相同攝氧量下向心運動和離心運動對脂代謝肌肉因子激活作用的對比研究
2019-01-18 03:22:02劉陽何玉秀梅家順
中國運動醫學雜志 2018年12期
關鍵詞:研究
劉陽 何玉秀 梅家順
1 河北師范大學體育學院人體運動生物信息測評省重點實驗室(石家莊 050021)
2 河北師范大學體育學博士后科研流動站
肥胖已經成為日益嚴重的社會問題,而運動是目前公認的安全、有效的減肥方法之一,然而何種運動方式或運動強度的減肥效果最佳目前仍不能確定[1,2]。運動減肥的最主要機制是骨骼肌收縮引起的能量代謝改變和脂肪分解增強。近年來研究發現,骨骼肌可作為一個內分泌器官向血液中釋放調節物質,Pedersen等在2003年首次將這些物質命名為“肌肉因子(Myokine)”[3]。在常見的肌肉因子中,鳶尾素(Irisin)和肌聯素(C1q tumornecrosis factor-related rotein 15,CTRP15)與脂代謝密切相關。Irisin是肌細胞內合成纖維連結蛋白Ⅲ型域包含蛋白5(fibronectin typeⅢ domain-containing protein 5,FNDC5),經蛋白酶剪切分泌入血的肌肉因子,可通過誘導解耦聯蛋白1(UCP-1)高表達,促進脂肪氧化分解。CTRP15同樣由骨骼肌分泌,可以促進游離脂肪酸向脂肪細胞內的轉運。這兩種脂代謝肌肉因子在運動減肥過程中扮演著重要作用[4,5]。
研究證明[6],不同的運動方案,即使能量消耗水平相當,其減肥效果也不相同。這可能與不同運動方案對肌細胞的刺激不同有關。我們之前的研究顯示[7],離心收縮對骨骼肌的刺激更為明顯,相對于向心收縮可以造成更顯著的肌細胞胞漿“鈣超載”現象,而細胞內鈣離子濃度的升高對CTRP15和Irisin的上游調控蛋白過氧化物酶體增殖物激活受體γ輔激活因子1α(PPARγ coactivator 1-alpha,PGC-1α)均有激活作用。因此離心運動相比于向心運動,可能會更強烈地刺激肌肉分泌脂代謝相關的肌肉因子。Trevor的研究顯示,能量消耗更低的下樓(離心收縮)運動與能量消耗更高的上樓(向心)運動的減體重效果相同,但離心運動對血脂、血壓和安靜心率的改善效果較向心運動更好[8]。這種現象可能與離心收縮刺激骨骼肌分泌更多脂代謝肌肉因子有關。
本研究給予大鼠一次相同攝氧量的向心和離心運動干預,對比觀察兩種運動后0~48小時內骨骼肌FNDC5、血清Irisin和Ctrp15的變化,初步從分子層面探討減肥運動處方中,肌肉的離心收縮形式是否能夠有效地刺激脂代謝肌肉因子的分泌,從而更顯著地改善機體脂代謝。
