葡萄糖3-脫氫酶工程菌的構建、表達及其合成3-酮對硝基苯井岡霉胺
2019-02-15 08:28:26張建芬
生物加工過程 2019年1期
張建芬,柯 薇,陳 虹
(浙江樹人大學 生物與環境工程學院,浙江 杭州 310015)
3-酮糖類化合物含有特殊羰基基團,是一種重要的合成砌塊,可作為先導化合物,用于選擇性合成氨基糖抗體、去垢劑、糖聚合體、食品添加劑和抗氧化劑等,具有廣泛的應用前景[1]。糖分子含有多個功能相近的羥基,以此為原料,通過化學反應選擇性合成3-酮糖類化合物難度較大,涉及多步的基團保護和去保護反應,而且污染大、能耗高。葡萄糖3-脫氫酶(glucoside 3-dehydrogenase,G3DH,EC.1.1.99.13)是1種以黃素腺嘌呤二核苷酸(FAD)為輔酶的新型氧化還原酶,能氧化吡喃糖上的C-3羥基為酮基[2]。G3DH在3-酮糖類化合物的生物合成中起著重要的作用。目前報道的能生產G3DH的微生物較少,主要有Agrobacteriumtumefaciens[3]、Flavobacteriumsaccharophilum[4]、Halomonas(Deleya)sp.A-15[5]、Cytophagemarinoflava[6]、Agaricusbisporus[7]、Stenotrophomonasmaltrophi-lia[8]和Sphingobacteriumfaecium[9]等,而且,已篩選到的G3DH的酶活性普遍不高,離滿足工業生產的實際需求還有一段距離。開發經濟高效的基因工程菌株,可解決自然進化的酶催化效率低、副產物多等問題,可以有效地提高生物催化劑的性能。目前,有關基因克隆G3DH的研究報道較少,僅有來自A.tumefaciens[10]和Halomonas[11]的G3DHs實現了在E.coli中的表達,而且活力較低或者表達量不高。自然界中蘊藏著豐富的未被發掘的新型酶資源,隨著DNA測序技術的發展和成本的降低,大量微生物基因組信息先后被測定和公開。基因挖掘技術可以從基因組數據庫快速發現新的有價值的酶,進一步豐富可被利用或改造的酶資源[12],已吸引了廣大科研工作者的關注。因此,基因挖掘新的G3DH酶資源,對于提高G3DH的催化反應效率、拓寬G3DH的應用范圍具有重要意義。
3-酮對硝基苯井岡霉胺是3-酮井岡羥胺A C-N裂解酶的酶反應底物,3-酮井岡羥胺A C-N裂解酶是酶解井岡霉素制備井岡胺等糖苷酶抑制劑的關鍵酶之一,具有重要的研究應用價值。3-酮對硝基苯井岡霉胺可通過G3DH生物轉化來制備。Takeuchi等[4]利用F.saccharophilum提取的粗酶液膜蛋白部分轉化,得到了3-酮對硝基苯井岡霉胺,作為C-N裂解酶的底物。但是用酶液轉化生產存在著酶液制備費時費力,收率低等缺點。項目組前期用S.maltrophilia細胞轉化制備3-酮對硝基苯井岡霉胺[13],由于野生菌株同時含有3-酮對硝基苯井岡霉胺裂解酶,會有副產物對硝基苯胺產生,使3-酮對硝基苯井岡霉胺產率下降,也會導致后續產物分離麻煩。隨后,項目組克隆了來自Sphingobacteriumfaecium的G3DH,并用于3-酮對硝基苯井岡霉胺的細胞轉化法生產[14]。盡管G3DH在合成中的應用前景不可忽視,目前所報道的G3DH酶種類非常有限,基因組挖掘這一方法可以快速發現新的有價值的G3DH。
本研究中,筆者通過基因挖掘的方法表達1種來自Glaciecolapolaris的G3DH,并優化其表達條件,并且用此重組G3DH工程菌細胞轉化法生產3-酮對硝基苯井岡霉胺。
1 材料與方法
1.1 菌株、質粒及引物
大腸桿菌(E.coli)DH5α和E.coliBL21(DE3),Novagen公司,克隆質粒pMD19-T、表達質粒pET-28(b),Takara公司。所需引物均由上海生工生物工程有限公司合成。
