黃曲霉產(chǎn)胞外β-1,3-1,4-葡聚糖酶的發(fā)酵條件優(yōu)化
2019-01-17 01:41:18陳瑤瑤劉楊柳李思霆賈英民
食品科學(xué)技術(shù)學(xué)報 2019年1期
關(guān)鍵詞:實驗
劉 璐, 陳 洲, 陳瑤瑤, 劉楊柳, 李思霆, 賈英民
(北京工商大學(xué) 食品學(xué)院, 北京 100048)
β-1, 3-1, 4-葡聚糖是由β-葡萄糖苷通過β-1, 3和β-1, 4-糖苷鍵混合連接而成的線性多糖,廣泛分布于大麥、燕麥、高粱、大米和小麥等谷物的胚乳細(xì)胞壁中[1-2]。β-葡聚糖具有黏性高、水溶性差等特點,這給富含β-葡聚糖谷類作物的應(yīng)用造成了眾多不利影響[3-4]。例如,在啤酒釀造中,因β-葡聚糖的過量存在,使得麥芽汁黏度增加,造成過濾困難,降低了啤酒的產(chǎn)量和質(zhì)量[5];在飼料工業(yè)中,過高的β-葡聚糖會造成動物胃液黏度的增加,阻礙動物的營養(yǎng)吸收,從而降低飼料的轉(zhuǎn)化率,限制了谷物及其副產(chǎn)品在飼料添加中的應(yīng)用[6-7]。可見,尋求β-葡聚糖的“綠色”、高效降解方法將成為有效提升其工業(yè)化應(yīng)用的關(guān)鍵。一般來說,β-葡聚糖可通過酸解、堿解、酶解及其協(xié)同降解等多種方式轉(zhuǎn)化生成小分子的葡萄糖或纖維寡糖。酶解法因具有安全高效、專一性強及無毒副產(chǎn)物生成等優(yōu)勢而成為β-葡聚糖降解的最理想方法[8]。β-1, 3-1, 4-葡聚糖酶(EC 3.2.1.73)屬于糖苷水解酶,它能專一性水解β-葡聚糖主鏈中的β-1, 3/1, 4-葡萄糖苷鍵,是一類重要的工業(yè)化酶制劑,目前在啤酒釀造、飼料營養(yǎng)強化等領(lǐng)域中應(yīng)用廣泛[9]。
β-1, 3-1, 4-葡聚糖酶主要來源于動、植物及微生物,微生物因產(chǎn)酶量高、發(fā)酵周期短且易于實現(xiàn)工業(yè)化生產(chǎn)等優(yōu)勢而成為該酶的主要生產(chǎn)者。產(chǎn)β-1,3-1,4-葡聚糖酶的菌株種類繁多,其中,曲霉菌是最主要真菌來源。目前的研究主要集中于黑曲霉β-1, 3-1, 4-葡聚糖酶[10-11],有關(guān)其他曲霉來源的β-1, 3-1, 4-葡聚糖酶的報道還很少,因此,進(jìn)一步挖掘產(chǎn)β-1, 3-1, 4-葡聚糖酶的曲霉菌種對促進(jìn)該酶的生產(chǎn)和應(yīng)用具有重要指導(dǎo)意義。