1 對象與方法
1.1 實驗動物與分組
潔凈級8周齡雄性SD大鼠78只,體重200.0±19.8 g,購于北京維通利華動物公司。自由攝食飲水,溫度控制在24℃±3℃、濕度40%~60%。隨機分為攝氧量測試組(OE組和OL組,n=6)、安靜對照組(C組,n=6)、向心運動組(E組,n=30)與離心運動組(L組,n=30)。參考相關研究,由于離心訓練會使骨骼肌在運動后0~6小時與24~48小時出現顯著的結構改變與細胞胞漿鈣超載現象[7],而鈣離子濃度又正是Irisin與CTRP15的上游信號,故對運動干預組在運動后0、6、12、24、48小時分別進行取材。具體分組見表1。

分組 亞組 干預方式 運動后取材時間點向心運動組(E組)E0 E6 E12 E24 E48向心運動即刻6小時12小時24小時48小時離心運動組(L組)L0 L6 L12 L24 L48離心運動即刻6小時12小時24小時48小時
1.2 運動方案
首先對OE和OL組在動物氣體分析儀(Oxymax Equal Flow,Columbus,美國)內的跑臺上進行特定跑速下的攝氧量測試。3天適應性訓練后進行正式測試。根據H?ydal的動物模型[9],OE組采用約70%VO2max強度跑臺訓練,跑速20 m/min,坡度為0°。OL組用相同的跑速,相比常用的-10°或-15°下坡跑方案,使用較小的下坡跑坡度(-5°)進行訓練時,OL組攝氧量與OE組較為接近。也有研究者使用接近-5°的坡度(相對坡度8.5%~10%)成功進行了離心運動訓練[10,11],故本研究最終選用-5°作為下坡跑運動干預的坡度。
通過觀察訓練時攝氧量曲線穩定至出現攝氧量平臺,取平臺期攝氧量平均數為該鼠在此訓練方案下的攝氧量,如測試時間超過10 min仍未出現平臺則實驗終止。測試所得OE組與OL組攝氧量數據見表2,兩組攝氧量獨立樣本t檢驗P=0.94,無顯著性差異,說明大鼠在20 m/min的跑臺運動中,采用0°平坡跑運動與-5°下坡跑運動攝氧量相同。

表2 向心與離心運動平臺期攝氧量
在OE與OL組確定運動攝氧量后,對E組與L組進行一次與OE和OL組相同跑速的跑臺運動。由于本研究采用一次運動的干預方法,參考相關研究[7,9],為避免大鼠對未知環境產生應激,首先進行3天低強度適應性運動,跑速5 m/min,時間10 min,之后進行正式運動干預,E和L組跑速均為20 m/min,時間60 min,E組跑臺坡度0°,L組為-5°。如有大鼠無法完成1小時的訓練而提前達到力竭,則將其從實驗對象中剔除。
