特定致腐菌對冷卻豬肉的致腐作用機制
2019-02-06 03:51:41戴金岳曾憲文方利敏姚明印劉木華黃林
肉類研究 2019年12期
戴金岳 曾憲文 方利敏 姚明印 劉木華 黃林


摘 要:為研究特定致腐菌在冷卻豬肉中的致腐作用機制,將韓國假單胞菌PS1(Pseudomonas koreensis PS1)和梭狀芽孢桿菌J4(Bacillus fusiformis J4)分別接種到新鮮冷卻豬肉中進行致腐實驗,以不接種致腐菌的豬肉為對照,于4 ℃條件下貯藏,每隔1 d測定各組肉樣的細菌總數、總揮發性鹽基氮(total volatile basic nitrogen,TVB-N)含量、蛋白質含量以及脂肪酶、彈性蛋白酶、胰蛋白酶和膠原蛋白酶活力。結果表明:各組肉樣細菌總數均呈現先上升后穩定的趨勢,貯藏第5天,接種P. koreensis PS1、B. fusiformis J4的肉樣細菌總數均超過8.0 (lg(CFU/g)),明顯大于對照組;接種特定致腐菌的2 組肉樣脂肪酶、彈性蛋白酶、胰蛋白酶和膠原蛋白酶活力均高于對照組,且呈現先上升后降低趨勢,至貯藏第7天,彈性蛋白酶、胰蛋白酶和膠原蛋白酶活力均達到峰值,而脂肪酶活力在貯藏第5天達到峰值;同時,各組肉樣蛋白質含量不斷降低,產物TVB-N含量明顯增加,貯藏第5天,接種特定致腐菌的2 組肉樣TVB-N含量均超出15 mg/100 g,明顯高于對照組。
關鍵詞:冷卻豬肉;特定致腐菌;酶活力;致腐機制
Abstract: Chilled fresh pork was inoculated with Pseudomonas koreensis PS1, Bacillus fusiformis J4 or nothing and stored at 4 ℃. Then, total viable count (TVC), total volatile basic nitrogen (TVB-N) content, protein content, and the activities of lipase, elastase, trypsin and collagenase in pork samples were determined. By doing so, we sought to explain the mechanism by which specific spoilage organisms (SSO) caused spoilage in chilled pork. The results showed that the TVC in all groups increased with storage time until reaching a plateau. The TVC in the two experimental groups exceeded 8.0 (lg (CFU/g)) on the fifth day, which was significantly higher than that of the control group. In the inoculated pork samples, the activities of the above four enzymes were higher than those in the control group, and showed a trend of increasing at first and then decreasing. The activities of elastase, trypsin and collagenase reached the peak on the seventh day, while the peak of lipase activity appeared on the fifth day. At the same time, the protein content decreased continuously in all groups, whereas the TVB-N content increased obviously. On the fifth day, the TVB-N content in the two experimental groups exceeded?15 mg/100 g, which was significantly higher than that of the control group.