通過初步探索,本課題組獲得了一株優(yōu)良的黃曲霉菌株,并發(fā)現(xiàn)該菌株具備分泌胞外β-1, 3-1, 4-葡聚糖酶的能力。為深入了解該菌株發(fā)酵產(chǎn)β-1, 3-1, 4-葡聚糖酶的水平以及該酶的功能特性,本研究重點對該菌株產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的發(fā)酵條件進(jìn)行了優(yōu)化,并初步探索了該酶降解谷物內(nèi)β-葡聚糖的應(yīng)用潛能。
1 實驗部分
1.1 實驗材料
大麥葡聚糖,愛爾蘭Megzyme公司;燕麥粉、大麥粉,北京永輝超市;酵母提取物、胰蛋白胨,英國Oxoid公司;吐溫、曲拉通和十二烷基硫酸鈉(SDS)等,中國國藥集團(tuán)化學(xué)試劑有限公司;玉米苞皮、小麥麩皮、稻殼、玉米芯、白酒酒糟、花生殼等農(nóng)業(yè)廢棄物,北京官園花鳥市場;其他試劑如無特殊說明均為分析純。
黃曲霉(CICC No.2476,不產(chǎn)黃曲霉毒素)購買于中國工業(yè)微生物菌種保藏中心,保存于本研究室。
初始發(fā)酵培養(yǎng)基(g/L):玉米芯20,胰蛋白胨10,酵母提取物10,NaCl 5,MgSO4·7H2O 0.5,KH2PO40.75,自然pH,121 ℃滅菌20 min。
1.2 實驗儀器
PDA培養(yǎng)基(g/L):馬鈴薯200,葡萄糖20,瓊脂粉20,自然pH,121 ℃滅菌20 min。
MJPS- 150型霉菌培養(yǎng)箱,上海精宏有限公司;ZWF- 100型搖床,上海智誠分析有限公司;HH.S11- 4型電熱恒溫水浴鍋,上海博訊醫(yī)療有限公司;TGL- 20BR型高速冷凍臺式離心機,上海安亭科學(xué)儀器廠;TU- 1810PC型紫外可見分光光度計,北京普析通用儀器設(shè)備有限責(zé)任公司。
1.3 實驗方法
1.3.1發(fā)酵產(chǎn)酶
將黃曲霉劃線接種于PDA平板上,置于30 ℃下培養(yǎng)3~5 d。將大小為1 cm2黃曲霉菌塊接種到含50 mL初始發(fā)酵培養(yǎng)基的250 mL錐形瓶中。在37 ℃、200 r/min的條件下培養(yǎng)4 d,離心(10 000r/min,10 min)取上清液,測定β-1, 3-1, 4-葡聚糖酶酶活力。
1.3.2發(fā)酵產(chǎn)酶的優(yōu)化
