醛酮還原酶AKR7A3原核表達(dá)條件的優(yōu)化及酶學(xué)活性檢測(cè)
2018-10-31 02:05:50孫婷婷孫基豐周紫薇褚春旭張昊
孫婷婷,孫基豐,周紫薇,褚春旭,張昊
(長(zhǎng)春理工大學(xué) 生命科學(xué)技術(shù)學(xué)院,長(zhǎng)春 130022)
AKR作為NADPH依賴的氧化還原酶[1]家庭成員當(dāng)中的一種,AKR家族的存在十分廣泛,在原核和真核生物當(dāng)中都存在該種類型的酶[2]。文獻(xiàn)表明人源AKR家族種類已發(fā)現(xiàn)了15個(gè)[3]AKR酶家族主要參與生物機(jī)體內(nèi)醛酮類化合物的代謝過(guò)程,并在醛類和相關(guān)化合物的解毒時(shí)期發(fā)揮著不可忽視的作用。AKR家族酶的底物主要有脂肪族和芳香族的醛/酮類的化合物[4]。所以該酶在生成激素、藥物的分解排出、炎癥反應(yīng)、解毒等生物活動(dòng)中有著不可忽視的作用[5]。AKR大多以37kDa的單體類型出現(xiàn)在細(xì)胞漿中[6-8]。不同種類間的分布以及作用是不一樣的,比如說(shuō)AKR6家族能夠控制K+通道的開(kāi)放,AKR1、AKR6、AKR7三個(gè)種類在哺乳動(dòng)物機(jī)體中均有表達(dá),其功能是參加生物的生命代謝過(guò)程[9]。有關(guān)報(bào)道顯示,AKR7亞類型中的成員AKR7A2和AKR7A3主要參加肝臟對(duì)黃曲霉毒素的解毒的過(guò)程,其中AKR7A3主要是參與肝臟中AFB1類的毒性解除作用[10]。但是關(guān)于AKR7A3酶學(xué)活性的相關(guān)研究很少。
大腸桿菌是科研實(shí)驗(yàn)中最常用的高效表達(dá)異源蛋白的原核表達(dá)工具,其優(yōu)點(diǎn)在于操作簡(jiǎn)便,培養(yǎng)條件簡(jiǎn)單[11]。但并不是所有外源遺傳物質(zhì)都能夠在這種表達(dá)體系中進(jìn)行有效表達(dá)[12],主要由于不同的遺傳物質(zhì)有不同構(gòu)造、mRNA的穩(wěn)定性[13]、翻譯、折疊、宿主細(xì)胞蛋白酶的降解作用,大腸桿菌密碼子在原核和真核基因使用上的區(qū)別,以及蛋白質(zhì)對(duì)宿主的產(chǎn)生相對(duì)的毒性[14]等因素。因此,本文對(duì)于人源AKR7A3基因序列進(jìn)行相應(yīng)的密碼子優(yōu)化,構(gòu)建原核表達(dá)體系,優(yōu)化表達(dá)條件,獲得最大產(chǎn)量的可溶性酶蛋白,再通過(guò)酶促動(dòng)力學(xué)實(shí)驗(yàn),獲得最優(yōu)酶活性條件。本文實(shí)驗(yàn)結(jié)果將為后續(xù)AKR7A3研究提供實(shí)驗(yàn)基礎(chǔ)。