Keywords: chilled pork; specific spoilage bacteria; enzyme activity; spoilage mechanism
DOI:10.7506/rlyj1001-8123-20190926-231
中圖分類號:TS251.51? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 文獻標志碼:A 文章編號:1001-8123(2019)12-0050-06
中國是世界豬肉生產和消費第一大國,多年來,豬肉產量和消費量一直位居世界第一[1],豬肉也是我國居民最主要的肉食品來源,因此保證豬肉品質安全至關重要。豬肉品質安全評價指標包括顏色、紋理、pH值、營養成分含量、嫩度和新鮮度等。新鮮度是肉品腐敗變質程度的直接反映,是評價肉品品質安全的最重要參數[2]。
變質豬肉不但營養價值下降甚至具有毒性,危及消費者健康。總揮發性鹽基氮(total volatile basic nitrogen,TVB-N)是蛋白質分解生成氨及胺類等含氮物質的總和,其含量會隨著肉品腐敗進程逐漸增加,與肉品腐敗程度呈正比。目前,TVB-N含量是我國國標評價肉品新鮮度的最重要指標[3]。
冷卻豬肉因能最大限度地保證口感與品質安全,已受到越來越多消費者的青睞[4]。但冷卻豬肉在貯藏過程中仍會腐敗變質,這是由于動物屠宰及加工過程中易被一些嗜冷致腐菌污染,肉的變質程度主要取決于致腐菌的種類和數量[5]。任靜等[6]通過對調理預制烤豬肉冷藏過程中微生物生長特性的研究發現,托盤包裝和真空包裝2 種包裝方式豬肉中存在的腐敗菌有假單胞菌﹑熱殺索絲菌、乳酸菌、節桿菌和氣單胞菌。Borch等[7]在豬肉、牛肉及其腌制肉制品中篩選鑒定出熱死環絲菌、假單胞菌屬﹑腸桿菌等腐敗微生物。Zhao Fan等[8]在對真空包裝冷卻豬肉貯藏過程中微生物的變化進行研究時發現,腐敗菌菌相隨貯藏時間的延長不斷發生變化,通過16S rDNA測得,貯藏第14天腸桿菌占所檢測到的各種腐敗微生物總序列的21.63%,第21天為6.50%。黎園園等[9]對解凍豬肉中優勢腐敗菌致腐能力的研究表明,假單胞菌和熱死環絲菌具有較強的致腐能力。
Skandamis等[10]研究表明,致腐菌群體感應現象在肉制品腐敗過程中起到重要作用。Zhu Suqin等[11]在4 ℃冷藏對蝦的腐敗過程中檢測到3 種自誘導劑,結合外源添加不同自誘導劑對冷藏對蝦腐敗影響的研究結果,說明群體感應影響冷藏對蝦的腐敗。目前,多數研究主要集中在肉品冷卻變質過程中的菌相變化、優勢致腐菌分類鑒定及群體感應等方面,而對特定致腐菌在冷卻肉中致腐作用機制的研究鮮見報道。