采用單因素實驗優(yōu)化黃曲霉產(chǎn)酶條件。考察碳源對黃曲霉產(chǎn)酶的影響,包括碳源種類(玉米芯、麩皮、燕麥粉、大麥粉、稻草、玉米苞皮、白酒酒糟、花生殼、葡萄糖、木糖、蔗糖、乳糖、麥芽糖、可溶性淀粉和葡聚糖)及其添加量(質(zhì)量濃度)0~50 g/L對產(chǎn)酶的影響;考察氮源種類(胰蛋白胨、酵母提取物、蛋白胨、牛肉膏、大豆蛋白胨、豆粕粉、硫酸銨和磷酸氫二銨)及其添加量(質(zhì)量濃度)(0~35 g/L)對產(chǎn)酶的影響;考察表面活性劑種類(吐溫-20、吐溫- 40、吐溫-60、吐溫-80、曲拉通-110、曲拉通-114和SDS(十二烷基硫酸鈉))及其添加量(質(zhì)量濃度)(0~30 g/L)對產(chǎn)酶的影響。繼續(xù)優(yōu)化培養(yǎng)基初始pH值(5.0 ~ 8.5)和培養(yǎng)溫度(25~40 ℃,每隔5 ℃),基于較佳培養(yǎng)基組成和較優(yōu)培養(yǎng)條件的研究結(jié)果,考察該菌搖瓶發(fā)酵的產(chǎn)酶過程。
1.3.3β-1, 3-1, 4-葡聚糖酶酶活力及蛋白含量測定
酶活力測定參照Tang等[12]的方法:150 μL大麥葡聚糖(質(zhì)量濃度為5 g/100 mL)底物置于45 ℃下預(yù)熱3 min,加入50 μL適當(dāng)稀釋的酶液并在45 ℃下反應(yīng)10 min;加入200 μL DNS(3,5-二硝基水楊酸)煮沸15 min顯色,最后加入200 μL酒石酸鉀鈉溶液(質(zhì)量濃度為40 g/100 mL),待冷卻后測定A540 nm值,以葡萄糖為標(biāo)準(zhǔn)。β-1, 3-1, 4-葡聚糖酶酶活力單位(U)定義為:在上述條件下每分鐘水解大麥β-葡聚糖生成1 μmol/L葡萄糖所需要的酶量。
蛋白含量的測定參照Lowry等[13]的方法,以牛血清白蛋白為標(biāo)準(zhǔn)。
1.3.4黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶降解谷物中β-葡聚糖能力研究
水解條件:向1 g大麥粉/燕麥粉中加入9 mL MOPS(4-(N-morpholino) propanesulphonic acid)緩沖液(50 mmol/L,pH值7.5)混勻;加入10 U/mL的胞外β-1, 3-1, 4-葡聚糖酶進(jìn)行水解反應(yīng),不同時間點取樣并于沸水中蒸煮5 min滅活,10 000 r/min離心10 min后取上清液待測。
TLC檢測條件:采用德國Merk公司60F 254型號硅膠板,展層劑為正丁醇/乙酸/水,三者體積比為2∶1∶1,顯色劑為5%硫酸甲醇溶液,展層2次。
1.3.5響應(yīng)面優(yōu)化試驗
基于單因素研究,選取麩皮濃度、吐溫-60濃度和溫度3個因素為自變量,以酶活為響應(yīng)值,以單因素實驗所得最適結(jié)果為響應(yīng)面試驗的中心,進(jìn)行三因素三水平中心組合設(shè)計響應(yīng)面試驗(表1)。通過多元回歸擬合,并對回歸方程進(jìn)行方差分析及擬合度檢驗,討論響應(yīng)面特征,確定優(yōu)化的產(chǎn)酶條件。最終通過實驗進(jìn)行驗證,將實驗值與預(yù)測值進(jìn)行比較,驗證模型的有效性。