1.3 取材
E組與L組進行一次運動后,分別在0、6、12、24、48小時取材。C組在E和L組取材結束后3小時內完成取材。大鼠腹腔注射5%戊巴比妥鈉(50 mg/kg),右心耳處取血,4℃靜置20 min,3500轉/min離心15 min后,抽取血清-80℃凍存。取血后剝離腿部肌肉,取右側腓腸肌肌腹部分液氮保存以備蛋白表達測試。E0、L0、E12、L12組左側腓腸肌固定后進行電鏡觀察(方法見1.4)。
1.4 骨骼肌微細結構觀察
根據以往研究[7],離心運動后12~24小時肌細胞微細損傷達到峰值。為驗證本研究-5°下坡跑訓練方案能夠達到離心運動對肌細胞造成的微細損傷效果,采用透射電鏡觀察運動后即刻與12小時腓腸肌微細結構變化。取材獲得的肌肉組織縱切成為3 mm×2 mm小塊,4%戊二醛固定,1/15 m磷酸緩沖液洗3次,1%鋨酸固定,1/15 m磷酸緩浸洗2次,丙酮梯度脫水(50%、70%、80%、90%、100%),丙酮滲透3小時,包埋劑滲透5小時,37℃聚合12小時,超薄切片,醋酸雙氧鈾和酸鉛雙重染色。隨機選取視野,觀察Z線和線粒體形態。
1.5 骨骼肌PGC-1α、FNDC5蛋白表達
使用蛋白質免疫印跡法(Western blot)測試大鼠腓腸肌PGC-1α、FNDC5蛋白表達量。肌肉勻漿并裂解后,BCA法測定蛋白濃度。加入5倍Loading Buffer,煮沸。SDS-PAGE凝膠體系,濃縮膠4%,分離膠10%~12%。80 V,50 mA電泳至樣品進入分離膠,120 V電泳直至結束。PVDF膜濕轉,恒流300 mA,90 min。脫脂奶粉封閉2小時,一抗4℃孵育過夜(PGC-1α、FNDC5一抗為兔抗鼠抗體,稀釋比例均為1∶2000,購自Abcam)。TBST洗四次后二抗(羊抗兔抗體,稀釋比例1∶10000,購自中杉金橋)孵育2小時,再洗5次后發光。凝膠成像系統分析蛋白質印跡灰度,使用GAPDH作為內參計算蛋白質表達。
1.6 血清Irisin、CTRP15含量
血清Irisin和CTRP15濃度采用酶聯免疫法(ELISA)測試,嚴格依照試劑盒步驟操作(Irisin試劑盒購自Phoenix Pharmaceuticals,CTRP15試劑盒購自 Nordic BioSite)。
1.7 統計學方法
