一株同步硝化-反硝化菌的絮凝特性
2019-02-15 08:31:36梁錫宏李政威吳重德周榮清
生物加工過程 2019年1期
關鍵詞:研究
李 丹,梁錫宏,李政威,金 垚,吳重德,周榮清
(四川大學 輕紡與食品學院 皮革化學與工程教育部重點實驗室,四川 成都 610065)
目前,最常用的脫氮方法是在好氧條件下利用自養菌進行硝化,在厭氧條件下利用異養菌進行反硝化[1]。然而,獨立的硝化反硝化池占地面積大、耗時長、花費高,且操作不便[2]。同時,由于自養硝化菌生長緩慢,易受高濃度的氨、有機物和其他化學元素的影響,使得硝化成為了脫氮過程的限速步驟[3]。近年來,研究人員從環境中分離獲得了異養硝化-反硝化菌[4-6],這些異養菌比自養菌生長更快,能夠利用有機物作為碳源,同時在好氧條件下將含氮物質轉變成N2。在同一個反應器中進行硝化-反硝化可以降低操作費用,提高氮的去除率,而對同步異養硝化-好氧反硝化菌研究的不斷深入,能為更好地實現同步硝化-反硝化提供更多的理論和實踐依據[7]。
活性污泥胞外聚合物(EPS)來源于細胞的分泌溶解、大分子水解以及廢水中吸附的有機物[8],對活性污泥的形成、污泥絮體結構、脫水沉降性能以及重金屬的吸附具有重要的影響[9]。據報道,EPS還會與水體中的各種元素發生生物、化學和物理反應,如配位、吸附、吸收和絮凝反應等,對研究水體與生物之間物質輸送和交換的途徑具有重要影響[10]。此外,胞外聚合物還能提高微生物對重金屬的抗性和吸附[11]。目前,對菌體胞外聚合物的研究多集中在人體口腔微生物[12]和益生菌等腸道微生物[13],而針對硝化-反硝化菌EPS的研究很少。此外,硝化反硝化菌EPS是硝化池和反硝化池中活性污泥能否形成絮凝體的關鍵。
本文中,筆者研究了1株脫氮菌的硝化反硝化性能和自聚集性能,并對其產生絮凝性的胞外聚合物特性進行研究,以期為未來的硝化反硝化菌胞外聚合物研究提供重要參考。
1 材料與方法
1.1 菌株
克雷伯氏菌(Klebsiellasp.TN-10),從制革廢水中篩選得到,保藏于中國典型培養物保藏中心,保藏編號為CCTCC NO:M2017193。
1.2 培養基及菌種培養
硝化培養基(g/L):丙酮酸鈉7、(NH4)2SO40.5、NaCl 1、MgSO40.5、EDTA 0.8、FeSO40.4、K2HPO41;pH 7.0。
反硝化培養基Ⅰ(g/L):丙酮酸鈉7、NaNO30.5、NaCl 1、MgSO40.5、EDTA 0.8、FeSO40.4、K2HPO41;pH 7.0。
反硝化培養基Ⅱ(g/L):丙酮酸鈉7、NaNO20.5、NaCl 1、MgSO40.5、EDTA 0.8、FeSO40.4、K2HPO41;pH 7.0。
從甘油管中將菌種TN-10接種到含50 mL培養基的三角瓶中,在150 r/min、30 ℃條件下培養12 h,將種子液按1%(體積分數)接種量接種到100 mL的上述培養基中,在150 r/min、30 ℃的搖床中進行培養。