1 材料與方法
1.1 AKR7A3基因合成
從GENE BANK里面獲取AKR7A3全序列,選取相應(yīng)酶切位點(diǎn),由上海生工合成并優(yōu)化得到AKR7A3-pET15b質(zhì)粒。
1.2 菌株和試劑
大腸桿菌(Escherichia coli)表達(dá)菌株BL21(DE3),由本實(shí)驗(yàn)室制備并保存;IPTG(異丙基硫代半乳糖苷)配成100mg/mL濃度備用。低相對(duì)分子質(zhì)量蛋白Marker和SDS-PAGE試劑購(gòu)自寶生物工程大連公司(進(jìn)口分裝);其他試劑均為國(guó)產(chǎn)分析純?cè)噭?/p>
1.3 AKR7A3原核表達(dá)
將重組質(zhì)粒AKR7A3-pET15b和pET 15b空載體分別進(jìn)行誘導(dǎo)表達(dá),于37℃培養(yǎng)至OD600約為0.6,加入IPTG誘導(dǎo)5h。菌體離心后棄上清,得到的菌體沉淀用100μL PBS重懸后進(jìn)行分離純化。蛋白樣品分別加入100μL 2×SDS PAGE上樣緩沖液,沸水中處理5min,12000rpm/min瞬時(shí)離心后取20μL上清進(jìn)行SDS聚丙烯酰胺凝膠電泳。以pET 15b空載體和未加IPTG的AKR7A3-pET15b菌為對(duì)照鑒定目的基因AKR7A3的表達(dá)情況。
1.4 溫度對(duì)重組菌AKR7A3表達(dá)的影響
BL21-pET15b-AKR7A3在轉(zhuǎn)速180rpm、5h、誘導(dǎo)劑終濃度為1mM/L條件不變,溫度分別為37℃、30℃、24℃、16℃的環(huán)境下進(jìn)行表達(dá)。將其菌體進(jìn)行收集、超聲破碎,將得到的上清進(jìn)行SDS聚丙烯酰胺凝膠電泳檢測(cè)。
1.5 轉(zhuǎn)速對(duì)重組菌AKR7A3表達(dá)的影響
BL21-pET15b-AKR7A3在溫度 37℃,5h,誘導(dǎo)劑終濃度為1mM/L條件不變,轉(zhuǎn)速分別為110rpm、130rpm、150rpm、180rpm的條件下進(jìn)行表達(dá)。將其菌體進(jìn)行收集、超聲破碎,離心使用上清進(jìn)行SDS聚丙烯酰胺凝膠電泳檢測(cè)。
1.6 時(shí)間對(duì)重組菌AKR7A3表達(dá)的影響
BL21-pET15b-AKR7A3在37℃、180rpm,誘導(dǎo)劑終濃度為1mM/L條件不變,表達(dá)時(shí)間分別為5h、10h、16h、20h的條件下進(jìn)行表達(dá)。將其菌體進(jìn)行收集、超聲破碎,離心使用上清進(jìn)行SDS聚丙烯酰胺凝膠電泳檢測(cè)。
1.7 誘導(dǎo)劑濃度對(duì)重組菌AKR7A3表達(dá)的影響