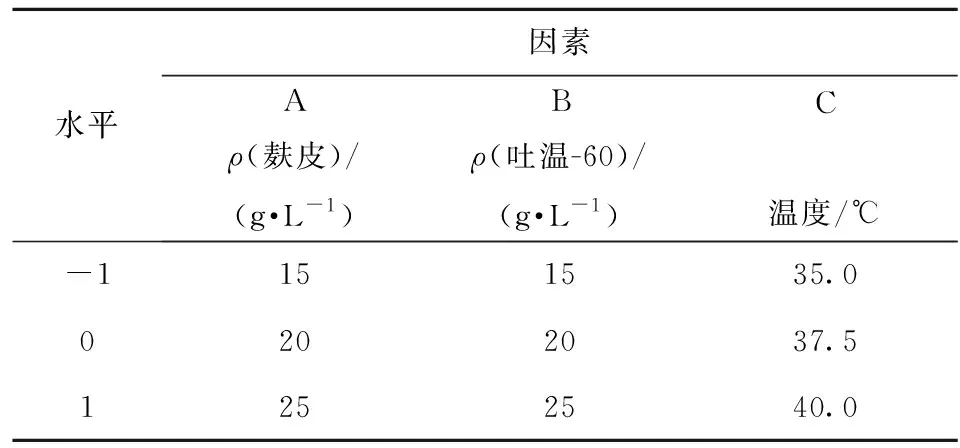
表1 響應(yīng)面設(shè)計的因素及水平
2 結(jié)果與分析
2.1 碳源對黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的影響
碳源對黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的能力有重要影響,實驗結(jié)果見表2。當(dāng)以麩皮為唯一碳源時,黃曲霉產(chǎn)酶能力最強,且在麩皮添加量為20 g/L時達(dá)到最高酶活,為35.5 U/mL。
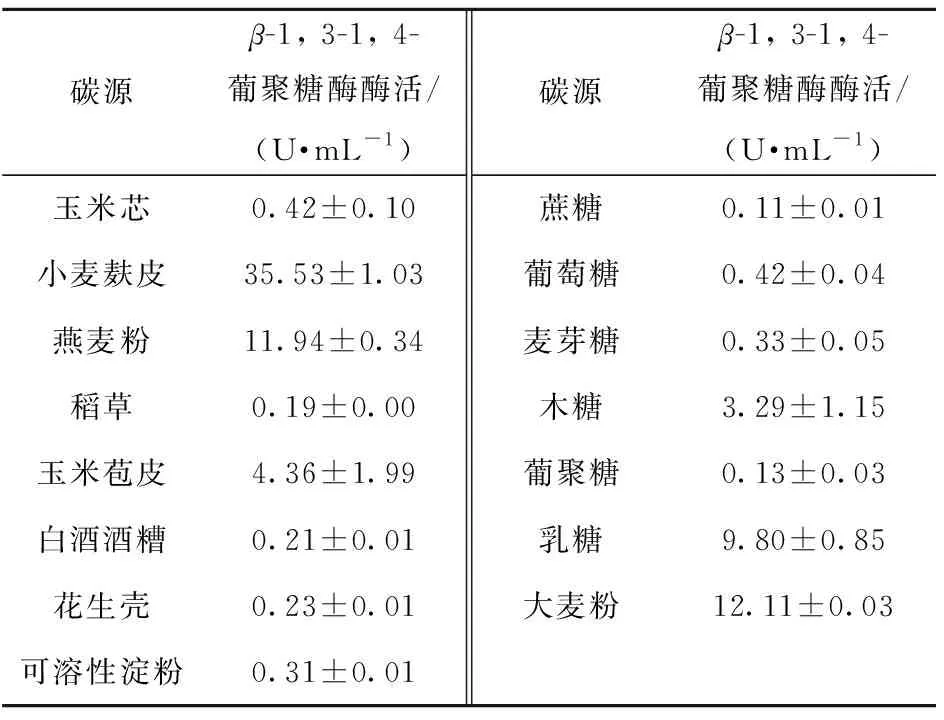
表2 碳源與黃曲霉發(fā)酵產(chǎn)β-1, 3-1, 4-葡聚糖酶的關(guān)系
碳源是微生物自身骨架合成過程中主要的營養(yǎng)物質(zhì)和能源物質(zhì),提供菌體生長所需的碳骨架和維持菌體生命活動所需的代謝能量,是影響菌體生長和代謝產(chǎn)物合成的重要因子[14]。不同的微生物由于代謝酶系可利用的碳源不同,且不同碳源條件下微生物產(chǎn)生的代謝產(chǎn)物具有一定的差異[15-16]。β-1,3-1, 4-葡聚糖酶是一種誘導(dǎo)型酶,不同碳源誘導(dǎo)其產(chǎn)酶能力不盡相同。本研究發(fā)現(xiàn),黃曲霉利用纖維質(zhì)原料誘導(dǎo)產(chǎn)酶的能力明顯強于其他單一糖類,可能是由于農(nóng)業(yè)廢棄物所富含的營養(yǎng)成分更豐富,利于黃曲霉的代謝利用。此外,農(nóng)業(yè)廢棄物價格低廉,用于發(fā)酵產(chǎn)酶還可以節(jié)約酶的生產(chǎn)成本。
2.2 氮源對黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的影響
氮源同樣能顯著影響黃曲霉產(chǎn)β-1, 3-1, 4-葡聚糖酶的能力,實驗結(jié)果見表3。由表3可知,當(dāng)以磷酸氫二銨為氮源時,黃曲霉的產(chǎn)酶水平最高,酶活可達(dá)68.3 U/mL。進(jìn)一步研究發(fā)現(xiàn),添加30 g/L的磷酸氫二銨最為合適,此時的酶活為79.1 U/mL。
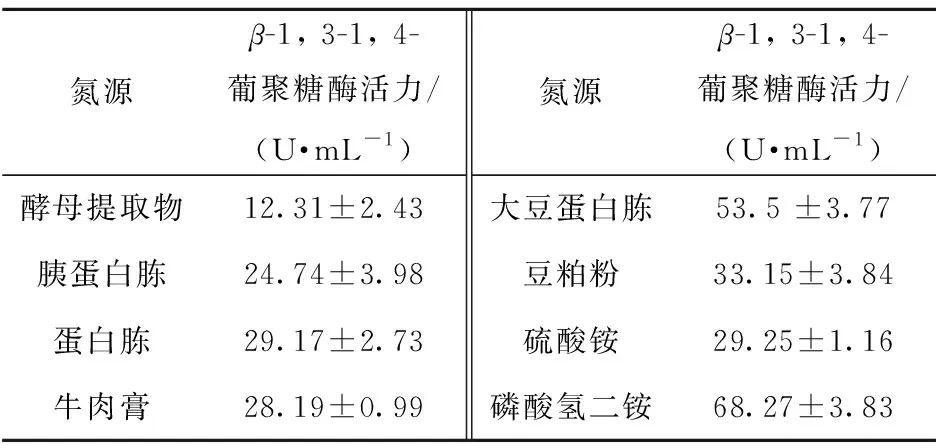
