IL-33/ST2信號通路與脂肪代謝關系研究進展
2019-01-03 05:16:20成萍
河南農業科學 2018年12期
, ,成萍, ,,,*
(1.聊城大學 生命科學學院,山東 聊城 252059; 2.聊城大學 藥學院,山東 聊城 252059)
肌內脂肪(Intramuscular fat,IMF)是衡量肉品質的重要指標,能夠影響肉的嫩度和風味,并且IMF所反映的大理石紋等級直接影響肉質量等級,決定了肉產品的經濟性狀。肌內脂肪是脂肪沉積的主要形式,脂肪組織通過代謝和細胞的內分泌功能在調節能量平衡中起著至關重要的作用。脂肪細胞分化是受多種細胞因子或信號通路調控的復雜網絡體系。
IL-33/ST2(Interleukin-33/interleukin-1 receptor like 1)是近年來新發現的能夠調控白色脂肪米色化、棕色或米色脂肪產熱的重要信號通路[1-3]。因此,闡明脂肪細胞分化、IL-33/ST2信號通路及IL-33/ST2信號通路在脂肪代謝中的作用機制,旨在為肥胖癥、代謝綜合征、胰島素糖尿病等代謝相關疾病的治療提供理論指導。
1 脂肪細胞分化
脂肪細胞分化過程包括2個階段,第1個階段是胚胎干細胞或脂肪組織中的間充質干細胞(Mesenchymalstem cells,MSCs)分化為脂肪祖細胞并進一步形成前體脂肪細胞,第2個階段是前體脂肪細胞終末分化為成熟脂肪細胞[4-5]。成熟脂肪細胞具有參與脂類合成、轉運、能量消耗以及生產能量平衡調節相關脂肪因子的功能。研究表明,在脂肪分化期間有許多成脂分化調控因子發揮著關鍵作用,例如CCAAT/增強子結合蛋白家族(CCAAT/enhancer-binding protein family,C/EBPs)、骨形成蛋白(Bone morphogenetic proteins,BMPs)、過氧化物酶體增殖物激活受體(Peroxisome proliferators-activated receptors,PPARs)[6-8]。
脂肪組織是機體調節能量平衡的中心,哺乳動物中存在2種不同功能類型的脂肪組織,分別為白色脂肪組織(WAT)和棕色脂肪組織(BAT)。WAT能夠作為儲存庫將過多的能量以甘油三酯的形式儲存起來,BAT能夠在寒冷和應激條件下為機體產生熱量。有關脂肪細胞分化的信號轉導機制已被廣泛研究,其中C/EBP家族成員、PPARγ和cAMP效應元件結合蛋白(cAMP-response element binding protein,CREB)是白色脂肪細胞特異性轉錄級聯中的關鍵因子,在脂肪細胞成熟期的早期階段,上調C/EBPβ和C/EBPδ蛋白能夠增強PPARγ和C/EBPα的表達,隨后PPARγ分別與C/EBPα、C/EBPβ構成反饋回路,進一步促進白色脂肪細胞的終末分化。