本課題組前期從變質冷卻豬肉中分離、篩選、鑒定出5 株致腐菌,通過TVB-N含量因子測定確定出2 株優勢致腐菌:韓國假單胞菌PS1(Pseudomonas koreensis PS1)和梭狀芽孢桿菌J4(Bacillus fusiformis J4)[12],并近一步利用原位熒光染色結合激光共聚焦掃描顯微鏡技術研究特定致腐菌P. koreensis PS1侵染豬肉的方式及其在豬肉中的分布情況,初步探索其對冷卻豬肉的致腐作用方式[13]。本研究將特定致腐菌P. koreensis PS1、B. fusiformis J4回接至冷卻豬肉,使其腐敗變質,測定該過程中細菌總數、蛋白質、TVB-N含量以及脂肪酶和蛋白酶活力的動態變化,更深入地探索特定致腐菌在冷卻豬肉中的致腐作用機制。
1 材料與方法
1.1 材料與試劑
冷卻里脊肉來自于不同的長白豬,購于江西省南昌市經濟技術開發區主要超市肉制品專柜。樣品置于密封袋內,放在加有冰袋的保溫箱中30 min內運回實驗室。
特定致腐菌P. koreensis PS1和B. fusiformis J4,課題組自行從變質冷卻豬肉中分離、鑒定、保藏[12]。
無水乙酸鈉﹑乙酸﹑氯化鈉﹑氫氧化鈉﹑鹽酸?西隴科學股份有限公司;氧化鎂﹑三氯乙酸 天津市大茂化學試劑廠;硫酸銅 天津市永大化學試劑有限公司;對硝基苯酚 上海展云化工有限公司;牛肉膏、蛋白胨(均為生化試劑) 北京奧博星生物技術有限責任公司;三羥甲基氨基甲烷(Tris)和福林酚 北京索萊寶(Solarbio)科技有限公司;地衣紅-彈性蛋白?美國Sigma公司;所用試劑均為分析純。
1.2 儀器與設備
722可見分光光度計、UV759紫外-可見分光光度計 上海佑科儀器儀表有限公司;JW-3022HR高速冷凍離心機 安徽嘉文儀器裝備有限公司;LDZX-50KBS立式壓力蒸汽滅菌鍋 上海申安醫療器械廠;LRH-250-C生化培養箱 廣東省韶關市泰宏醫療器械有限公司;HH恒溫水浴鍋 江蘇省金壇市中大儀器廠。
1.3 方法
1.3.1 菌懸液制備
將甘油管保藏的特定致腐菌P. koreensis PS1和B. fusiformis J4菌種在試管斜面培養基上活化,取長勢良好、無污染的菌體斜面,用無菌水制備成菌懸液,并利用染色顯微鏡檢計數法計數[14],即取5 μL菌懸液于載玻片1 cm2的區域內涂勻,用0.1 g/100 mL結晶紫染色,油鏡鏡檢并隨機選取10 個視野進行細菌計數,取平均值計算出菌懸液細胞含量,再用無菌水調整菌體初始濃度約為1×103 個/mL。
1.3.2 肉樣制備
在無菌操作臺上將肉樣表面切除約0.5 cm后,切成大小約為4 cm×4 cm×2 cm的樣本60 個,平均分成3 組,用無菌鑷子將每個樣品置于酒精燈上灼燒30 s,進行表面滅菌;取其中2 組分別在制備好的P. koreensis PS1和B. fusiformis J4菌懸液中浸潤10~15 s進行接種[9,15],取出瀝干放入無菌塑料袋中密封,剩余1 組不接種作為對照組;樣品全部置于4 ℃冰箱保存,分別于第1、3、5、7、9天的相同時段,3 組肉樣中各取4 個肉樣進行細菌總數、蛋白質、TVB-N含量以及脂肪酶、彈性蛋白酶、胰蛋白酶和膠原蛋白酶活力測定。