數據使用平均數±標準差(±s)表示。蛋白表達的計算以對照組數據作為干預前基礎值(baseline),取其表達量為1,并將所有組條帶灰度值除以對照組灰度值,得到相對蛋白表達量。使用雙因素方差分析(Twoway ANOVA)進行數據統計,以運動方式(向心和離心)和取材時間(對照組作為運動前、運動后0~48小時)為固定因子。如兩因子存在顯著交互作用,則以簡單效應(simple effect)分別分析兩種運動前后不同時間點的變化差異;如不存在顯著交互作用,則以主效應(main effect)來反映兩種運動的效應差異;多重比較采用最小顯著性差異法(least significant difference,LSD)兩兩比較各時間點間和各運動組間差異。統計軟件為SPSS24.0,取P<0.05為顯著性差異,P<0.01為非常顯著性差異。
2 結果
2.1 離心與向心運動對骨骼肌微細結構的影響
如圖1所示,在一次運動結束后即刻,向心運動組骨骼肌細胞Z線排列較為整齊,離心運動組則出現了Z線扭曲的現象。在運動結束后12小時,向心運動組出現了少量的Z線扭曲,而離心運動組則出現了大量的Z線扭曲和明顯的線粒體膨大現象,顯示坡度縮小至-5°的下坡跑依然能夠造成相當程度的骨骼肌微細結構改變。
2.2 離心和向心運動對骨骼肌PGC-1α和FNDC5蛋白表達的影響
如圖2、表3所示,雙因素方差分析顯示,取材時間(0~48小時)和運動方案(向心、離心)兩因素無顯著交互效應(P>0.05),不同運動方案間存在非常顯著的主效應(P<0.01),說明離心運動比向心運動對PGC-1α表達作用更明顯。不同時間點之間也存在顯著主效應(P<0.01),多重比較顯示,運動后即刻、6小時、24小時和48小時,兩組PGC-1α蛋白表達量均顯著高于對照組(P<0.05或P<0.01),12小時與安靜對照無顯著差異。說明離心和向心訓練后,PGC-1α除12小時回落至基線值外,在48小時內一直維持著高表達。
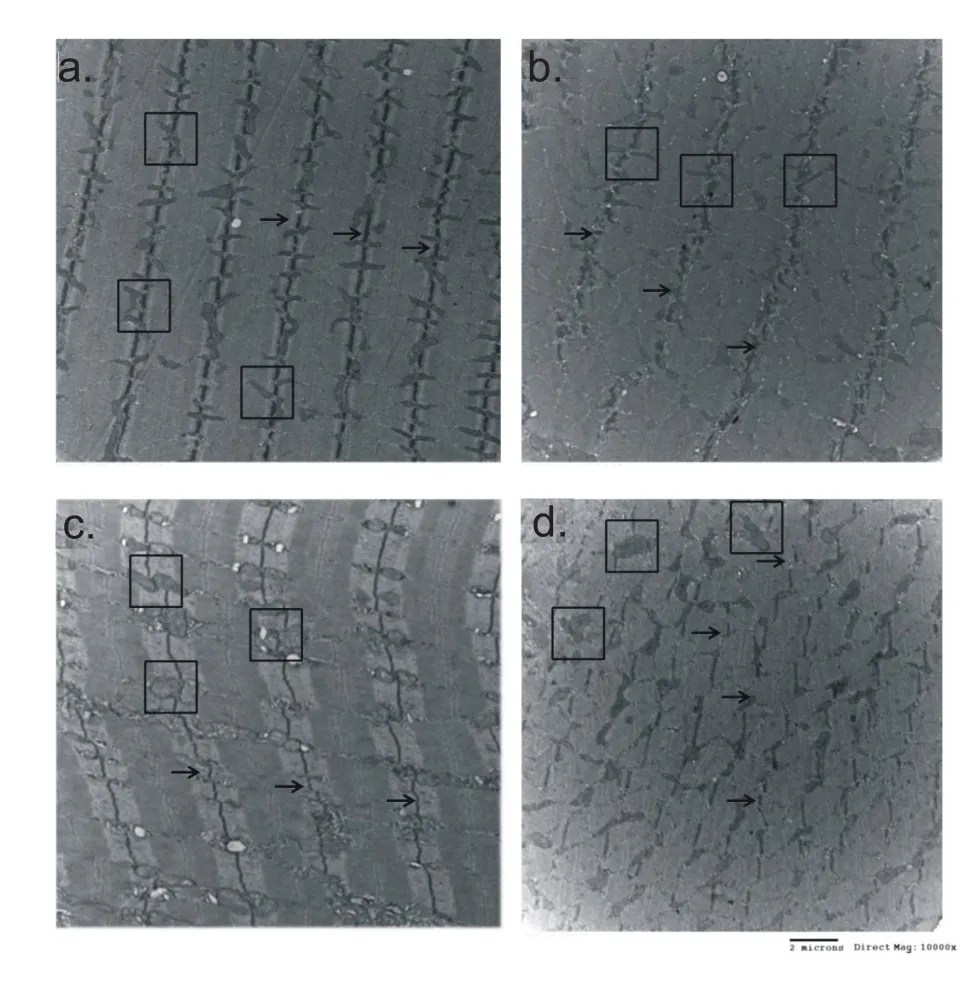
圖1 運動后即刻與12小時骨骼肌微細結構變化
雙因素方差分析結果顯示,取材時間(0~48小時)和運動方案(向心、離心)兩因素無顯著交互效應(P>0.05),運動方案存在顯著的主效應(P<0.05),取材時間存在非常顯著的主效應(P<0.01)。說明離心運動相比向心運動能更高地激活FNDC5表達,且運動后不同時間點FNDC5的表達具有顯著性變化。多重比較結果顯示,在運動后6小時和24小時,FNDC5蛋白表達量相比對照組基線值有非常顯著性升高(P<0.01),其他時間點無顯著差異。這說明離心和向心運動后,FNDC5出現6小時和24小時兩次高峰表達。