BMP7能夠激活形成棕色脂肪細胞的完整程序,包括誘導早期調節因子PRDM16(PR domain-containing 16)表達,增強脂肪轉錄因子PPARγ和C/EBPs表達,并且促進p38促分裂原活化蛋白激酶(Mitogen-activated protein kinase,MAPK)途徑的活化[9]。因此,進一步闡明IL-33/ST2信號通路在脂肪細胞分化及脂肪代謝中的功能具有重要意義。
2 IL-33/ST2信號通路
2.1 ST2的變異體及其調控機制
白介素1受體樣1(IL1RL1,習慣稱之為ST2)是白介素1受體家族成員之一,在1989年最先被命名,多年來一直被認為是與免疫和炎癥相關的孤兒受體[10]。人的ST2基因有4個轉錄變異體,其中2個變異體是最重要的,分別為跨膜ST2受體(ST2L,也被稱為IL1RL1-b)和血清可溶性ST2受體(sST2,也被稱為IL1RL1-a)。ST2的選擇性啟動子以及其mRNA的3′端選擇性剪接,導致了sST2和ST2L變異體的產生[11]。ST2具有1個近端啟動子和1個遠端啟動子,它們能夠影響ST2基因的轉錄調控機制,但是對sST2和ST2L的轉錄調控還不完全清楚[12]。在Schmitz等[13]報道中提及白介素-33(IL-33)是ST2的配體,能夠參與sST2和ST2L的轉錄調控,這為更好地理解sST2和ST2L的功能提出了思路。
sST2的產生受多種信號通路的調控。研究表明,在sST2存在的情況下,使用IL-33處理心肌細胞后,觀察到血管緊張素Ⅱ和苯腎上腺素的促肥大效應被阻斷[14]。若在肺泡上皮細胞和心肌細胞中,用特殊的抑制劑CAPE對核因子-κB(Nuclear factor-κB,NF-κB)信號通路進行阻斷,可以阻止這些細胞產生sST2[15]。在人類乳腺癌細胞系MCF-7中,成纖維細胞生長因子2可通過MEK/ERK信號通路,促進sST2的產生[16]。在人類支氣管上皮細胞中,溶血磷脂酸能夠通過一種依賴于NF-κB或JNK的方式來增加sST2的表達量[17]。
IL-33/ST2信號通路能夠激活Ⅱ型CD4+T細胞(Type 2 CD4+T-helper cells,Th2)效應細胞,并釋放Th2相關細胞因子來參與炎癥和免疫反應。在這個過程中,ST2L能夠調節IL-33對Th2的炎癥作用,而sST2則與Th2炎癥反應的衰減相關。例如,在小鼠的模型試驗中發現,IL-33能夠通過與ST2L的相互作用,減少心肌纖維化和細胞凋亡、防止心肌細胞肥大,從而達到對心肌細胞的保護[14]。
2.2 IL-33與ST2的相互作用