1.3.3 指標測定
1.3.3.1 肉樣細菌總數、蛋白質和TVB-N含量測定
采用GB/T 4789.2—2016《食品安全國家標準 食品微生物學檢驗 菌落總數測定》[16]中的平板菌落計數法測定細菌總數;采用GB/T 5009.5—2016《食品安全國家標準 食品中蛋白質的測定》[17]中的分光光度法測定蛋白質含量;采用GB 5009.228—2016《食品安全國家標準 食品中揮發性鹽基氮的測定》[18]中的半微量定氮法測定TVB-N含量。
1.3.3.2 肉樣酶活力測定
參考趙培培[19]的方法測定彈性蛋白酶活力;參考孫文斌[20]的方法測定脂肪酶和膠原蛋白酶活力;參考劉亮亮[21]的方法測定胰蛋白酶活力。4 種酶的酶活力單位均為U/g。
1.4 數據處理
采用Excel 2010軟件進行數據處理,利用Origin 2018軟件進行圖形繪制,DPS 7.05軟件進行顯著性分析(P<0.05為差異顯著);測定結果用平均值±標準差表示,實驗均重復3 次。
2 結果與分析
2.1 冷卻豬肉腐敗過程中細菌總數動態變化
優勢致腐菌生長繁殖是引起肉樣腐敗變質的主要原因,因此微生物數量是衡量肉樣腐敗程度的一個重要指標。由圖1可知,隨著貯藏時間的延長,對照組和接種特定致腐菌肉樣的細菌總數均呈增加趨勢,且在貯藏第5天進入穩定期,此時接種特定腐敗菌的肉樣細菌總數均超過8.0 (lg(CFU/g)),明顯高于對照組肉樣細菌總數(6.82 (lg(CFU/g)))。這說明接種至肉樣中的特定致腐菌P. koreensis PS1、B. fusiformis J4及肉樣自身所帶的微生物在4 ℃低溫下仍能利用肉樣中的營養物質快速生長繁殖,細菌總數明顯增加,但隨著營養物質的消耗和代謝產物的積累以及豬肉體系整個微環境pH值的改變,從而使得致腐菌的生長變得緩慢[22]。
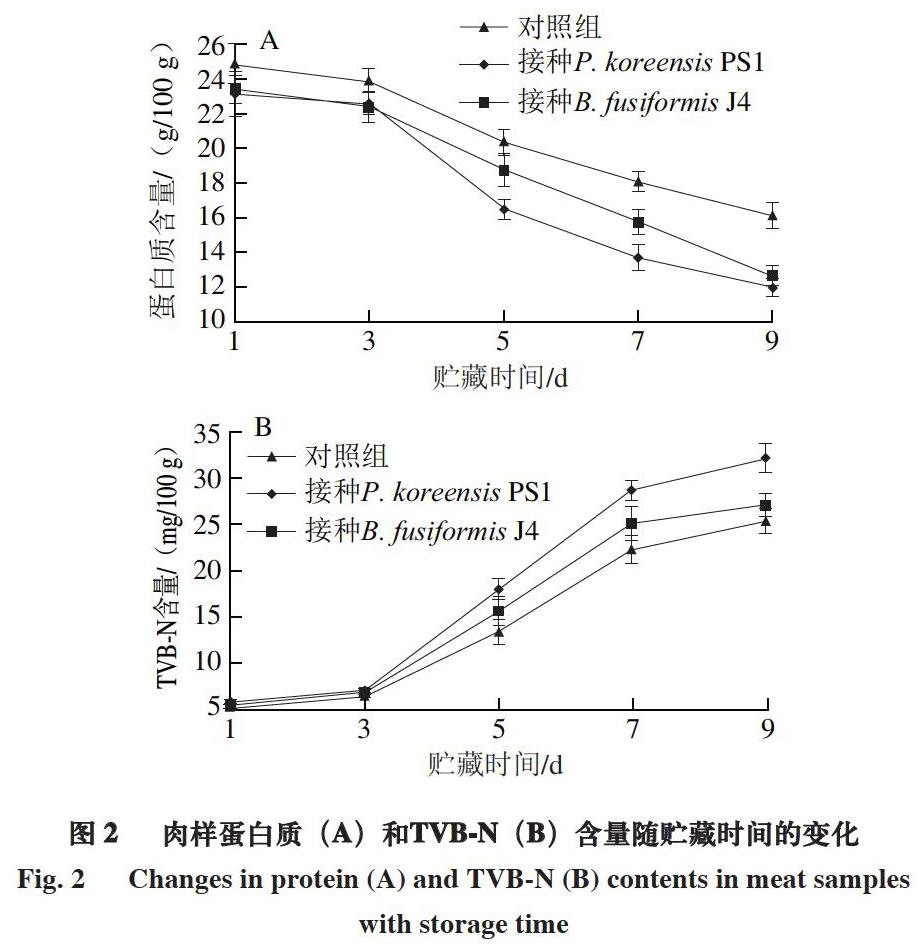
2.2 冷卻豬肉腐敗過程中蛋白質和TVB-N含量動態變化
蛋白質作為里脊肉中主要的營養物質會被微生物分解利用,同時產生氨氣、尸胺和腐胺等生物胺,TVB-N含量是評價肉品新鮮度最重要的指標,我國國標規定生鮮肉類TVB-N含量不可超過15 mg/100 g。
由圖2可知:各組肉樣中蛋白質和TVB-N含量在貯藏前3 d變化不明顯,這是由于腐敗菌首先利用肉樣中的糖原作為生長基質[23];隨著貯藏時間的延長,肉樣蛋白質含量明顯下降,TVB-N含量明顯增加,至貯藏第5天,接種特定致腐菌P. koreensis PS1、B. fusiformis J4肉樣的TVB-N含量均大于15 mg/100 g,明顯高于對照組。在冷卻豬肉變質過程中,接種特定致腐菌P. koreensis PS1、B. fusiformis J4的肉樣細菌總數增加、蛋白質被消耗和TVB-N含量的變化速率均大于對照組。同時,特定致腐菌P. koreensis PS1在肉樣中的生長趨勢與B. fusiformis J4類似,但其對蛋白質分解產生TVB-N的能力明顯優于B. fusiformis J4,這與本課題組之前的研究一致[12]。