表3 氮源與黃曲霉發(fā)酵產(chǎn)β-1, 3-1, 4-葡聚糖酶的關(guān)系
2.3 表面活性劑對黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的影響
表面活性劑對黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的影響結(jié)果如圖1。非離子型表面活性劑對黃曲霉產(chǎn)酶的促進(jìn)作用明顯高于離子型表面活性劑,以吐溫-60的作用效果最好。添加20 g/L的吐溫-60時黃曲霉產(chǎn)酶水平最高,酶活可達(dá)123.3 U/mL。
吐溫-60的添加顯著促進(jìn)了黃曲霉產(chǎn)酶的能力,可能是由于吐溫-60能夠有效增強細(xì)胞膜的通透性,有助于胞內(nèi)酶透過細(xì)胞膜而分泌出來,從而提高胞外酶的產(chǎn)量[17]。另外,添加濃度的變化會對黃曲霉產(chǎn)酶能力造成較大影響,添加20 g/L的吐溫-60時產(chǎn)酶水平最高,但繼續(xù)增大吐溫-60的添加量,酶活又出現(xiàn)下降的趨勢,說明過高濃度的表面活性劑可能會一定程度上抑制菌株產(chǎn)酶。

圖1 表面活性劑與黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的關(guān)系Fig.1 Effect of surfactants on β-1, 3-1, 4-glucanase production
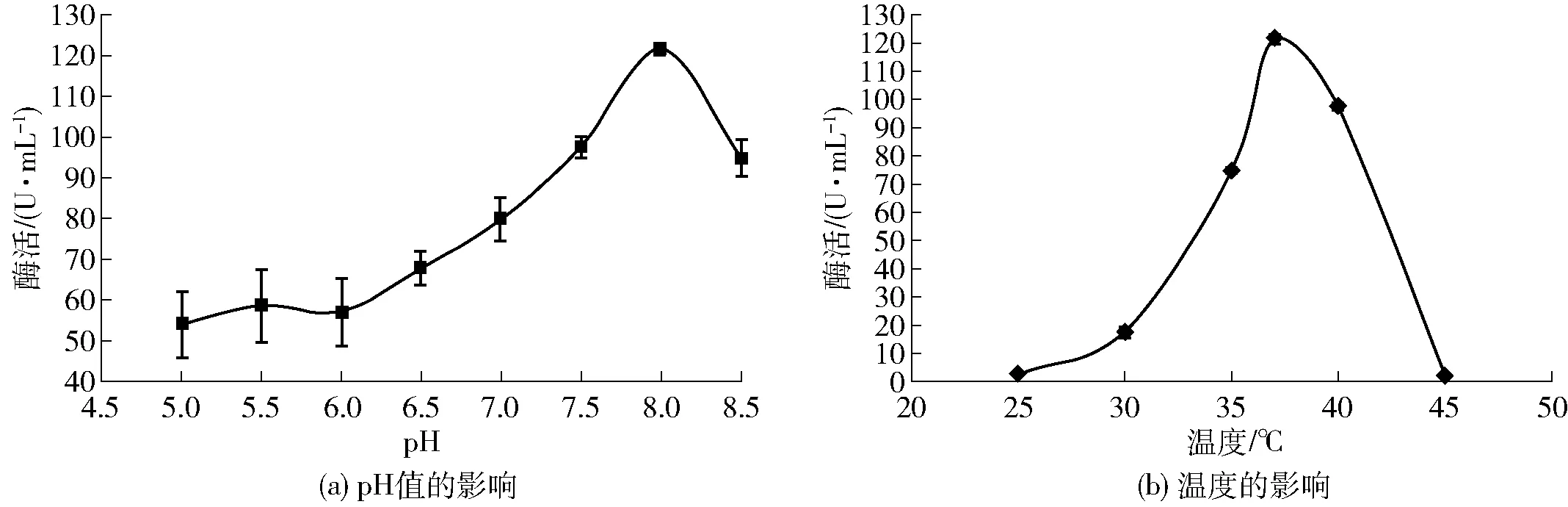
圖2 初始pH值和溫度與黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶能力的關(guān)系Fig.2 Effects of initial pH and temperature on β-1, 3-1, 4-glucanase production
2.4 培養(yǎng)條件對黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的影響
培養(yǎng)基的初始pH值及培養(yǎng)溫度對黃曲霉發(fā)酵產(chǎn)酶的影響較大,實驗結(jié)果見圖2。研究發(fā)現(xiàn),培養(yǎng)基初始pH值為8.0、發(fā)酵溫度選擇37 ℃時酶活最高,為121.5 U/mL。