1.3 硝化-反硝化能力測定

菌體反硝化性能的測定:將TN-10接種到反硝化培養基Ⅰ和Ⅱ中,培養條件同上,間隔8 h取樣,用同樣的方式處理樣品,測定TN-10在反硝化培養基中的生長和氮的降解情況。
1.4 自聚集性能測定
將活化12 h的種子液按照體積分數1%的接種量接種到硝化培養基中,分別收集培養至對數期、穩定期和衰亡期的發酵液。將樣品在10 000 r/min、4 ℃下離心5 min,去除上清液,并用H3PO4緩沖液(PBS)清洗2次,重懸到PBS中,調整菌體OD600至0.6。將30 mL菌懸液放入50 mL離心管中,每間隔1 h取樣,測定上清液的OD600,菌體的自聚集性計算可以采用式(1)[14]。
自聚集性=(1-At)/A0×100%
(1)
式中:At為t時刻的OD600;t分別為1、2、3、4和5 h;A0為0時刻的吸光值。
菌體的聚集過程可以看成是可逆的化學反應。使用偽一級動力學方程來描述[15],見式(2)。
At=Ae(1-e-tk1)
(2)
式中:k1=h-1;Ae代表平衡時的自聚集性;At表示t時刻的OD600,與式(1)一致。
1.5 胞外聚合物(EPS)的提取與測定
采用陽離子交換樹脂法對菌體胞外聚合物進行提取。將40 mL在硝化培養基中培養12 h的菌體在10 000 r/min、4 ℃下離心5 min,棄上清液,收集菌體。使用超純水將菌體洗滌2遍并重懸到超純水中。將細胞懸液放入250 mL三角瓶中并加入一定量的樹脂(每克干菌體中加70 g樹脂)。將三角瓶放入搖床培養,在150 r/min條件下提取8 h后,離心收集上清液并過0.45 μm濾膜,測定EPS提取液中的蛋白和多糖含量,胞外聚合物的含量以蛋白質和多糖含量表征[16]。多糖的測定采用蒽酮-H2SO4法[17],蛋白質的測定采用改良的BCA法[18]。
1.6 傅里葉紅外光譜分析
將EPS提取液使用旋轉蒸發儀濃縮,在冷凍干燥儀中進行冷凍干燥,得到干燥的固體結晶。使用傅里葉紅外光譜儀對EPS組成進行分析。干燥的EPS與溴化鉀按1∶ 100(質量比)混勻,在瑪瑙研缽中研磨,并放入壓片機進行壓片后,用Nicolet380型傅里葉紅外光譜儀(美國THERMO Fisher Scientific)對樣品進行掃描,分辨率為0.4 cm-1,測定波數范圍為4 000~400 cm-1。
1.7 蛋白質二級結構的測定與分析
使用J-715型圓二色光譜儀(JASCO公司)對EPS溶液進行掃描。掃描范圍為190~250 nm,在1 mm石英比色皿中以超純水做空白,每間隔1 nm記錄一次數據。使用CD pro程序包中的SELON和CONTIN處理CD譜數據,計算蛋白質二級結構[19]。
2 結果與討論
2.1 TN-10的硝化-反硝化能力

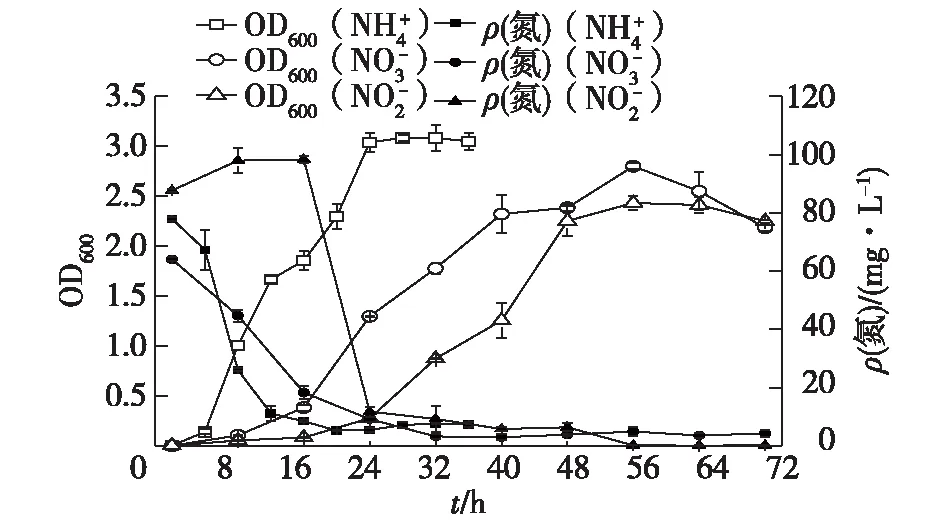
圖1 TN-10的硝化-反硝化性能Fig.1 The nitrifying-denitrifying characteristics of TN-10