1.2 主要試劑和儀器
PCR所用試劑,TaKaRa公司;限制性內切酶、T4 DNA連接酶,Fermentas公司;低分子質量標準蛋白、感受態細胞制備試劑盒,上海生工生物工程有限公司;質粒提取試劑盒、PCR產物回收試劑盒、膠回收試劑盒、細菌基因組提取試劑盒,Axygen公司。Bradford蛋白濃度試劑盒,南京建成公司。蛋白胨和酵母粉,OXOID公司。對硝基苯井岡霉胺為實驗室合成,參照文獻[13]的方法。其他試劑均為市售分析純。
酶標儀、蛋白純化儀和鎳離子親和層析柱,美國Bio-Rad公司。
1.3 基因挖掘
在NCBI的“protein”數據庫中,以“glucose 3-dehydrogenase”或“glucoside 3-dehydrogenase”為檢索詞,得到多條不同來源的G3DH序列。采用Clustal W進行G3DH的多序列比對,分析G3DH的保守序列。在ExPASy的ProtParam上,計算G3DH的理論理化特性,排除預測可溶性低的,確定1種來自Glaciecolapolaris的G3DH(命名為GpG3DH)為研究對象。通過jcat對GpG3DH的基因序列進行了在E.coli中表達的密碼子優化。在GpG3DH基因兩端加上NcoⅠ和XhoⅠ的限制酶識別位點后,由上海生工生物工程有限公司合成。
1.4 工程菌的構建
用NcoⅠ和XhoⅠ酶切處理質粒pET-28(b)和GpG3DH,膠回收載體和GpG3DH基因酶切產物,在16 ℃下,T4 DNA連接酶連接過夜,轉化到E.coliBL21(DE3)中,轉化子經菌落PCR驗證和測序驗證后,獲得陽性重組菌株E.coliBL21/pET28b-GpG3DH。
1.5 重組酶的表達純化
將工程菌接種于5 mL含50 μg/mL卡那霉素的LB液體培養基中,于37 ℃、180 r/min振蕩培養過夜。取1 mL培養液轉接于100 mL含50 μg/mL卡那霉素的LB液體培養基中,于37 ℃、180 r/min振蕩培養至OD600為0.8時,向培養物中加IPTG至終濃度為0.1 mmol/L,于16 ℃誘導表達12 h,4 ℃、4 000g離心10 min,收集菌體,并用生理鹽水洗滌2次。稱取10 g濕菌體,加入20 mL 0.1 mol/L Tris-HCl(pH 8.0)緩沖液懸浮細胞,于冰浴中進行超聲波細胞破碎(工作2 s,間隔5 s,工作時間20 min)。4 ℃、8 000g離心30 min,收集上清液作為粗酶液。粗酶液用0.22 μm的濾膜過濾后,利用鎳離子層析柱(Bio-Rad,20 mL)對粗酶液進行一步純化,純化的酶液經超濾脫鹽后用于酶活力測定。
1.6 酶活力測定
在比色皿中加入2.6 mL pH 6.0的0.1 mmol/L的磷酸鹽緩沖液,0.1 mL 0.1 mol/L的葡萄糖溶液,0.2 mL 0.4 mmol/L的2,6-二氯酚靛酚溶液,0.1 mL適當稀釋的酶液,混勻。以蒸餾水做空白調零,在600 nm下測吸光度減少值。每隔30 s測定,直至3 min。
G3DH酶活力單位定義:1個酶活力單位(U)相當于在上述條件下,于25 ℃、pH 6.0條件下,1 min還原1 μmol 2,6-二氯酚靛酚所需的酶量。
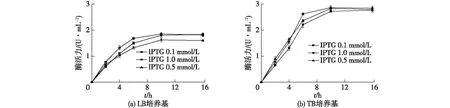
圖2 重組酶的表達條件優化Fig.2 Optimization of expression conditions of recombinant enzyme
1.7 3-酮對硝基苯井岡霉胺的細胞轉化法生產
在50 mL的小三角瓶中,加入25 mg干質量的菌體,10 mL 50 mmol/L的磷酸鹽緩沖液(pH 7.0),充分混合后,加入2 mL 50 mmol/L對硝基苯井岡霉胺,在30 ℃、100 r/min水浴搖床中開始反應。每隔一段時間,取出0.5 mL用于檢測3-酮對硝基苯井岡霉胺的含量。