IL-33作為新發現的細胞因子,是IL-1家族的新成員之一,能被大多數細胞分泌,機體多以其應對組織損傷[18]。IL-33通過與膜表面受體ST2L和IL-1R輔助蛋白(IL-1 receptor accessory protein,IL-1RAP)結合,組成受體復合物來發揮細胞功能。IL-33結合ST2L時,IL-1RAP能夠增強IL-33對ST2L的親和力,是后續信號傳導順利進行的必要條件[19]。IL-33和ST2L的相互作用能夠激活MAPK和幾種生化途徑,主要是由于NF-κB激酶復合體抑制子的激活,觸發了NF-κB的活性所致[20]。但也有人認為,IL-33可能具有不依賴于ST2L受體的胞內功能。另有研究發現,sST2與IL-33結合會中斷IL-33和ST2L之間的相互作用,從而導致它們在細胞中的功能被阻斷,因此,sST2也被認為是一種誘騙受體(Decoy receptor)(圖1)。在這種情況下,ST2系統不但能夠以ST2L跨膜異構體的形式作為IL-33行使功能的介導者,而且可以通過sST2異構體的形式抑制IL-33的功能。另外,IL-33也能夠參與調控ST2L和sST2的mRNA轉錄,它能夠增加ST2L的mRNA表達量,并降低sST2的mRNA表達量[22]。
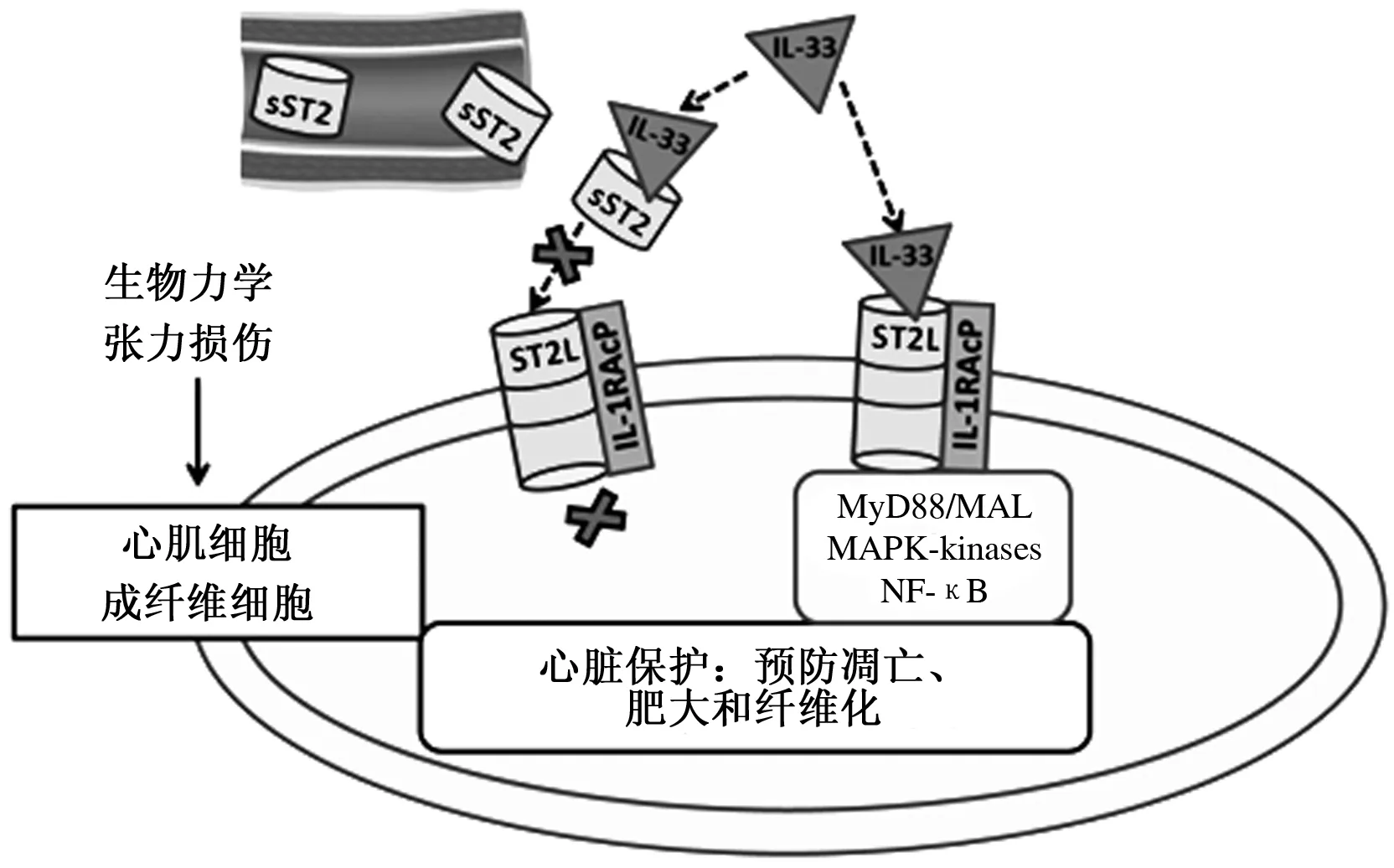
參考文獻[21]
肥胖與代謝紊亂及許多健康問題密切相關,是導致糖尿病、心血管疾病及部分癌癥表型的主要危險因素[23]。關于IL-33/ST2與肥胖的關系,已有報道表明,在脂肪組織和肝臟中表達的IL-33/ST2對肥胖有一種“自然”的保護作用,原因是IL-33/ST2途徑的激活增加了抗炎性細胞因子并減少了脂肪細胞分化和脂肪儲存[24]。為研究IL-33/ST2對肥胖影響的分子機制,分別對心臟、脂肪組織和肝臟中IL-33/ST2系統mRNA的表達情況進行檢測,發現與對照組相比,肥胖Zucker大鼠IL-33的mRNA表達水平在脂肪組織中顯著偏低,ST2L的mRNA表達水平在肝臟中顯著偏低,sST2的mRNA表達水平在所有供試組織中均顯著偏低,因此,肥胖Zucker大鼠組織可以通過減少sST2的表達來促進IL-33對膜受體ST2L的作用,以形成一種肥胖保護機制[25]。但是,目前尚不知IL-33/ST2信號通路具體是以何種方式或途徑來調控脂肪細胞分化的。
3 IL-33/ST2與脂肪細胞代謝
3.1 IL-33及其相關基因與白色脂肪米色化