pH值的變化會使微生物合成酶的類型和酶系的組成產(chǎn)生不同程度的改變,進(jìn)而影響微生物的代謝活動;因此pH值是影響微生物代謝產(chǎn)物的產(chǎn)量及活性的重要因素[18]。由圖2(a)可知,隨初始pH值的增加,酶活不斷增加,當(dāng)pH值升至8.0時酶活最高,繼續(xù)提高pH值時酶活又開始逐漸降低。溫度是影響微生物生長代謝的重要因素,隨著溫度的升高微生物的生長不斷加快,從而促進(jìn)微生物代謝以加快分泌酶等活性物質(zhì)。在37 ℃下培養(yǎng)黃曲霉,能夠分泌最高活力的胞外β-1, 3-1, 4-葡聚糖酶,但隨著溫度的繼續(xù)升高,酶活又開始出現(xiàn)較大幅度的下降。這可能是由于溫度的升高一定程度上抑制了菌株的正常生長,造成代謝異常從而導(dǎo)致了酶活的降低。
2.5 黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的過程分析
確定優(yōu)化培養(yǎng)基及培養(yǎng)條件后,最終考察了培養(yǎng)時間對黃曲霉產(chǎn)β-1, 3-1, 4-葡聚糖酶的影響(圖3)。β-1, 3-1, 4-葡聚糖酶的酶活力隨著培養(yǎng)時間的延長而增長,發(fā)酵至第6天時酶活力最高,達(dá)155.4 U/mL。
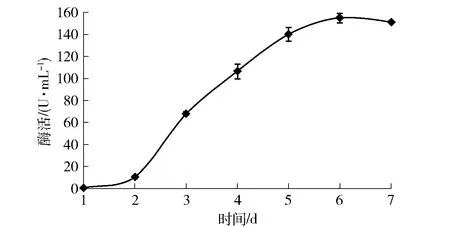
圖3 黃曲霉發(fā)酵產(chǎn)胞外β-1, 3-1, 4- 葡聚糖酶的產(chǎn)酶歷程Fig.3 Time course of β-1, 3-1, 4-glucanase production from A. flavus
目前,國內(nèi)外已報道的β-1, 3-1, 4-葡聚糖酶主要來源于細(xì)菌,如芽孢桿菌和類芽孢桿菌等[19-22],有關(guān)真菌產(chǎn)β-1, 3-1, 4-葡聚糖酶的研究還很少。曲霉菌是多數(shù)工業(yè)酶制劑生產(chǎn)的重要菌種,也是目前真菌源β-1, 3-1, 4-葡聚糖酶研究的主要對象,但關(guān)于黃曲霉分泌的β-1, 3-1, 4-葡聚糖酶的相關(guān)研究至今未見報道。
2.6 響應(yīng)面優(yōu)化結(jié)果分析
表4是依據(jù)響應(yīng)面設(shè)計的不同因素及水平的試驗結(jié)果。表5則是對模擬回歸系數(shù)的方差分析結(jié)果。

表4 響應(yīng)面試驗結(jié)果
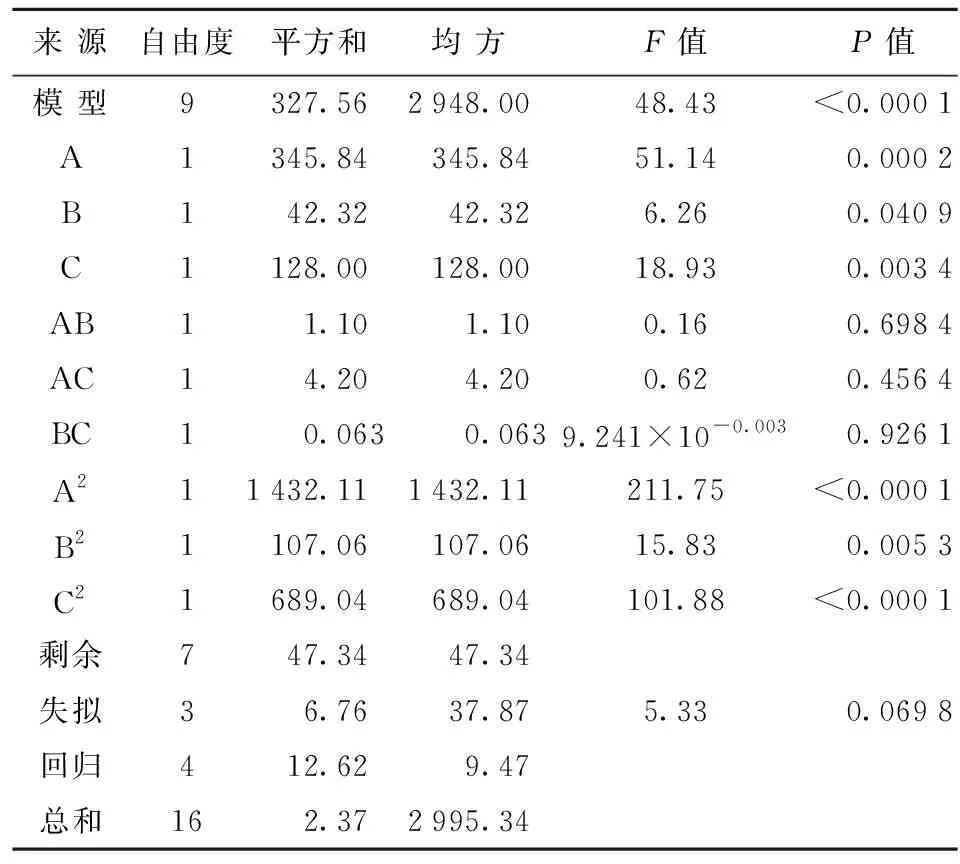
表5 響應(yīng)面試驗回歸分析
表4、表5結(jié)果表明,因變量和自變量間的線性關(guān)系明顯,線性回歸系數(shù)值P<0.05,表現(xiàn)為顯著性,說明該實驗方案可行。同時,失擬項系數(shù)值P為0.069 8(>0.05),表現(xiàn)為不顯著,表明試驗?zāi)P蛿M合度較好,誤差等偶然因素對試驗的影響不顯著。