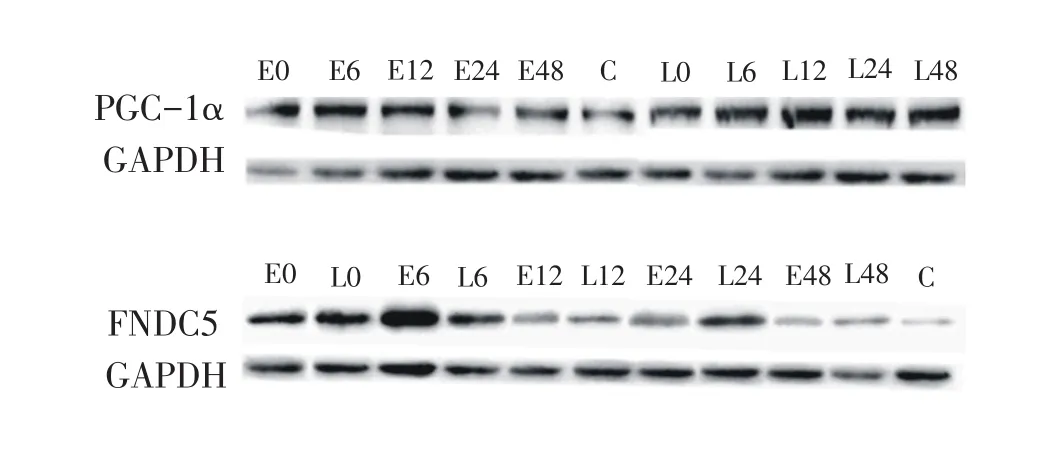
圖2 訓練后48小時內PGC-1α與FNDC5蛋白印跡
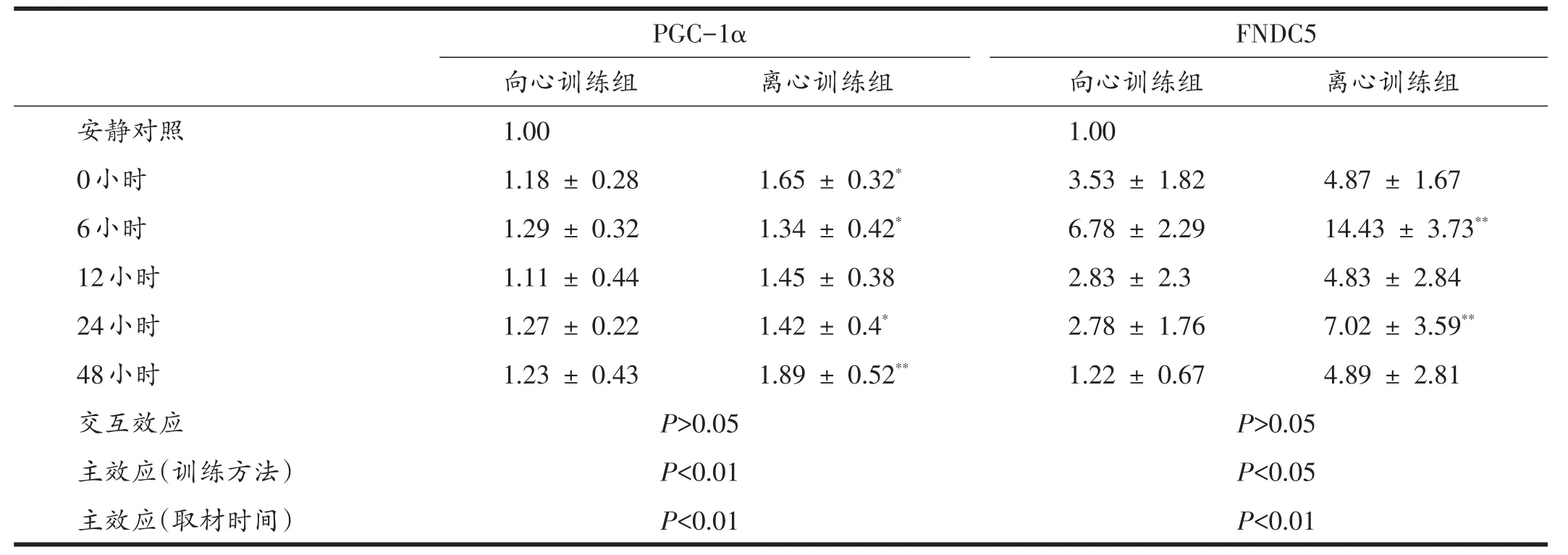
表3 訓練后48小時內PGC-1α與FNDC5蛋白表達
2.3 離心和向心運動對血清Irisin濃度的影響
血清Irisin濃度表達如表4所示。雙因素方差分析顯示,取材時間(0~48小時)和運動方案(向心、離心)兩因素存在非常顯著的交互效應(P<0.01),提示不同的運動組,Irisin在不同的時間點變化規律不同。
以取材時間點為單因素進行簡單效應分析顯示,相比于對照組:向心運動后48小時內,只有6小時有非常顯著的升高(P<0.01),其余時間點變化無顯著性;而離心運動組在運動后12和48小時有非常顯著的升高(P<0.01),其余時間點無顯著性變化。
以不同訓練方法為單因素進行簡單效應分析顯示,運動后6小時離心運動組Irisin濃度顯著性低于向心運動組(P<0.05),運動后12和48小時離心運動組非常顯著性高于向心運動組(P<0.01)。
2.4 離心和向心運動對血清CTRP15濃度的影響
血清CTRP15濃度表達如表4所示。雙因素方差分析顯示,取材時間(0-48小時)和方案(向心、離心)因素存在非常顯著的交互效應(P<0.01),提示兩組數據變化規律不一致。
以取材時間點為單因素進行簡單效應分析顯示,相比于對照組,向心運動后48小時內,只有6小時有非常顯著的升高(P<0.01),其余時間點變化無顯著性;而離心運動組在運動后即刻、12和48小時有非常顯著的升高(P<0.01),其余時間點無顯著性變化。
以不同訓練方法為單因素進行簡單效應分析顯示,運動后0小時向心運動組CTRP15濃度顯著性低于離心運動組(P<0.05),12和 48小時后向心運動組CTRP15濃度非常顯著性高于離心訓練組(P<0.01),而在6小時向心運動組CTRP15濃度顯著性高于離心運動組(P<0.05)。
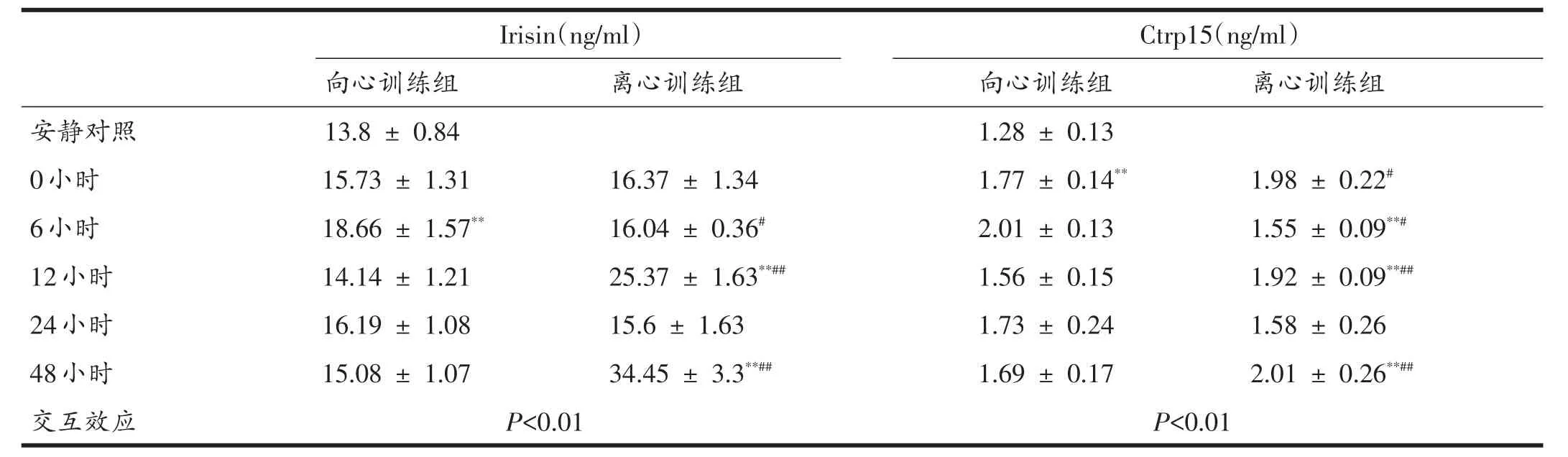
表4 訓練后48小時內血清Irisin與CTRP15濃度
3 討論
3.1 離心和向心運動對骨骼肌PGC-1α蛋白表達的影響