BL21-pET15b-AKR7A3在37℃、180rpm,5h條件不變,誘導(dǎo)劑終濃度分別為0.40mM/L、0.60mM/L、0.80mM/L、1.00mM/L環(huán)境下進(jìn)行表達(dá)。將其菌體進(jìn)行收集、超聲破碎,離心使用上清進(jìn)行SDS聚丙烯酰胺凝膠電泳檢測(cè)。
1.8 醛酮還原酶AKR7A3酶活性動(dòng)力學(xué)檢測(cè)
1.8.1 最佳反應(yīng)溫度確定
酶反應(yīng)體系300μL;DL-甘油醛作為底物;NADPH作為體系輔酶;2%的乙腈為共溶劑;100mM/L PBS為反應(yīng)的緩沖溶液;PH需要控制在7.0;設(shè)置反應(yīng)溫度為20℃、25℃、30℃、37℃以及40℃,反應(yīng)時(shí)間達(dá)到5分鐘,反應(yīng)后在340nm處測(cè)定吸光數(shù)值。
1.8.2 最佳反應(yīng)PH值確定
酶反應(yīng)體系300μL;DL-甘油醛為底物;NADPH作為體系輔酶;2%的乙腈為共溶劑;100mM/L PBS為反應(yīng)的緩沖溶液;反應(yīng)溫度設(shè)置為37℃;將其PH值設(shè)置為5.00、6.00、7.00、8.00以及9.00,反應(yīng)時(shí)間達(dá)到5分鐘,反應(yīng)后在340nm.處測(cè)定吸光數(shù)值。
1.8.3 最佳反應(yīng)NADPH輔酶濃度確定
酶反應(yīng)體系300μL;DL-甘油醛為底物;NADPH作為體系輔酶;2%的乙腈為共溶劑;100mM/L PBS為反應(yīng)的緩沖溶液;反應(yīng)溫度為37℃;PH設(shè)置為7.00,NADPH輔酶濃度設(shè)置為0.10mmol/L、0.15mmol/L、0.20mmol/L、0.30mmol/L、0.40mmol/L,反應(yīng)時(shí)間達(dá)到5分鐘,反應(yīng)后在340nm處測(cè)定吸光數(shù)值。
1.8.4 最佳反應(yīng)底物濃度確定
酶反應(yīng)體系300.00μL;DL-甘油醛為底物;NADPH作為體系輔酶;2%的乙腈為共溶劑;使用100mM/L PBS作為反應(yīng)的緩沖溶液;反應(yīng)溫度為37℃;PH控制在7.0;底物濃度分別為2.00mmol/L、4.00mmol/L、 6.00mmol/L、 8.00mmol/L、10.00mmol/L,反應(yīng)時(shí)間達(dá)到5分鐘,反應(yīng)后在340nm處測(cè)定吸光數(shù)值。
1.8.5 最佳加酶量確定