2.3 冷卻豬肉腐敗過程中脂肪酶活力動態變化
豬肉脂肪中的亞油酸與油酸在氧氣和致腐菌所產脂肪酶的作用下先形成過氧化物,再被分解成醛類﹑酮類等小分子物質,致使豬肉的食用價值喪失[24]。
由圖3可知:在貯藏前3 d,接種特定致腐菌P. koreensis PS1、B. fusiformis J4和對照組肉樣脂肪酶活力快速上升;貯藏第5天,3 組肉樣的脂肪酶活力均達到峰值,分別為53.85、51.83、47.51 U/g,而后酶活力均出現下降趨勢,在此過程中接種P. koreensis PS1和B. fusiformis J4肉樣的脂肪酶活力保持較高水平,均高于對照組。可見,特定致腐菌P. koreensis PS1、B. fusiformis J4均可產生活性較強的脂肪酶,加快對肉樣肌纖維及黏膜中脂肪的破壞,更有利于致腐菌向肉樣深層侵染。
2.4 冷卻豬肉腐敗過程中胰蛋白酶﹑膠原蛋白酶及彈性蛋白酶活力動態變化
肉樣蛋白質的分解依靠胰蛋白酶﹑膠原蛋白酶和彈性蛋白酶等的作用來完成,膠原蛋白的破壞導致肉樣感官品質發生改變,同時更有利于腐敗微生物的侵染。
由圖4可知,肉樣腐敗過程中胰蛋白酶﹑膠原蛋白酶和彈性蛋白酶活力均出現相似的變化趨勢,接種致腐菌P. koreensis PS1﹑B. fusiformis J4及對照組肉樣的上述3 種蛋白酶活力均呈現先上升后下降的趨勢,至貯藏第7天,彈性蛋白酶﹑膠原蛋白酶﹑胰蛋白酶活力均達到峰值,且接種P. koreensis PS1﹑B. fusiformis J4肉樣的蛋白酶活力均明顯高于對照組,此后3 種蛋白酶活力略有降低。Tsigarida等[25]發現,屠宰后豬肉本身存在內源酶的糖酵解作用,而該作用在pH 5.4~5.5時會被終止,此后內源酶對豬肉的影響相較于腐敗菌的酶解作用可被忽略。腐敗菌先以糖類﹑脂質為底物進行分解,因而貯藏前3 d 3 種蛋白酶活力無明顯變化,之后胰蛋白酶、膠原蛋白酶和彈性蛋白酶活力均快速增加,蛋白質的分解也加快(圖2A),這可能是由于蛋白質氧化降解受活性氧的影響,而不飽和脂肪酸及脂肪氧化產物丙二醛對活性氧的生成具有促進作用[26-27]。
3 討 論
冷卻豬肉在加工和運輸過程中易被一些致腐菌侵染,致使豬肉品質下降,因此為更好地保鮮冷卻豬肉,需對致腐菌在豬肉中的致腐過程作進一步探討。顧春濤等[28]研究冷鮮牛肉微生物菌群變化時發現,假單胞菌和腸桿菌均屬于優勢致腐菌,其中假單胞菌致腐能力較強。同時Duan Shan等[29]研究發現,假單胞菌在4 ℃條件下冷藏的羅非魚中屬于優勢腐敗菌,且細菌總量與貯藏時間呈正比,將分離篩選得到的熒光假單胞菌和梭狀芽孢桿菌分別接種于冷卻豬肉,發現2 種腐敗菌在冷卻豬肉中快速生長,且細菌總數不斷增加,這與Li Ning等[30]對冷鮮豬肉貯藏過程中細菌群落組成研究中得出的隨著肉樣貯藏時間的延長致腐菌數量不斷上升的結果相一致。