免疫細胞和WAT的米色化(Beiging)關系緊密,Ⅱ型先天淋巴細胞(ILC2)最初是在脂肪相關淋巴簇中發現的[26],對于招募和維護駐留在WAT中的嗜酸性粒細胞至關重要[27]。IL-33能夠刺激ILC2產生大量的Ⅱ型細胞因子IL-5和IL-13,繼而導致WAT的米色化。關于ILC2誘導米色脂肪細胞產生的機制,近期研究已經做了很好的解釋。使用ILC2的興奮劑IL-33處理后,會導致小鼠WAT中ILC2數量增加,同時米色脂肪細胞和耗氧量也隨之增加,與之相反的是,IL-33的處理對BAT沒有影響;進一步研究發現,IL-33-/-型小鼠的體質量比對照組明顯增加,機體內WAT明顯增多,但ILC2和米色脂肪細胞的數量均減少;重組無淋巴小鼠(Rag2-/-Il2rg-/-)在只有ILC2的情況下就足以導致WAT的米色化,從而顯示了IL-33和ILC2在調控脂肪組織生長中的重要作用[2-3]。此外,IL-13誘騙受體(IL-13 decoy receptor,IL-13Rα2)能夠通過調控內源IL-13的表達水平,限制IL-33在脂肪組織中的調控作用。相關研究發現,使用IL-33處理后,小鼠血清和組織中IL-13的含量增加,并伴隨著嗜酸性粒細胞、巨噬細胞(Alternatively activated macrophages,AAMs)和Ⅱ型固有淋巴樣細胞的含量升高,同時體質量、脂肪量和空腹血糖水平隨之降低,上述癥狀在IL-13Rα2缺陷型小鼠中更為突出,但在IL-13缺陷型小鼠中卻呈現減弱趨勢;除此之外,還檢測到在肥胖小鼠脂肪組織中IL-13Rα2表達量較高,在IL-13Rα2缺陷型小鼠中IL-13的表達量較高[28]。因此,可以說明IL-13Rα2能夠限制IL-33/IL-13軸在肥胖中的保護作用,可作為脂肪組織中的關鍵檢驗點。
ILC2控制WAT米色化的不同機制如圖2所示,當ILC2被IL-33刺激后,可以通過多種方式調控脂肪組織的生物學功能。首先,被刺激后的ILC2能夠產生Ⅱ型細胞因子IL-5,以激活WAT中的嗜酸性粒細胞,從而選擇性激活AAMs[30-31]。與此同時,被激活的嗜酸性粒細胞能夠觸發并產生IL-4,作為對IL-4的應答,AAMs能夠產生去甲腎上腺素,從而導致脂肪組織的米色化[1]。此外,Lee等[3]從脂肪祖細胞角度闡明了米色脂肪細胞產生的機制,即當IL-4受體(IL-4Rα)的組織特異性缺失時,接收到該信號的IL-4可繞過AAM-去甲腎上腺素軸直接作用于脂肪祖細胞,這是由于在IL-4的刺激下,能夠上調脂肪祖細胞中米色脂肪細胞分化相關基因的表達,但IL-4Rα信號在已分化的脂肪細胞中是無用的。并且有趣的是,IL-4Rα不但能為IL-4傳遞信號,而且還能結合IL-13,即使在缺乏嗜酸性粒細胞產生IL-4的情況下,也會有大量的脂肪祖細胞產生。因此,由ILC2產生的IL-13可以通過與IL-4Rα的結合,直接作用于脂肪祖細胞,并能與嗜酸性粒細胞生成的IL-4形成協同作用,從而促進米色脂肪細胞的產生與發育。但在米色脂肪細胞產生的過程中,關于IL-4和IL-13的協同作用機制以及兩者之間的相對貢獻仍有待進一步研究。
