腸癌組織中E2F7蛋白表達變化及其與上皮-間質轉化(EMT)的關系。
1 資料與方法
1.1 臨床材料 收集2017年1~12月本院收治的原發性結腸癌患者65例,均接受手術治療,術中取癌組織經病理檢查確診;男34例、女31例,年齡31~82(54.5±10.8)歲;腫瘤高分化22例,中分化15例,低分化28例;TNM分期Ⅰ~Ⅱ期34例,Ⅲ~Ⅳ期31例;淋巴結轉移36例,遠處轉移16例;術前均未接受輔助放化療及靶向藥物治療。另選取其中15例患者的癌旁正常組織(距腫瘤邊緣>5 cm)作為對照,男8例、女7例,年齡34~73(53.0±11.1)歲。兩種組織的患者性別、年齡均有可比性。切取的腫瘤組織與癌旁正常組織標本均用甲醛固定。患者均簽署知情同意書,且本研究獲蘇州大學附屬第一醫院倫理委員會批準。
1.2 結腸癌組織及結腸正常組織中E2F7、E-cadherin及vimentin蛋白檢測 采用免疫組化法。組織標本經固定、石蠟包埋,制成4 μm厚的切片。石蠟切片用二甲苯脫蠟,梯度乙醇水化;用10 mmol/L檸檬酸鹽緩沖液(pH 6.0)進行高壓抗原修復;3% H2O2中浸泡10 min,以消除內源性過氧化物酶活性;1×PBS洗滌后用10%胎牛血清封閉,37 ℃孵育1 h;滴加稀釋好的一抗,4 ℃過夜;1×PBS洗滌后用生物素化二抗處理樣品,二氨基聯苯胺顯像,蘇木素復染,封片。兔抗人E2F7多克隆抗體購于英國Abcam公司,兔抗人EMT相關因子(E-cadherin、vimentin)多克隆抗體購于美國Cell Signaling Technology公司,免疫組化試劑盒和DAB顯色試劑盒均訂購于上海優寧維生物科技股份有限公司,其他試劑和設備均由蘇州大學附屬第一醫院中心試驗室提供。陽性結果判定:在400倍鏡下每張切片隨機選取5個視野進行觀察,每個視野計數約100個細胞,以胞膜、胞質或胞核含棕黃色顆粒判定為表達陽性,并進行評分。細胞染色強度評分:細胞未染色為0分,淡黃色為1分,棕黃色為2分,棕褐色為3分。陽性細胞比例評分:陽性細胞比例<5%為0分,5%~25%為1分,26%~50%為2分,>50%為3分。兩項評分乘積≤2為表達陰性,>2為表達陽性。
1.3 統計學方法 采用SPSS19.0統計軟件。采用Mann-WhitneyU檢驗比較結腸癌組織與癌旁正常組織中蛋白陽性表達率,采用四格表卡方檢驗或精確概率法分析E2F7蛋白表達與臨床病理特征的關系,用Spearman秩相關分析E2F7與E-cadherin、vimentin蛋白表達的相關性。P<0.05為差異有統計學意義。
2 結果