豬肉富含蛋白質、脂肪等物質,其腐敗的原因主要是由于這些物質被致腐菌產生的蛋白酶及脂肪酶所降解,因而酶解反應是影響冷卻豬肉腐敗的重要因素[31]。Arslan等[32]發現,假單胞菌能產生具有較高活性的蛋白酶。Zhang Qiuqin等[33]將熒光假單胞菌回接至雞肉湯,進行致腐實驗,最終檢測到較強的蛋白酶和脂肪酶活性。肉制品腐敗過程中的脂質和蛋白質氧化被認為是相互影響的。Li等[27]在對脂質氧化分解產物丙二醛對兔肌球蛋白結構的影響進行研究時發現,丙二醛能促進蛋白質的氧化分解。本研究通過測定接種特定致腐菌冷卻豬肉腐敗過程中多種酶的活力變化表明,肉樣腐敗在致腐菌所產脂肪酶和蛋白酶的共同作用下完成。特定致腐菌產生的脂肪酶能夠對肉樣肌纖維膜及黏膜中的脂肪進行氧化分解,破壞肌肉結構組織,促進致腐菌的侵染繁殖,但隨著酶的作用和活力的增強,脂肪氧化分解產物醛、酮、酸等積累,會反饋阻遏和抑制相關脂肪酶的作用,并且貯藏5 d后出現細菌總數沒有明顯下降而脂肪酶活力快速下降的現象。同時,隨著致腐菌對肉樣深層的侵染繁殖,其彈性蛋白酶﹑膠原蛋白酶和胰蛋白酶等多種蛋白酶的活力有所增加,在貯藏第7天達到最大值,相應地,TVB-N含量也在此時增加最快,之后由于生物胺等代謝產物的積累,導致蛋白酶活力呈現下降趨勢。這與Zhang Qiuqin等[33]在將熒光假單胞菌接種至雞肉湯進行的致腐實驗中得到的蛋白酶與脂肪酶活力變化趨勢相類似。
致腐菌在生長初期能夠優先利用糖類﹑脂質等物質,而后產生蛋白酶以降解豬肉蛋白質,供其生長,這也是TVB-N含量在貯藏初期變化不明顯的原因。此后隨著貯藏時間的延長,肉樣蛋白質不斷被分解,TVB-N則會大量累積,產生該現象的直接原因可能與致腐菌的生長繁殖有關,這也與黎園園等[9]研究豬肉優勢腐敗菌產TVB-N的情況相近。因此降低致腐菌酶體系的活力,能夠很大程度上抑制腐敗的產生,這也為冷卻豬肉的保鮮提供了相關的理論依據。
4 結 論
特定致腐菌P. koreensis PS1﹑B. fusiformis J4在新鮮冷卻豬肉中仍能快速生長,導致細菌總數呈現先上升后穩定的趨勢,特定致腐菌優先產生脂肪酶,導致肌肉肌纖維及黏膜中脂肪氧化降解,并在酶活力較強的彈性蛋白酶﹑膠原蛋白酶和胰蛋白酶等多種蛋白酶作用下,促使肉樣中蛋白質分解,并增加TVB-N的釋放,從而加速冷卻豬肉的腐敗變質。
參考文獻:
[1] 梁澤閩, 趙文豪, 陶紅軍. 我國豬肉產業國際競爭力研究[J]. 中國豬業,?2019, 14(5): 21-28. DOI:10.16174/j.cnki.115435.