進(jìn)一步分析了各因素交互作用,響應(yīng)面分析結(jié)果見圖4。通過考察3個因素相互間作用對響應(yīng)值的影響,再經(jīng)Design-Expert 8.0軟件分析獲得優(yōu)化條件為麩皮19.14 g/L、吐溫-60 21.09g/L、溫度37.87 ℃,相應(yīng)的酶活為156.9 U/mL。最終經(jīng)修正,選擇麩皮19.0 g/L、吐溫-60 21.0 g/L、溫度38 ℃進(jìn)行驗證實驗,測得酶活為155.9 U/mL,結(jié)果與預(yù)測值相當(dāng)。
經(jīng)單因素實驗和響應(yīng)面試驗優(yōu)化發(fā)酵條件后,黃曲霉產(chǎn)β-1, 3-1, 4-葡聚糖酶的最高酶活力達(dá)155.9 U/mL,產(chǎn)酶水平高于已報道的黑曲霉(20.0U/mL)[10],但低于泡盛曲霉的產(chǎn)酶水平[23]。與其他絲狀真菌相比,黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶的能力明顯高于樟絨枝霉和嗜熱擬青霉,但不如米黑根毛霉[12,24-25]。此外,本研究的黃曲霉還可利用農(nóng)業(yè)廢棄物(麩皮)來制備β-1, 3-1, 4-葡聚糖酶,這不僅有助于降低酶的生產(chǎn)成本,還利于增加農(nóng)業(yè)廢棄物的附加值。
2.7 黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶降解谷物內(nèi)β-葡聚糖的應(yīng)用潛力初探
β-1, 3-1, 4-葡聚糖酶能夠特異性水解β-葡聚糖,具備促進(jìn)富含β-葡聚糖類谷物營養(yǎng)吸收的功能。為研究黃曲霉β-1, 3-1, 4-葡聚糖酶的應(yīng)用潛能,重點考察了該酶降解大麥粉和燕麥粉水溶液(富含β-葡聚糖)的能力,實驗結(jié)果見圖5。由TLC分析結(jié)果顯示,該酶能夠直接降解大麥粉和燕麥粉中的可溶性β-葡聚糖;隨著水解反應(yīng)的進(jìn)行,水溶液中的可溶性β-葡聚糖最終被降解成葡萄糖和少量低聚糖。

圖4 各因素交互作用的響應(yīng)面分析結(jié)果Fig.4 Response surface plot showing effects of interactions among three factors on enzyme activity