酶反應(yīng)體系300μL;DL-甘油醛為底物;NADPH作為體系輔酶;2%的乙腈為共溶劑;100mM/L PBS作為反應(yīng)的緩沖溶液;反應(yīng)溫度為37℃;PH控制在7.00;酶的用量分別為16ng、40ng、64ng、80ng、96ng幾個(gè)梯度,反應(yīng)時(shí)間達(dá)到5分鐘,反應(yīng)后在340nm處測(cè)定吸光數(shù)值。
1.8.6 最佳反應(yīng)底物確定
酶活反應(yīng)體系300uL;DL-甘油醛作為底物;NADPH作為體系輔酶;2%的乙腈作為共溶劑;100mM/L PBS作為反應(yīng)的緩沖溶液;反應(yīng)溫度為37℃;PH控制在7.0;底物分別為睪丸酮、孕酮、DL-甘油醛、雌二醇、甲睪酮、雌酮,反應(yīng)時(shí)間達(dá)到5分鐘,反應(yīng)后在340nm處測(cè)定吸光數(shù)值。
2 結(jié)果與分析
2.1 BL21-pET15b-AKR7A3表達(dá)菌構(gòu)建結(jié)果
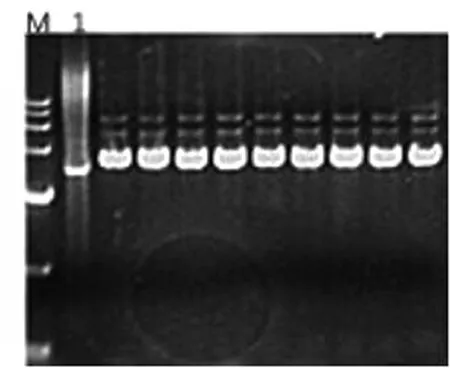
圖1 BL21-pET15b-AKR7A3質(zhì)粒鑒定
圖1中可以看出,M為15000marker,1泳道為空的pET-15b載體,其余條帶均比1泳道高。證明在氨芐青霉素平板上挑選的單菌落均成功轉(zhuǎn)入pET15b-AKR7A3質(zhì)粒,并說(shuō)明AKR7A3的表達(dá)載體已成功構(gòu)建,能用于后續(xù)實(shí)驗(yàn)。
2.2 AKR7A3蛋白表達(dá)結(jié)果
圖2中可以看出,M為15000marker,1泳道為BL21-pET15b-AKR7A3表達(dá)菌的上清液,其中目的蛋白醛酮還原酶AKR7A3大量表達(dá),其大小約為36kDa。4泳道為NPI-1流出液,其中無(wú)目的蛋白條帶,說(shuō)明醛酮還原酶AKR7A3與鎳柱充分結(jié)合。2,3泳道為洗脫后流出液,其中3泳道無(wú)雜蛋白,蛋白純度能夠達(dá)到99%。
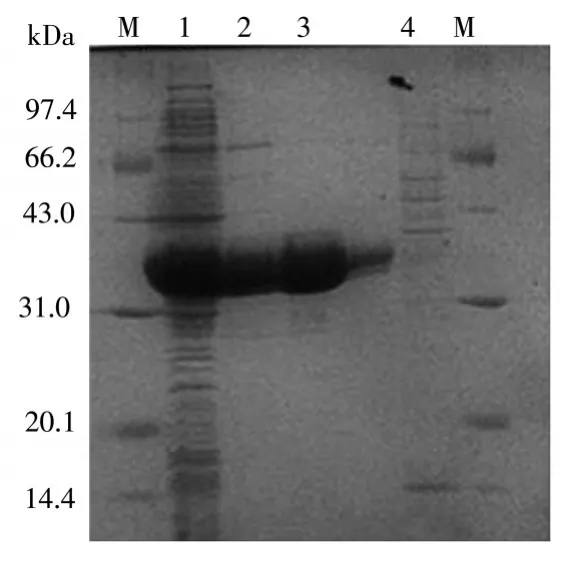
圖2 AKR7A3蛋白表達(dá)SDS-PAGE電泳圖
2.3 表達(dá)溫度優(yōu)化結(jié)果
在圖3中可以看出,醛酮還原酶AKR7A3在溫度為37℃、30℃、24℃、16℃環(huán)境下的表達(dá)情況。其中4泳道溫度為16℃,其上清中醛酮還原酶AKR7A3表達(dá)量是最多的。
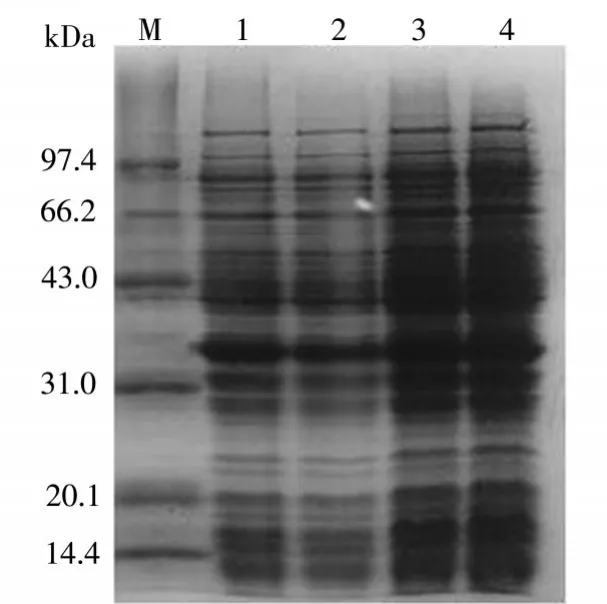
圖3 溫度優(yōu)化結(jié)果
2.4 表達(dá)轉(zhuǎn)速優(yōu)化結(jié)果
圖4中可以看出,醛酮還原酶AKR7A3在轉(zhuǎn)速分別為110rpm、130rpm、150rpm、180rpm條件下的表達(dá)情況。1泳道是轉(zhuǎn)速為110rpm,上清中的醛酮還原酶AKR7A3表達(dá)量最多。