2.1 結腸癌組織及結腸正常組織中E2F7、E-cadherin、vimentin蛋白表達比較 E2F7蛋白主要定位于細胞核中,E-cadherin蛋白主要定位于細胞膜上,vimentin蛋白主要定位于胞質中。結腸癌組織及結腸正常組織中E2F7蛋白陽性表達率分別為78.5%(51/65)、33.3%(5/15),E-cadherin蛋白陽性表達率分別為29.2%(19/65)、73.3%(11/15),vimentin蛋白陽性表達率分別為80%(52/65)、13%(2/15)。與結腸正常組織比較,結腸癌組織中E2F7、vimentin蛋白陽性表達率高(P均<0.05),E-cadherin蛋白陽性表達率低(P<0.05)。
2.2 結腸癌組織中E2F7蛋白陽性表達與患者臨床病理特征的關系 結腸癌組織中E2F7蛋白陽性表達率與腫瘤直徑、淋巴結轉移和TNM分期有關(P均<0.05),與患者年齡、性別及組織分化程度無關(P均>0.05)。見表1。
2.3 結腸癌組織中E2F7蛋白表達與E-cadherin、vimentin蛋白表達的相關性 Spearman秩相關分析結果顯示,結腸癌組織中E2F7蛋白表達與E-cadherin蛋白表達呈負相關(r=-0.39,P=0.001),與vimentin蛋白表達呈正相關(r=0.53,P<0.01)。
3 討論
研究發現,E2Fs參與細胞增殖、分化、凋亡、周期調控和DNA損傷應答過程。迄今為止,已發現E2Fs的8個家族成員(E2F1~8)[11,12]。根據功能,E2Fs分為轉錄激活因子(E2F1~3)和轉錄抑制因子(E2F4~8);根據其結構特點,則可分為典型E2Fs(E2F1~6)和非典型E2Fs(E2F7、8)。目前有關E2Fs成員在腫瘤的發生發展中作用機制的研究已取得不錯進展,部分E2Fs成員在腫瘤診斷與治療等方面的臨床價值已得到肯定[13]。E2F7是近年來新發現的非典型E2Fs成員,與典型E2Fs不同,E2F7缺乏與同分化調控蛋白結合所需的DIM域以及介導基因轉錄的反式激活結構域,取而代之的是2個包含異二聚體殘基的非典型DNA結合域,因此E2F7可通過非DP蛋白依賴的方式與DNA結合,以發揮轉錄抑制作用[14,15]。
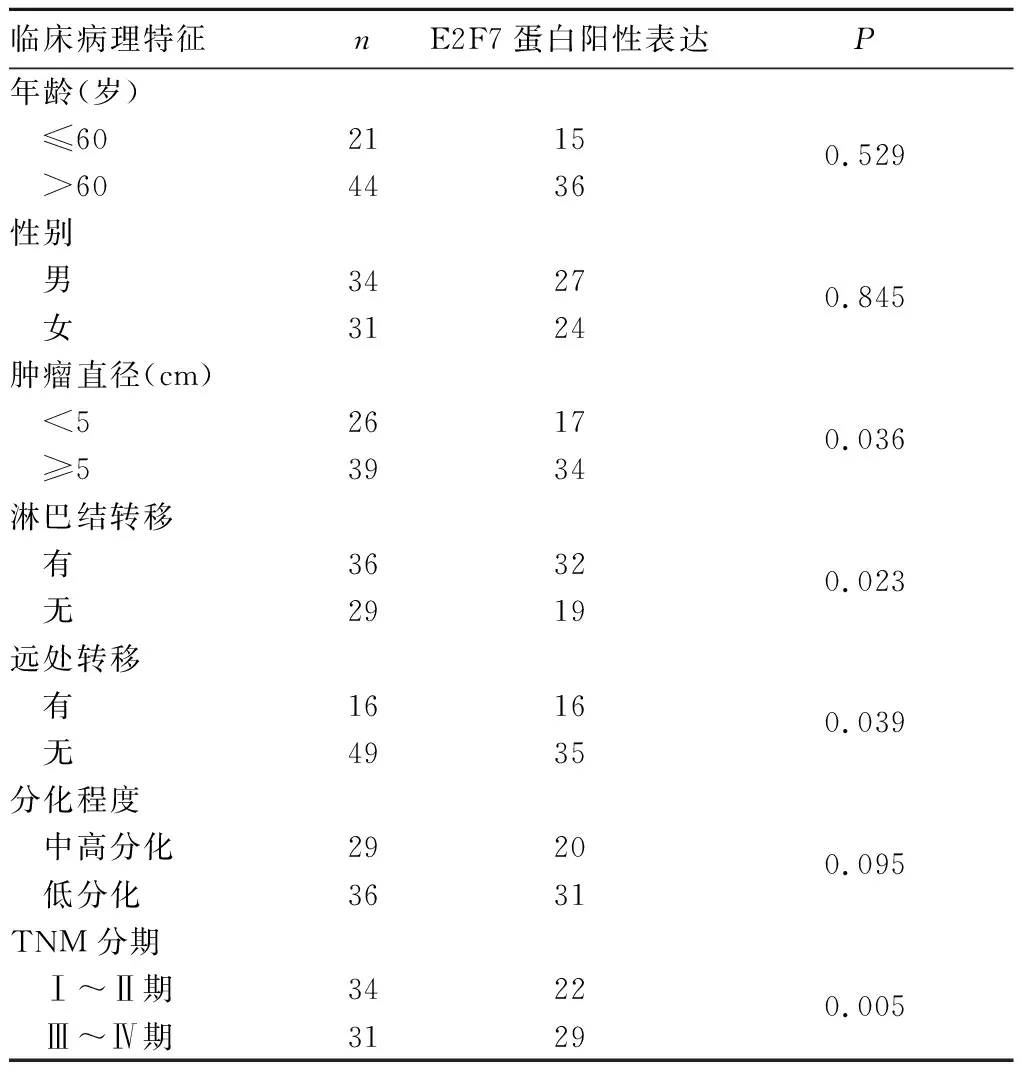
表1 結腸癌組織中E2F7蛋白陽性表達與患者臨床病理特征的關系(例)
有研究認為E2F7可通過抑制增殖相關miRNA的轉錄來抑制細胞的增殖能力[16]。但近年來隨著對E2F7深入研究,發現其在許多惡性腫瘤組織中可能發揮著促癌作用。E2F7在神經膠質瘤組織中表達上調,可作為神經膠質瘤的獨立預后標記物[17]。Chu等[18]研究顯示在乳腺癌中過表達E2F7可以抑制miR-15a/16轉錄,使Cyclin E1和Bcl-2表達上調,進而增強了乳腺癌細胞對他莫昔芬的耐藥性。也有研究表明在非小細胞肺癌組織中,miR-935可通過靶向抑制E2F7表達來抑制腫瘤的轉移和侵襲[19]。在膽囊癌組織中過表達E2F7可促進腫瘤細胞的遷移,而抑制E2F7表達能同時增加E-cadherin表達并降低vimentin的表達,以推動EMT過程[20]。提示E2F7可能參與腫瘤的侵襲和遷移,并影響EMT過程。