PGC-1α是細胞內重要的能量調節因子,普遍存在于線粒體豐富的組織如骨骼肌、肝臟、棕色脂肪內。它可作用于多種與能量代謝密切相關的轉錄因子及核受體,進而調節脂肪酸氧化、糖異生、組織產熱、線粒體合成等能量代謝過程。運動、寒冷、饑餓等多種應激狀態都可以激活PGC-1α信號,以促使機體改變能量代謝狀態進而適應新的環境[12]。運動可增加骨骼肌PGC-1α表達,進而改變肌細胞的能量代謝,但不同的運動方式對其作用不同。相對于向心收縮,離心收縮可更強烈地促使肌細胞質內Ca2+的釋放[13],而Ca2+濃度又是刺激PGC-1α表達的重要信號,PGC-1α的提高反過來又能夠抑制過高的Ca2+濃度[14],所以離心運動是否可以更強烈地激活PGC-1α值得進一步研究。
不同肌肉收縮形式對PGC-1α的影響研究相對較少,Conwright對絕經期女性進行共10組、每組10次最大力量的離心抗阻訓練后發現,PGC-1α蛋白表達并沒有顯著性升高[15]。宋超對小鼠分別進行上坡跑向心訓練(5°,10 m/min)和下坡跑離心訓練(-10°,13 m/min)后發現,離心訓練在運動結束后1小時可顯著提升PGC-1α表達,但升高程度沒有上坡跑提升幅度高[16]。離心收縮對骨骼肌是一個較強的刺激,而PGC-1α過表達可以有效地保護離心收縮導致的骨骼肌微損傷[17],該研究中離心訓練對PGC-1α提升的效果不明顯可能是由于訓練強度的不適宜所導致的。Conwright的研究采用的是最大力量的離心抗阻訓練,訓練強度很高,但抗阻訓練導致肌肉實際的運動時間很短,訓練量不足。Edgett對青年男性分別進行3組不同峰值強度(分別為最大攝氧量的73%、100%、133%)的高強度間歇訓練,通過肌肉活檢發現,雖然3種訓練方法都可提高PGC-1α的mRNA量,但峰值為133%最大攝氧量的訓練對PGC-1α的mRNA提升量顯著性低于73%、100%組,提示并非運動強度越高,對PGC-1α信號的激活越明顯[18]。
本研究采用中等強度下坡跑離心訓練(-5°,20 m/min)對SD大鼠進行干預,在我們前期的研究中,此模型已經被證實能夠較為有效地使肌細胞Ca2+濃度升高。大鼠PGC-1α蛋白表達在運動后0~6小時和24~48小時分別出現兩次峰值,與肌細胞離心訓練后鈣超載的變化規律基本相似[7],且電鏡照片結果也與離心運動造成的骨骼肌微細結構變化類似(實驗結果2.1),表明本研究采用的離心訓練方案可能是通過造成胞漿Ca2+濃度升高,進而激活PGC-1α信號。同時,離心訓練和向心訓練兩者存在非常顯著的主效應(P<0.01),且離心訓練的值較高,說明即使攝氧量不同,離心運動依然比向心運動更顯著地激活了PGC-1α的表達。