G:葡萄糖圖5 黃曲霉產(chǎn)胞外β-1, 3-1, 4-葡聚糖酶降解谷物中葡聚糖的TLC分析Fig.5 TLC analysis of hydrolytic products of glucan from grain by β-1, 3-1, 4-glucanase from A. flavus
微生物源β-1, 3-1, 4-葡聚糖酶的主要功能是降解β-葡聚糖并生產(chǎn)纖維寡糖。纖維寡糖作為一種益生元,具有調(diào)節(jié)腸道菌群平衡的特殊功效,但纖維寡糖卻并不能被機體消化和吸收,還有可能造成機體腹部脹氣等不適反應(yīng)。本研究發(fā)現(xiàn),黃曲霉分泌的胞外β-1, 3-1, 4-葡聚糖酶不但能實現(xiàn)天然谷物中β-葡聚糖的有效降解,還能將β-葡聚糖降解成小分子的葡萄糖。這一獨特屬性對于該酶在葡萄糖制備和生產(chǎn)領(lǐng)域,以及促進(jìn)人類或者動物機體消化、吸收富含β-葡聚糖類的谷物具有重要實用價值。
3 結(jié) 論
本研究通過單因素實驗和響應(yīng)面分析法優(yōu)化了黃曲霉產(chǎn)β-1, 3-1, 4-葡聚糖酶的液體發(fā)酵條件,得到優(yōu)化的發(fā)酵條件:麩皮19 g/L、磷酸氫二銨30 g/L、吐溫-60 21 g/L、NaCl 5 g/L、MgSO4·7H2O 0.5 g/L、KH2PO40.75 g/L、培養(yǎng)基初始pH值8.0、培養(yǎng)溫度38 ℃、培養(yǎng)時間6 d。在此條件下發(fā)酵,黃曲霉能產(chǎn)155.9 U/mL的β-1, 3-1, 4-葡聚糖酶,其產(chǎn)酶水平高于多數(shù)國內(nèi)外報道的絲狀真菌。麩皮是小麥制粉后的副產(chǎn)品,資源豐富且成本低廉,是微生物工業(yè)發(fā)酵的良好碳源。本研究發(fā)現(xiàn)黃曲霉可利用麩皮作為碳源生產(chǎn)β-1, 3-1, 4-葡聚糖酶,有效降低了生產(chǎn)成本。另外,水解實驗發(fā)現(xiàn),黃曲霉分泌的β-1, 3-1, 4-葡聚糖酶具有獨特水解性能,不僅能有效降解天然谷物粉中的可溶性β-葡聚糖,還能將這些β-葡聚糖降解成葡萄糖,這對于促進(jìn)天然谷物的消化和吸收具有重要意義。
猜你喜歡
作文·小學(xué)低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學(xué)生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學(xué)低年級(2024年2期)2024-04-29 00:00:00
作文·小學(xué)低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