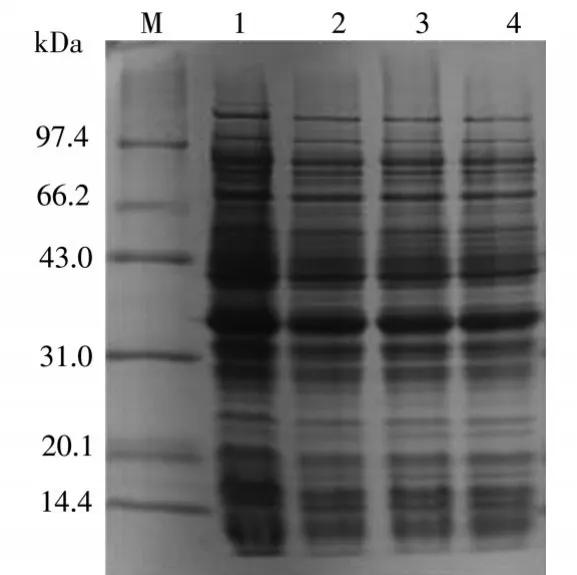
圖4 轉(zhuǎn)速優(yōu)化結(jié)果
2.5 表達(dá)時(shí)間優(yōu)化結(jié)果
在圖5可以看到,醛酮還原酶AKR7A3表達(dá)時(shí)間分別為5h、10h、16h、20h條件下的表達(dá)情況。其中4泳道為20h,上清中的醛酮還原酶AKR7A3表達(dá)量是最多的。
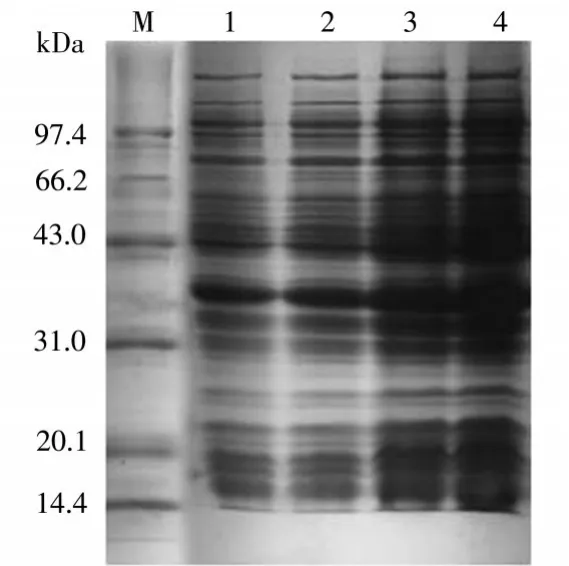
圖5 表達(dá)時(shí)間優(yōu)化結(jié)果
2.6 誘導(dǎo)劑IPTG濃度優(yōu)化結(jié)果
在圖6中可以看到,醛酮還原酶AKR7A3在誘導(dǎo)劑終濃度分別為0.40mM/L、0.60mM/L、0.80mM/L、1.00mM/L條件下的表達(dá)情況。4泳道誘導(dǎo)劑濃度為1.0mM/L,上清中的醛酮還原酶AKR7A3表達(dá)量是最多的。
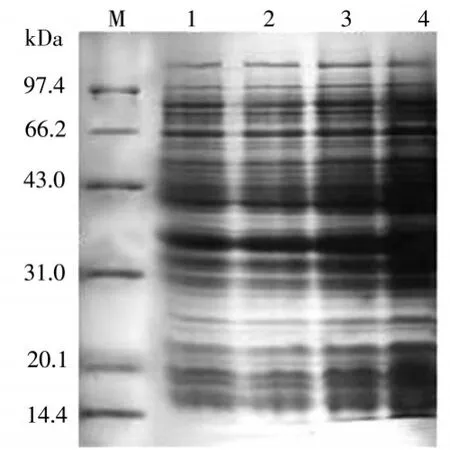
圖6 表達(dá)時(shí)間優(yōu)化結(jié)果
2.7 醛酮還原酶AKR7A3酶活性檢測(cè)
從溫度、PH、底物、NADPH、酶量等因素優(yōu)化來(lái)看,測(cè)得最佳動(dòng)力學(xué)參數(shù)如下:
在不同溫度(20℃、25℃、30℃、37℃、40℃)下檢測(cè)AKR7A3酶活力,結(jié)果表明在37℃時(shí)酶活力最高,如圖7(a)。在不同PH值(5.00、6.00、7.00、8.00、9.00)下檢測(cè)AKR7A3酶活力,結(jié)果表明在PH=7酶活力最高,如圖7(b)。不同NADPH濃度(0.10、0.15、0.20、0.30、0.40mmol/L)下檢測(cè)AKR7A3酶活力,結(jié)果表明在0.2mmol/L時(shí)酶活力最高,如圖7(c)。在底物不同濃度(2.00、4.00、6.00、8.00、10.00mmol/L)下檢測(cè)AKR7A3酶活力,結(jié)果表明在2.00mmol/L時(shí)酶活力最高,如圖7(d)。在不同加酶量(16.00ng、40.00ng、64.00ng、80.00ng、96.00ng)下檢測(cè)AKR7A3酶活力,結(jié)果表明在100μL時(shí)酶活力最高,如圖7(e)。在不同種類反應(yīng)底物下檢測(cè)酶活力,結(jié)果表明睪丸酮>雌酮>孕酮>甲睪酮>DL-甘油醛,但是在雌二醇為底物時(shí)醛酮還原酶檢測(cè)AKR7A3酶活力顯著小于其他底物,原因是由于雌二醇中沒(méi)有醛基/酮基,無(wú)法在醛酮還原酶的作用下進(jìn)行相關(guān)反應(yīng)。如圖7(f)。
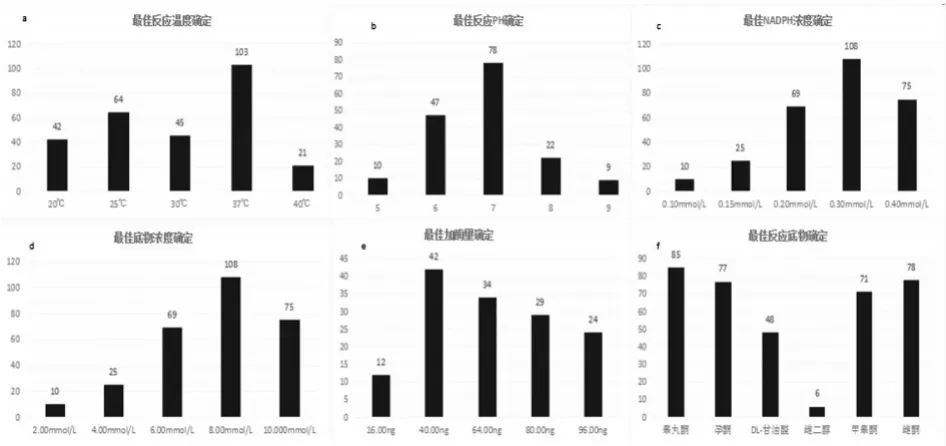