2.2 自聚集性能
為了解菌體的自聚集性能,筆者測定了菌體的生長曲線和在不同生長周期的自聚集性能變化情況,結果如圖2所示。由圖2(a)可知:TN-10從4 h起進入對數期,在24 h到達穩定期,從32 h開始進入衰亡期。收集處于12 h(對數期)、24 h(穩定期)和36 h(衰亡期)的菌體,測定菌體的自聚集率。由圖2(b)可知:TN-10表現出較好的自聚集性,其自聚集率達到20%左右,但在不同時期,菌體的自聚集性無顯著差異。隨著培養時間的延長,菌體的自聚集性呈現緩慢增加的趨勢,到達衰亡期,菌體的自聚集率最高。這可能是因為細菌在增殖或衰亡時,會脫落外膜和釋放周質蛋白,衰亡期的細胞存在因細胞自融釋放的胞內蛋白,使得菌體外的聚合物較對數期和穩定期多[24]。Wang等[16]研究了Enterobactersp.strain FL在不同時間的自聚集性,發現在培養12 h時自聚集率大約為18%,但隨著時間的延長,自聚集性能逐漸增加,在培養48 h時,自聚集率能達到50%。由此可見,菌體的自聚集率與菌體的生長周期相關。
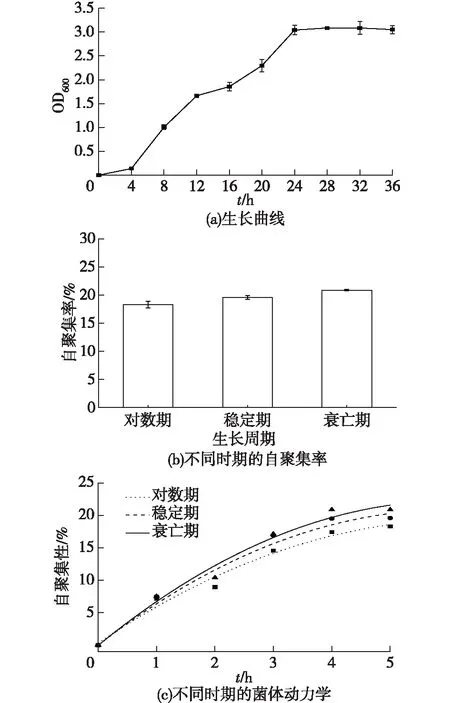
圖2 TN-10的生長曲線與自聚集性能Fig.2 Growth curve and auto-aggregationability of TN-10
菌體的自聚集動力學表明菌體克服靜電排斥后的自聚集能力。本實驗中,筆者對菌體的自聚集動力學進行研究,結果如圖2(c)所示。由圖2(c)可知:不同時期的菌體聚集率隨時間的增加而呈現緩慢上升的趨勢,但菌體的聚集性在5 h時還未達到完全平衡。同時,從0~5 h,衰亡期菌體的自聚集率始終比對數期和穩定期菌體的自聚集率高,其原因可能是處在不同時期的菌體胞外聚合物性質不同。不同條件下的菌體聚集動力學也相差較大。Zhang等[25]研究活性污泥的自聚集性發現,其自聚集率最大為50%左右,在3 h后保持穩定。
2.3 胞外聚合物組成
為研究菌體呈現出自聚集性能的原因,對菌體的胞外聚合物進行研究。為避免胞內蛋白溢出和細胞外膜脫落對菌體胞外聚合物成分測定的干擾,收集對數期的菌體對其胞外聚合物組成進行研究,結果如表1所示。由于胞外聚合物的主要成分為蛋白質、多糖等,測定蛋白質和多糖的含量來表征EPS[26]。由表1可知:蛋白質含量為每克干細胞中23.84 mg(相當于15.11 mg/L),多糖的含量為每克干細胞中18.64 mg(11.81 mg/L),胞外聚合物以蛋白質和多糖的總量計為每克干細胞中42.48 mg(26.81 mg/L)。TN-10的EPS中的蛋白質含量高于多糖含量,蛋白質和多糖含量的差異與菌體的生長特性和絮凝性相關。很多菌體EPS的多糖含量高于蛋白質含量,特別是在活性污泥的研究中,但有部分菌體的蛋白質含量高于多糖含量,如Acinetobactersp.YY-5[27]和Brevibacillussp.[26]等。