1.8 底物和產物的分析方法
對硝基苯井岡霉胺、3-酮對硝基苯井岡霉胺和對硝基苯胺的濃度用高效液相色譜法(HPLC)分析。細胞轉化液經12 000 r/min離心10 min后,上清液用0.22 μm的濾膜過濾后,經高效液相色譜分析,采用的是C18反相柱,25%的乙腈水溶液作為流動相,流速為1 mL/min,在398 nm下紫外檢測分析。
2 結果與討論
2.1 表達質粒pET28b-GpG3DH的構建
構建所得重組質粒pET28b-GpG3DH,經NcoⅠ和XhoⅠ酶切后,結果見圖1。由圖1可知,重組質粒中含有目的條帶,約1.68 kb,與預測一致。質粒測序驗證正確,說明重組質粒構建成功。
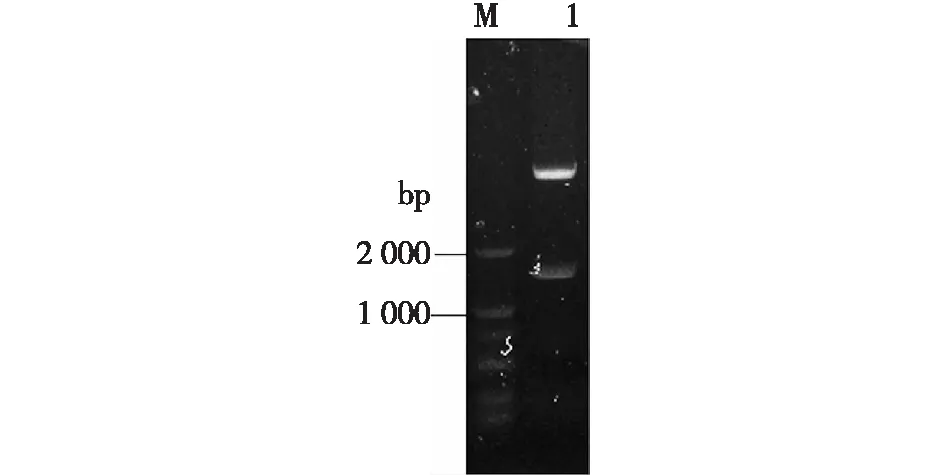
M—標準DNA; 1—酶切質粒DNA圖1 重組質粒pET28b-GpG3DH雙酶切電泳圖Fig.1 Electrophoresis of double digests of recombinantplasmid pET28b-GpG3DH
2.2 重組酶的表達條件優化
在不同的培養基(LB培養基和TB培養基)下,分別加入終濃度為0.10、0.50和1.00 mmol/L的IPTG,16 ℃下誘導表達16 h。每隔2 h取樣分析,結果如圖2所示。由圖2可知,相對于LB培養基,在TB培養基下培養,重組酶的活力更高。此外,IPTG的用量為0.1~1.0 mmol/L時,重組酶的活力影響不大。因此,確定最佳的GpG3DH表達條件:37 ℃培養至OD600為0.8時,加入IPTG至終濃度為0.5 mmol/L,16 ℃誘導表達10 h。在此條件下,GpG3DH的酶活力達到2.83×10-2U/mL。酶活力比已發現的G3DH略低[3-9],后續可以通過定向進化等分子改造手段提高GpG3DH酶活力。
2.3 基因GpG3DH的表達結果
葡萄糖3-脫氫酶基因GpG3DH的表達情況不好,優化表達條件并沒有大幅度增加GpG3DH的可溶表達,表達結果如圖3所示。由圖3可知,G3DH的分子量為5.5×104。據報道,已發現的G3DH的分子量在5.5×104~6.8×104[3-9],GpG3DH的分子量與文獻[3-9]報道一致。于是GpG3DH的催化反應采用E.coilBL21/pET28b-GpG3DH全細胞反應體系。

1,4—對照實驗(不加IPTG誘導);2—菌體破碎液的上清;3—菌體破碎液的沉淀;M為標準蛋白圖3 GpG3DH的SDS-PAGE電泳Fig.3 SDS-PAGE electrophoresis of GpG3DH expression
2.4 3-酮對硝基苯井岡霉胺的細胞轉化法生產
采用重組菌細胞轉化法生產3-酮對硝基苯井岡霉胺。3-酮對硝基苯井岡霉胺的產率Y3-KpNPV用式(1)定義。
Y3-KpNPV=C3-KpNPV/CpNPV,0
(1)
式中:C3-KpNPV表示表示任一時段3-酮對硝基苯井岡霉胺的濃度。
將菌體在不同濃度EDTA(1、5、10、15、20和40 mmol/L)的細胞轉化體系下反應。24 h后檢測對硝基苯井岡霉胺以及3-酮對硝基苯井岡霉胺的含量,結果如圖4所示。
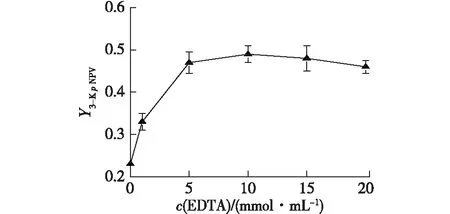
圖4 EDTA濃度對3-酮對硝基苯井岡霉胺生成的影響Fig.4 Effect of EDTA concentration onN-p-nitrophenyl-3-ketovalidamine