3.2 骨骼肌細胞內合成的FNDC5經剪切修飾后釋放入血的
運動對FNDC5與Irisin的影響是目前的研究熱點,Irisin是骨骼肌細胞內合成的FNDC5經剪切修飾后釋放入血的細胞因子,可通過調控UCP-1促進脂肪燃燒。細胞內的FNDC5受PGC-1α調控激活合成,骨骼肌是合成FNDC5并向血液分泌Irisin的最重要的組織[19],其次脂肪也具備類似的功能。
離心運動可較為明顯地通過Ca2+的釋放激活PGC-1α,那么是否可以對FNDC5與Irisin產生激活作用呢?此方面的研究較少。于濤等[20]研究發現一次-16°的下坡跑離心運動后,1~3天內FNDC5 mRNA較安靜時明顯升高,而血清Irisin含量只在運動后24小時升高,之后便回落。但該研究并未與向心運動進行對比。本實驗結果顯示,在20 m/min的跑速下,平坡跑與較小坡度下坡跑的攝氧量強度沒有顯著差異(見1.2訓練方案),造成FNDC5/Irisin的不同變化趨勢,可能主要是由不同肌肉收縮形式造成的。本研究發現,無論離心或向心運動,都可以顯著提升運動后FNDC5在6小時和24小時的表達,且離心訓練較向心訓練提升的更明顯(存在顯著的主效應)。這與PGC-1α的變化規律類似,說明相同攝氧量強度下離心運動較向心運動能更顯著地提升Ca2+—FNDC5信號。
值得注意的是,本研究中雖然離心運動組FNDC5顯著高于向心運動組,但離心組血清Irisin只在12和48小時顯著高于向心組,在6小時卻低于向心組。這可能是由于運動雖然可以通過PGC-1α提高FNDC5的合成量,但血清Irisin濃度不僅受骨骼肌FNDC5合成量的影響,同時受到血容量、機體代謝等諸多因素影響,從而表現出一種快速變化且與PGC-1α和FNDC5不同步的規律。大量研究表明,一次運動可以迅速提高血清Irisin濃度,而長期運動后Irisin濃度不變甚至降低[20-22],且FNDC5與Irisin的變化并不同步[23],這與本研究的中FNDC5與Irisin變化不同步的結果是相符的。以上這些結果說明,FNDC5相比于Irisin作為反映運動刺激骨骼肌分泌促脂肪氧化的細胞因子的指標,更為穩定可靠。本研究發現,雖然E組與L組在不同的時間點各有Irisin的升高,但離心運動組FNDC5表達在48小時內始終顯著高于向心組(主效應顯著),說明離心運動對PGC-1α—FNDC5的激活作用比向心運動更顯著,促Irisin分泌的效果更明顯。
3.3 離心和向心運動對CTRP15的影響