王亮等[28]對1株白腐真菌(Phanerochaetechrysosporium)的EPS進行了研究,結果表明穩定期時的EPS為125 mg/L,其中蛋白質占30%,糖類占50%。Wang等[16]研究了1株異養反硝化菌(Enterobactersp.strain FL)的自聚集性能和胞外聚合物的關系,結果表明,該菌在12 h的自聚集率為17%左右,提取該菌的EPS發現,其蛋白質含量為每克干細胞中210 mg,多糖含量為每克干細胞中10 mg,其原因可能是該菌會產生較多的胞外酶。

表1 TN-10 EPS 中的蛋白質和多糖含量
注:以每克干細胞質量計。
2.4 胞外聚合物紅外光譜圖

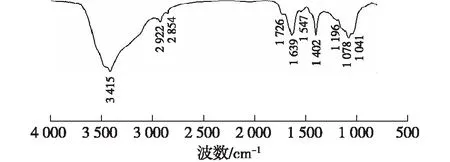
圖3 EPS的紅外特征光譜Fig.3 FT-IR spectra of EPS
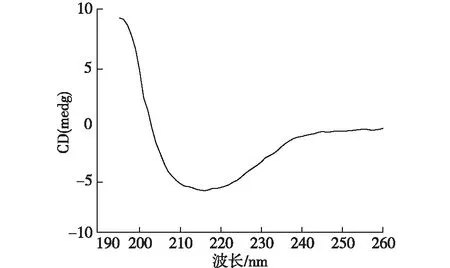
圖4 EPS的圓二色譜圖Fig.4 Circular dichroism spectra of EPS
2.5 圓二色譜測蛋白質二級結構
由于EPS中的蛋白質比多糖對絮凝性起著更重要的作用[16],而TN-10的EPS溶液中蛋白質含量高于多糖含量,使用圓二色譜對提取的胞外聚合物中蛋白質二級結構進行研究,結果如圖4所示。由圖4可知:在208和222 nm處的明顯雙峰是α-螺旋結構的特征峰,β-折疊在195和217 nm處有特征吸收峰,無規則卷曲在195 nm處有負的吸收峰[33]。利用CD pro程序包對圓二色譜的數據進行計算,其結果見表2。由表2可知:蛋白質中α-螺旋結構占主要比例,含量為55.08%。β-折疊和β-轉角所占的比例分別為7.31和13.72%。菌體的絮凝性和自聚集能力與β-折疊、α-螺旋和三螺旋結構結構所占的比例成正相關,但是會受到無規則卷曲和β-反向折疊的抑制[34]。在TN-10的EPS中對絮凝性起促進作用的α-螺旋和β-折疊所占比例之和為62.39%,高于對絮凝性起抑制作用的無規則卷曲所占比例23.89%,這也是TN-10具有良好絮凝性的原因。
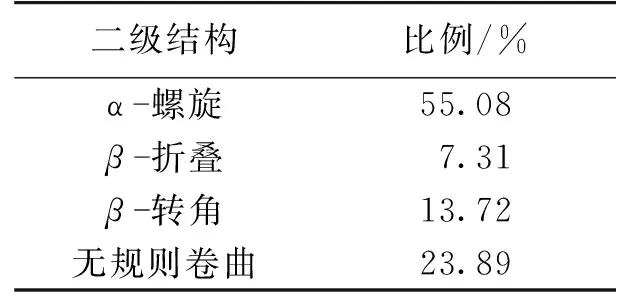
表2 EPS中蛋白質的二級結構
3 結論
筆者以1株硝化-反硝化菌Klebsiellasp.TN-10為研究對象,對其硝化-反硝化性能進行了考察,并對其絮凝性能進行了研究,主要研究結論如下。
2)研究了菌體的自聚集能力和聚集動力學,發現菌體在衰亡期的自聚集能力最強。
3)提取TN-10的胞外聚合物,測定其組成和含量,其中蛋白質含量為23.84 mg、多糖含量為18.64 mg(以每克干細胞質量計算)。
4)測定了EPS的紅外光譜,并對EPS溶液中的蛋白質二級結構進行測定,其組成為α-螺旋55.08%、β-折疊7.31%、β-轉角13.72%和無規則卷曲23.89%。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