EMT是指上皮細胞在某些特定條件下發生轉化,失去細胞極性而獲得侵襲性、運動性、抗凋亡性等間充質細胞表型特征的生物學過程,包含復雜的分子機制[21,22]。EMT不僅是胚胎發育的關鍵機制,還在成人慢性炎癥、組織再生、創傷修復和器官纖維化等生理病理過程中發揮重要作用[23]。研究表明,EMT是腫瘤細胞獲得侵襲能力的關鍵步驟,與腫瘤的轉移和侵襲密切相關[24]。E-cadherin是典型的上皮細胞標志分子,參與維持細胞連接。研究表明E-cadherin缺失是EMT過程的一個早期事件,會導致細胞急性復合體的破壞,這是細胞發生分裂、侵襲的前提[25]。vimentin是只存在于間質細胞中的細胞骨架蛋白,其在各種惡性腫瘤中的表達與腫瘤的侵襲、轉移密切相關[26]。E-cadherin和vimentin都是EMT最具代表性的分子標志物,E-cadherin表達失活和vimentin表達上調是EMT發生的關鍵環節之一[27,28]。
本研究發現,結腸癌組織中E2F7蛋白陽性表達率明顯高于癌旁正常組織,提示E2F7在結腸癌組織中呈高表達;而E-cadherin和vimentin在結腸癌組織中的表達也與EMT基因表達譜相符。E2F7蛋白陽性表達的結腸癌患者,其腫瘤直徑更大、TNM分期更晚,且更容易發生淋巴結轉移。表明E2F7可能具有促進結腸癌生長、轉移、侵襲的作用。為進一步闡明E2F7參與腫瘤轉移和侵襲的作用機制,分析結腸癌組織中E2F7與E-cadherin及vimentin蛋白表達的相關性,結果表明E2F7與E-cadherin蛋白表達呈負相關,而與vimentin蛋白表達呈正相關。提示E2F7參與推動EMT的發生。
綜上所述,E2F7在結腸癌組織中呈高表達,E2F7高表達可能促進結腸癌的生長、轉移和侵襲,其對腫瘤轉移侵襲的促進可能是通過EMT過程來實現的,E2F7可能成為結腸癌治療的新型靶標。但本研究收集的樣本數量有限,需要增加樣本量來證實以上結果,并且E2F7在結腸癌中的作用通路有待進一步闡明。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
西南國防醫藥(2016年7期)2016-12-01 06:01:15
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