圖7 最佳酶活性確定
綜上所述,在 37℃;PH=7;NADPH 濃度0.2mmol/L;底物濃度2.00mmol/L;加酶量100μL;睪丸酮為底物時(shí)酶活力最高。
3 結(jié)論
本文為了能夠使得人源醛酮還原酶AKR7A3在該大腸桿菌BL21中正常表達(dá),在GEN BANK中查找人源AKR7A3基因序列,依照大腸桿菌表達(dá)的相關(guān)特征,對(duì)人源基因序列進(jìn)行密碼子的相應(yīng)優(yōu)化。合成優(yōu)化后的醛酮還原酶AKR7A3基因序列,構(gòu)建了AKR7A3-pET15b載體。使用大腸桿菌BL21作為宿主菌進(jìn)行表達(dá),并對(duì)AKR7A3表達(dá)的溫度、轉(zhuǎn)速、表達(dá)所需時(shí)間和誘導(dǎo)劑濃度等表達(dá)條件進(jìn)行優(yōu)化。實(shí)驗(yàn)得到最優(yōu)的表達(dá)條件為16℃、110rpm/h、20小時(shí)以及誘導(dǎo)劑濃度達(dá)到1.00mM/L。在優(yōu)化后BL21-pET15b-AKR7A3能夠大量表達(dá)可溶性好、活性高、純度達(dá)99.0%的醛酮還原酶AKR7A3。酶活性動(dòng)力學(xué)檢測(cè)結(jié)果表明,在37℃;PH=7;NADPH 濃度 0.2mmol/L;底物濃度2.00mmol/L;加酶量100μL;睪丸酮為底物時(shí)酶活力最高。
后續(xù)將在此基礎(chǔ)上,將通過(guò)基因重組大量制備AKR7A3酶蛋白,進(jìn)行酶激活劑及抑制劑的相關(guān)研究,從而篩選出對(duì)醛酮還原酶AKR7A3高效酶激活劑及抑制劑,為將來(lái)研究醛酮還原酶AKR7A3與相關(guān)癌癥發(fā)生與發(fā)展的基礎(chǔ)研究、以及及診斷與治療藥物研究提供實(shí)驗(yàn)基礎(chǔ)。
猜你喜歡
房地產(chǎn)導(dǎo)刊(2022年5期)2022-06-01 06:20:14
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
建材發(fā)展導(dǎo)向(2021年12期)2021-07-22 08:06:48
建材發(fā)展導(dǎo)向(2021年7期)2021-07-16 07:07:52
中學(xué)生數(shù)理化(高中版.高二數(shù)學(xué))(2021年12期)2021-04-26 07:43:48
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年12期)2021-03-08 01:28:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