從圖4可以看出,隨著EDTA濃度的增加,3-酮對硝基苯井岡霉胺的產率逐漸增加。可見,用EDTA處理細胞,能使目標產物3-酮對硝基苯井岡霉胺的產率大大增加。據文獻[13-14]報道及前期的研究結果,用EDTA處理細胞,能提高G3DH催化糖及其衍生物的轉化率,降低副產物的產率。對于天然酶,EDTA還能抑制Ca2+依賴的3-酮對硝基苯井岡霉胺降解酶活性。本文的研究結果與前期研究結果[13]一致。重組菌E.coilBL21/pET28b-GpG3DH雖不含3-酮對硝基苯井岡霉胺降解酶,但是,EDTA能夠去除外膜蛋白的脂多糖,使得細胞膜的通透性增加[4]。細胞經EDTA處理后,生成的中間產物3-酮對硝基苯井岡霉胺容易及時地到達外環境,這樣就使得被裂解為對硝基苯胺的幾率大大降低。當EDTA濃度大于10 mmol/L時,3-酮對硝基苯井岡霉胺的產率有所下降,其原因可能是高濃度的EDTA存在對酶有抑制作用。因此,確定細胞轉化體系中,最佳的EDTA用量為10 mmol/L。
考察不同的pH條件下,對硝基苯井岡霉胺轉化為3-酮對硝基苯井岡霉胺的產率。菌體先用10 mmol/L EDTA 處理,在30 ℃、100 r/min條件下轉化 24 h,結果見圖5。

圖5 pH對3-酮對硝基苯井岡霉胺合成的影響Fig.5 Effect of pH on N-p-nitrophenyl-3-ketovalidamine
從圖5可以看出,在pH 7.5時,3-酮對硝基苯井岡霉胺的產率最高。因此,確定細胞轉化體系中,pH為7.5。
考察不同的溫度(20、25、30 ℃)下,對硝基苯井岡霉胺轉化為3-酮對硝基苯井岡霉胺的產率。以對硝基苯井岡霉胺和3-酮對硝基苯井岡霉胺的濃度對轉化時間作圖,結果如圖6所示。
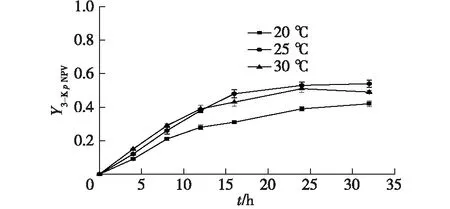
圖6 溫度對3-酮對硝基苯井岡霉胺的合成影響Fig.6 Effects of temperature on N-p-nitrophenyl-3-ketovalidamine
從圖6可以看出,在不同溫度下的曲線形狀大致相同。在轉化初期,隨著溫度的上升,轉化速率呈上升的趨勢,其可能的原因是隨著溫度的升高,細胞膜的通透性增加,使得化學物質容易進入細胞質。因此,對硝基苯井岡霉胺容易通過細胞膜,產物容易通過到達細胞外。轉化12 h后,在30 ℃下,3-酮對硝基苯井岡霉胺的產量增加較緩慢。因此,筆者選擇25 ℃為最佳的3-酮對硝基苯井岡霉胺轉化溫度。在25 ℃、 pH為7.5的反應條件下轉化32 h,3-酮對硝基苯井岡霉胺的產率(Y3-KpNPV)可以達到0.54。在前期研究中[14],S.faecium菌的G3DH的最佳轉化條件為30 ℃、 pH為7.6,與本文的結果相近。
3 結論
采用基因挖掘技術,一種來自于Glaciecolapolaris的葡萄糖3-脫氫酶基因被發現,并進行了GpG3DH基因工程菌的構建和表達。表達質粒pET28b-GpG3DH被成功構建,并在E.coliBL21(DE3)中實現部分可溶表達。經鎳柱純化和電泳檢測,G3DH的分子量為5.5×104。隨后,優化了GpG3DH的表達條件,結果表明,相對于LB培養基,TB培養基能顯著提高G3DH酶活力。重組菌在37 ℃培養至OD600為0.8時,加入IPTG至終濃度為0.5 mmol/L,16 ℃誘導表達10 h,GpG3DH的酶活力達到2.83×10-2U/mL。最后,將重組菌細胞用于轉化對硝基苯井岡霉胺生產3-酮對硝基苯井岡霉胺的研究,最佳的轉化體系為EDTA 10 mmol/L、pH為7.5、25 ℃,轉化32 h,3-酮對硝基苯井岡霉胺的產率(Y3-KpNPV)為0.54。相比較于來自Sphingobacteriumfaecium的重組菌,本實驗3-酮對硝基苯井岡霉胺的產率略低。后續可以通過酶定向進化、定點突變等分子改造手段提高GpG3DH酶活力。