Brestoff等[2]對ILC2和ILC3所表達的部分肥胖相關基因的比較研究發現,前蛋白轉化酶枯草溶菌素1(Proprotein convertase subtilisin/kexin type 1,PCSK1)以及它的靶基因PENK(Proenkephalin A)在ILC2中表達量較高。其中,蛋氨酸腦啡肽(Methionine enkephalin,MetEnk)是PENK加工后的一個產物,由ILC2產生。當ILC2受到IL-33的刺激時,MetEnk的產量隨之增加。利用MetEnk處理供試動物,會導致WAT耗氧量升高以及米色脂肪細胞標記物增溫素——UCP1的表達量增加。同時還發現,MetEnk的δ1阿片受體(δ1 opioid receptor,Oprd1)在供試動物腹股溝的WAT中表達量較高。這些數據表明,ILC2可以通過分泌MetEnk,直接促進WAT轉化為米色脂肪,而在這個過程中,WAT中的Oprd1受體作為感應媒介,能夠誘導并促進米色脂肪細胞生成相關基因的表達。但是在MetEnk結合Oprd1受體直接促進WAT米色化的過程中,IL-4和IL-13的表達量均無顯著變化,因此,這種表型很可能與IL-4和IL-13無關,但具體的作用機制還不明確。除此之外,該途徑中哪些細胞是MetEnk的靶細胞,仍需進一步探究。
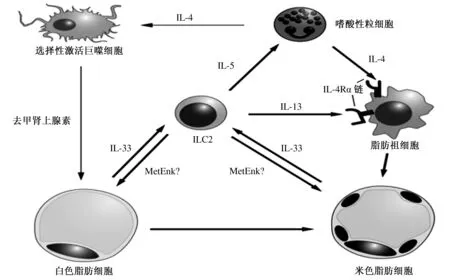
參考文獻[29]
雖然已經探索得知部分關于ILC2對小鼠WAT米色化的作用機制,但相關的機制是否也存在于人類中還不太清楚。Brestoff等[2]的研究結果表明,在瘦人的脂肪組織中ILC2構成了譜系標記陰性細胞的大部分。而Zeyda等[24]研究發現,在肥胖人的脂肪組織中IL-33的表達量增加,而在肥胖人的脂肪組織以及高脂肪飲食的小鼠脂肪組織中ILC2含量卻大大減少。對于這些相互矛盾的研究結果,推測可能是由于在肥胖的情況下,ILC2對IL-33的響應是失調的,但具體的作用機制還有待于進一步探究。Wood等[32]研究指出,人類WAT的細胞可以產生IL-33,也可以通過促炎刺激(如TNFα)來增加IL-33。然而,是否脂肪細胞本身是IL-33的主要來源,以及ILC2如何影響這種內源性反饋機制還有待證實。最后,發現如何利用這些途徑來增加體內IL-33的水平將是非常令人興奮的研究,并為肥胖癥的新療法鋪平道路。
3.2 IL-33/ST2與棕色或米色脂肪產熱