[2] LI Yongyu, ZHANG Leilei, PENG Yankun, et al. Hyperspectral imaging technique for determination of pork freshness attributes[C]//Sensing for Agriculture and Food Quality and Safety III. International Society for Optics and Photonics, Orlando, Florida, United States, 2011: 80270H. DOI:10.1117/12.883652.
[3] 中華人民共和國國家衛生和計劃生育委員會, 國家食品藥品監督管理總局. 食品安全國家標準 鮮(凍)畜、禽產品: GB 2707—2016[S]. 北京: 中國標準出版社, 2016.
[4] 顧海寧, 李強, 李文釗, 等. 冷卻豬肉貯存中的品質變化及貨架期預測[J]. 現代食品科技, 2013, 29(11): 2621-2626. DOI:10.13982/j.mfst.1673-9078.2013.11.023.
[5] BOUJU-ALBER A, PILET M F, GUILLOU S. Influence of lactate and acetate removal on the microbiota of French fresh pork sausages[J]. Food Microbiology, 2018, 76: 328-336. DOI:10.1016/j.fm.2018.06.011.
[6] 任靜, 牛海力, 張歡, 等. 調理預制烤豬肉冷藏過程中微生物生長特性的研究[J]. 中國食品學報, 2017, 17(7): 230-238. DOI:10.16429/j.1009-7848.2017.07.029.
[7] BORCH E, KANTMUERMANS M L, BLIXT Y. Bacterial spoilage of meat and cured meat products[J]. International Journal of Food Microbiology, 1996, 33(1): 103-120. DOI:10.1016/0168-1605(96)01135-X.
[8] ZHAO Fan, ZHOU Guanghong, YE Keping, et al. Microbial changes in vacuum-packed chilled pork during storage[J]. Meat Science, 2015, 100: 145-149. DOI:10.1016/j.meatsci.2014.10.004.
[9] 黎園園, 董慶利, 梁娜, 等. 解凍豬肉中優勢腐敗菌致腐能力研究[J]. 食品科學, 2011, 32(5): 123-127.
[10] SKANDAMIS P N, NYCHAS G J E. Quorum sensing in the context of food microbiology[J]. Applied and Environmental Microbiology, 2012, 78(16): 5473-5482. DOI:10.1128/AEM.00468-12.
[11] ZHU Suqin, WU Haohao, ZENG Mingyong, et al. The involvement of bacterial quorum sensing in the spoilage of refrigerated Litopenaeus vannamei[J]. International Journal of Food Microbiology, 2015, 192: 26-33. DOI:10.1016/j.ijfoodmicro.2014.09.029.
[12] 黃林, 陳全勝, 張燕華, 等. 冷卻豬肉優勢腐敗菌分離鑒定及致腐能力測定[J]. 食品科學, 2013, 34(1): 205-209.
[13] 黃林, 趙杰文, 陳全勝, 等. 冷卻豬肉優勢腐敗菌原位熒光染色檢測方法研究[J]. 中國食品學報, 2014, 14(3): 145-150. DOI:10.16429/j.1009-7848.2014.03.004.
[14] 沈萍, 陳向東. 微生物學實驗[M]. 4版. 北京: 高等教育出版社,?2007: 218.
[15] 彭勇. 冷卻豬肉常見腐敗微生物致腐能力的研究[D]. 北京: 中國農業大學, 2005: 18-19.
[16] 中華人民共和國國家衛生和計劃生育委員會, 國家食品藥品監督管理總局. 食品安全國家標準 食品微生物學檢驗 菌落總數測定: GB 4789.2—2016[S]. 北京: 中國標準出版社, 2016.
[17] 中華人民共和國國家衛生和計劃生育委員會, 國家食品藥品監督管理總局. 食品安全國家標準 食品中蛋白質的測定:?GB/T 5009.5—2016[S]. 北京: 中國標準出版社, 2016.
[18] 中華人民共和國國家衛生和計劃生育委員會. 食品安全國家標準 食品中揮發性鹽基氮的測定: GB 5009.228—2016[S]. 北京: 中國標準出版社, 2016.