2012年Seldin等發現了參與調節脂代謝的肌肉因子CTRP15[5]。CTRP15主要在骨骼肌表達,釋放入血后可上調脂肪細胞與肝對血液游離脂肪酸的攝取。目前調控CTRP15的上游信號尚不完全清楚,運動、飲食、代謝與應激都可以影響到CTRP15的合成量。目前已知的是,肌細胞內CTRP15受cAMP和Ca2+濃度的調節[21]。運動對CTRP15的作用的研究結果也不完全一致,Seldin對大鼠進行2周的跑輪訓練后發現,無論是骨骼肌還是血液內的CTRP15都顯著升高,但Seldin的研究發現,在9周的跑輪訓練后,骨骼肌CTRP15的mRNA顯著降低[5]。造成這些研究結果不盡相同的原因可能與不同運動對Irisin的作用效果差異很大的機制類似:CTRP15和Irisin都是受Ca2+濃度或激素調節的肌肉因子,其運動后變化的時程可能較短。目前對CTRP15進行運動干預都是采用數周的長期干預,長期訓練后,機體對不同的訓練方式的適應程度不同,同時取材時間點的差異都可能造成結果的不同。所以對CTRP15進行一次運動后不同時間點的研究就較為必要。
本研究發現,與對照組相比,無論是離心還是向心運動,都可以顯著提高血清CTRP15濃度,但兩組在不同的時間點升高的幅度不同,造成這種現象的主要原因,可能是離心和向心運動激活了不同的CTRP15上游調控信號。在攝氧量相同的條件下,離心運動對胞漿Ca2+濃度的作用雖然更顯著,但向心運動可能通過cAMP等其他途徑提高了骨骼肌CTRP15的分泌量。但同時應該注意到,向心運動組CTRP15只有在運動后6小時顯著升高;離心運動組雖然在6和24小時回落到基線水平,但在0、12和48小時持續維持在高濃度,說明離心運動可以使肌肉在更長時間內維持CTRP15的高分泌水平,在更長的時間內持續調節機體脂肪轉運。
科學合理地制定減肥或改善脂代謝的運動處方,就必須考慮肌肉的收縮形式造成的影響。已有學者發現特定的離心運動可以達到更理想的提高有氧能力和改善脂代謝的作用[8,24,25]。本研究對離心運動促進骨骼肌脂代謝肌肉因子的分泌的相關結果,可為相關運動處方的制定提供理論依據。
4 結論
(1)相同攝氧量下,離心運動對PGC-1α、FNDC5的激活作用比向心運動更顯著。
(2)相同攝氧量下,離心和向心運動均可提高Irisin與CTRP15濃度,但離心運動可在運動后更長時間內促進兩肌肉因子的高分泌。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