IL-33及冷刺激能夠促進ILC2的招募和激活,在棕色或米色脂肪產熱中也扮演著重要的角色。IL-33的表達量在高脂飲食和瘦素缺乏性肥胖條件下會顯著降低,這是由于WAT中ILC2以及嗜酸性粒細胞的含量降低所致。相應的使用IL-33處理高脂肪飲食小鼠,ILC2和嗜酸性粒細胞的數量以及UCP1的表達量會顯著增加。冷刺激也能夠誘導WAT中IL-33和UCP1的表達,從而增加ILC2和嗜酸性粒細胞的數量。但如果中和了IL-33信號,便可逆轉冷刺激的促進作用,例如通過局部注射6-羥基多巴胺(6-OHDA)切斷交感神經可顯著抑制冷刺激誘導的IL-33和ILC2/嗜酸性粒細胞途徑。盡管Brestoff等[2]和Lee等[3]的研究在很大程度上具有相同的觀點,但與Lee等不同的是,Brestoff等利用IL-33對IL-4Rα-/-動物進行處理后,UCP1+脂肪細胞的數量增加,從而表明IL-33可以直接作用于WAT的米色化過程,而與IL-4信號無關。除此之外,激活ILC2所產生的MetEnk可以直接與WAT中的Oprd1受體結合,也能促進米色脂肪細胞生成及產熱。
Odegaard等[33]也從另一個角度證實了IL-33/ST2信號通路可作為小鼠圍產期脂肪產熱的“許可證”,研究表明,IL-33是一種能激活Ⅱ型免疫反應的細胞因子,在棕色和米色脂肪細胞的解偶聯呼吸中具有“開關”作用。如圖3所示,在IL-33或ST2缺失的情況下,棕色脂肪細胞能夠正常發育,但不能表達出正常的UCP1 mRNA剪接轉錄本,導致UCP1蛋白缺失、解耦聯呼吸和溫度調節功能受損。
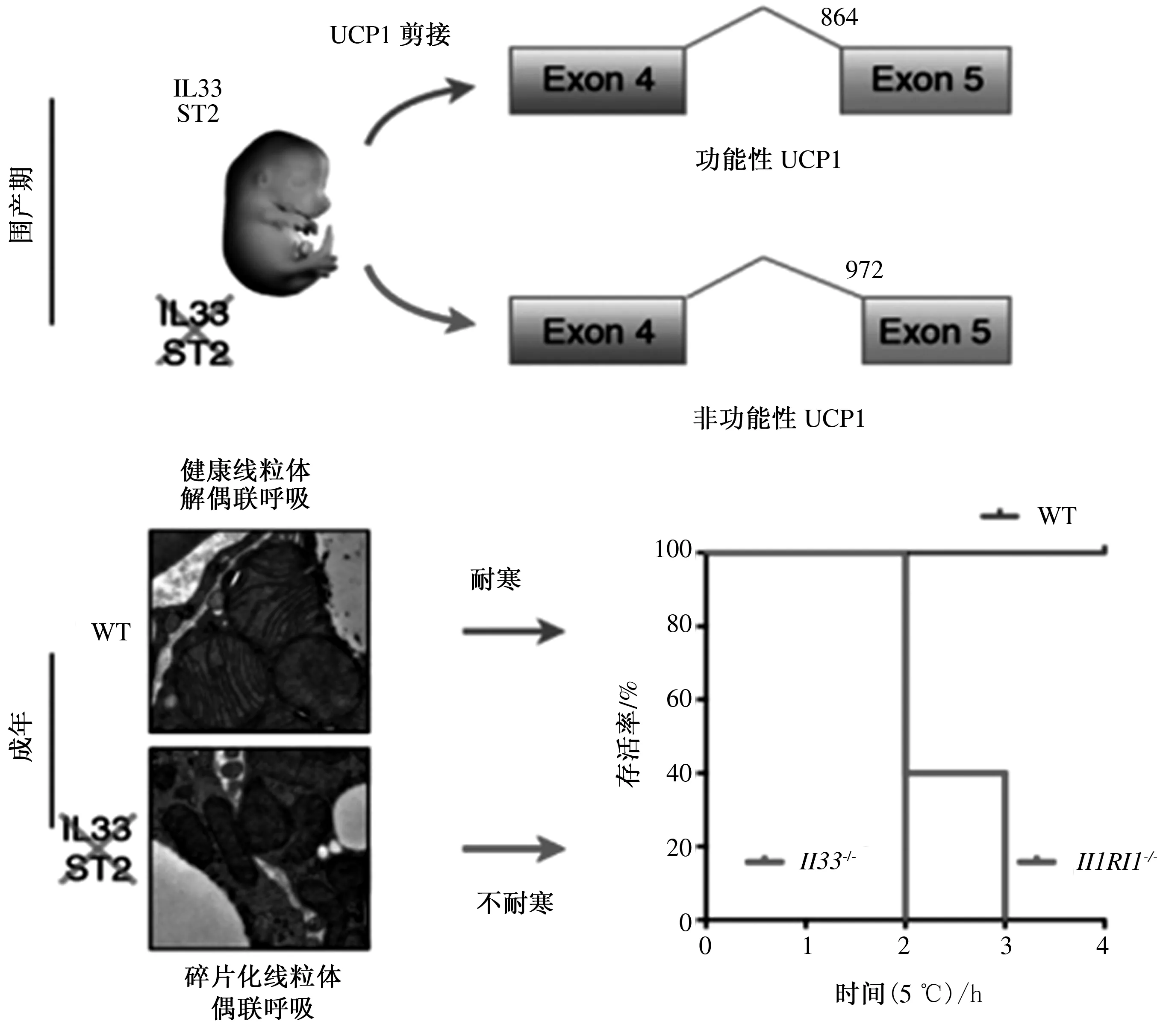
參考文獻[33]
4 小結
目前對脂肪沉積分子機制的理解,尤其是對前體脂肪細胞到成熟脂肪細胞分化過程中調控機制的理解仍然是有限的。鑒于IL-33/ST2L/sST2蛋白在不同生物系統的不同細胞中具有不同的功能,涉及范圍廣泛,IL-33/ST2信號通路在脂肪代謝中的研究是目前一個新興的研究熱點,關于IL-33/ST2信號通路在前體脂肪細胞分化過程中的功能及調控機制也尚不清楚。因此,該通路在脂肪分化中還有一系列的科學問題有待進一步研究:(1)在IL-33和ST2不同剪接變異體中,哪些變異體具有調控前體脂肪細胞分化的功能;(2)IL-33/ST2信號通路引發哪些下游基因的表達來調控前體脂肪細胞分化;(3)IL-33/ST2信號通路調控前體脂肪細胞分化的作用機制是什么。對于IL-33/ST2信號通路在脂肪細胞分化過程中的功能及作用機制的研究,將對深入了解脂肪沉積的調控機制產生重要的指導意義。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年3期)2019-02-01 06:12:26
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