[19] 趙培培. 產新型彈性蛋白酶產吲哚金黃桿菌的分離鑒定及酶學性質研究[D]. 雅安: 四川農業大學, 2012: 13-14.
[20] 孫文斌. 海洋微生物源膠原蛋白酶等酶系的研究[D]. 大連: 大連工業大學, 2014: 14-17.
[21] 劉亮亮. 羊胰酶提取工藝及水解酪蛋白動力學研究[D]. 蘭州: 甘肅農業大學, 2010: 19.
[22] AKKERMANS S, VAN IMPE J F. Mechanistic modelling of the inhibitory effect of pH on microbial growth[J]. Food Microbiology, 2018, 72: 214-219. DOI:10.1016/j.fm.2017.12.007.
[23] 黃敏欣, 趙文紅, 白衛東, 等. 冷卻豬肉腐敗菌的研究進展[J]. 肉類工業, 2015(5): 38-42; 46.
[24] CARLSEN C U, MOLLER J K S, SKIBSTED L H. Heme-iron in lipid oxidation [J]. Coordination Chemistry Reviews, 2005, 249(3/4):?485-498. DOI:10.1016/j.ccr.2004.08.028.
[25] TSIGARIDA E, NYCHAS G J E. Ecophysiological attributes of a Lactobacillus sp. and a Pseudomonas sp. on sterile beef fillets in relation to storage temperature and film permeability[J]. Journal of Applied Microbiology, 2001, 90(5): 696-705. DOI:10.1046/j.1365-2672.2001.01292.x.
[26] LUND M N, HEINONEN M, BARON C P, et al. Protein oxidation in muscle foods: a review[J]. Molecular Nutrition and Food Research, 2011, 55(1): 83-95. DOI:10.1002/mnfr.201000453.
[27] LI S J, KING A J. Structural changes of rabbit myosin subfragment 1?altered by malonaldehyde, a byproduct of lipid oxidation[J]. Journal of Agricultural and Food Chemistry, 1999, 47(8): 3124-3129. DOI:10.1021/jf990028y.
[28] 顧春濤, 畢偉偉, 朱軍莉. 冷鮮牛肉貯藏中菌群結構及優勢菌致腐性的分析[J]. 食品科學, 2019, 40(18): 76-82. DOI:10.7506/spkx1002-6630-20180925-263.
[29] DUAN Shan, ZHOU Xingzhi, MIAO Jianyin, et al. Succession of bacterial microbiota in tilapia fillets at 4 ℃ and in situ investigation of spoilers[J]. World Journal of Microbiology and Biotechnology, 2018, 34(5): 69. DOI:10.1007/s11274-018-2452-5.
[30] LI Ning, ZHANG Youxiong, WU Qingping, et al. High-throughput sequencing analysis of bacterial community composition and quality characteristics in refrigerated pork during storage[J]. Food Microbiology, 2019, 83: 86-94. DOI:10.1016/j.fm.2019.04.013.
[31] 趙麗珺, 謝晶, 馮淑嬌, 等. 冷卻豬肉中產蛋白酶腐敗菌的分離鑒定[J]. 食品工業科技, 2014, 35(7): 134-139; 143. DOI:10.13386/j.issn1002-0306.2014.07.041.
[32] ARSLAN S, EYI A, ?ZDEMIR F. Spoilage potentials and antimicrobial resistance of Pseudomonas spp. isolated from cheeses[J]. Journal of Dairy Science, 2011, 94(12): 5851-5856. DOI:10.3168/jds.2011-4676.
[33] ZHANG Qiuqin, YE Keping, WANG Huhu, et al. Inhibition of biofilm formation of Pseudomonas aeruginosa by an acylated homoserine lactones-containing culture extract[J]. LWT-Food Science and Technology, 2014, 57(1): 230-235. DOI:10.1016/j.lwt.2013.12.022.
