小黃黝魚繁殖期與非繁殖期的兩性異形
2018-10-29 08:04:38金錦錦張方方仇玉萍陳國柱
四川動物 2018年5期
金錦錦, 張方方, 仇玉萍, 陳國柱*
(1. 西南林業大學,云南省滇池濕地生態系統國家定位觀測研究站,昆明650224; 2. 西南林業大學,濕地學院/國家高原濕地研究中心,昆明650224)
兩性異形在動物界中普遍存在(Shine,1989;Morietal.,2017),研究證實其具有重要的進化生物學意義及生態適應意義(Estlanderetal.,2017)。對于入侵物種,兩性異形可能在促進其成功入侵過程中扮演著重要角色(Jeschke & Strayer,2006;樊曉麗等,2016;MacGregoretal.,2017;Milleretal.,2017;萬安等,2017)。入侵物種數量龐大,對其兩性異形進行過針對性研究的種類只占極少部分,極需深入系統地開展相關研究工作。
魚類普遍存在個體大小、體色和局部特征的兩性異形(Parker,1992;Pyron,1996)。部分成功入侵的魚類中存在兩性異形現象,如尼羅河尖吻鱸Latesniloticus(Dadeboetal.,2005)、黑口新蝦虎魚Neogobiusmelanostomus(Kornisetal.,2012)、麥穗魚Pseudorasboraparva(Záhorskáetal.,2013)、鲇Silurusasotus(樊曉麗等,2014)、食蚊魚Gambusiaaffinis(樊曉麗等,2016)、東部食蚊魚Gambusiaholbrooki(Sreanetal.,2017)等。這些魚類涵蓋了已知因性選擇及繁殖選擇壓力而形成的魚類個體大小兩性異形的3種表現形式(Parker,1992;林植華等,2007):雌魚大于雄魚,如尖吻鱸、食蚊魚,主要由提高雌魚繁殖力選擇壓力所致;雄魚大于雌魚,如麥穗魚,主要由雄魚護卵及爭奪雌魚選擇壓力所致;雌雄大小相似,如鲇魚,主要由提高受精率等選擇壓力所致。顯然,入侵魚類中復雜多樣的兩性異形形式提示它們適應環境壓力的多樣性較為豐富,針對性對某類入侵魚類兩性異形問題進行深入分析將有助于理解其入侵機制。
小黃黝魚Micropercopsswinhonis屬鱸形目Perciformes蝦虎魚亞目Gobiidae沙塘鱧科Odontobutidae,是廣泛分布于我國平原地區湖泊沿岸帶和河流緩流區的淡水小型魚類(易伯魯,1982),中國特有種(伍漢霖,鐘俊生,2008)。由于半個多世紀前的廣泛引種,小黃黝魚已經成為我國西南云貴高原(何記昌,劉振華,1985)及西北蒙新高原湖泊(郭焱等,2005)的典型入侵種,并逐步向青藏高原區域擴散(陳鋒,陳毅峰,2010;丁慧萍等,2014)。小黃黝魚種群數量優勢對入侵區域,如滇池(袁剛等,2010)、洱海(Tangetal.,2013)等高原湖泊的魚類群落和生態環境造成了一定影響;其入侵對云南香格里拉瀕危魚類中甸葉須魚Ptychobarbuschungtienensis是一個重要威脅(Panetal.,2010);其入侵與瀘沽湖中3種土著特有裂腹魚類的滅絕關系密切(孔德平等,2006)。因此,無論是基于對其種群控制還是基于對其資源利用,與繁殖相關的小黃黝魚對高原湖泊的入侵機制及其適應性均是亟待解決的重要科學問題。
過去人們針對入侵至朝鮮半島小黃黝魚的繁殖生物學及生態學進行了研究,如繁殖特性(Kim & Kim,1996)、卵子發育(Kim & Kim,1996;Kim,1996)、種群生態(Kim & Kim,1997)、胚胎和幼體發育特征(Iwataetal.,2001)、食物組成(Araoetal.,2010)等。盡管我國很早就意識到小黃黝魚入侵對高原區域土著魚類有重要影響,但迄今未進行過相關的入侵生態學研究。小黃黝魚存在兩性異形現象(易伯魯,1982),但僅限于形態、體色的描述而缺少定量分析(陳詠霞,管敏,2011),也缺少對這種現象與其入侵過程的相關分析。本文利用生態形態學分析方法(張堂林等,2008),觀察小黃黝魚繁殖期與非繁殖期形態特征的兩性異形,并分析其形成原因,為研究小黃黝魚對我國西南高原生境的入侵機制及其對高原湖泊的生態影響奠定基礎。
1 材料與方法
1.1 樣品采集地
樣品采集地位于云南省昆明市西郊團結鄉花紅園社區所在的山間盆地,該地是中華青鳉Oryziassinensis模式標本產地(陳銀瑞等,1989),同時也是小黃黝魚在滇池流域的一個典型入侵地。地形及濕地生境詳細描述參見仇玉萍等(2015)及陳國柱(2016),海拔約2 200 m。盆地西南側為三家村水庫(豐水期水面約1 km2),水庫壩下為低緩的丘陵盆地。水庫的湖濱帶、低洼耕地在棄荒后形成的山地沼澤、灌溉溝渠及排水渠道為小黃黝魚主要棲息生境。盆地出水口為地下溶洞,可能以潛流形式匯入滇池流域水系。
1.2 樣品采集與分析
2016年9月—2017年8月,逐月從樣品采集地用手抄網(孔徑為1.2 mm,網口30 cm)及地籠網(孔徑約3 mm)采集小黃黝魚,10%甲醛溶液現場固定備檢(1—2月枯水期、8月豐水期樣品不足,故不列入分析)。通過對雌魚的性腺觀察、卵巢稱量以及卵子計數,計算雌魚的性腺指數(gonadosomatic index,GSI),參考《魚類生態學》(殷名稱,1995)中的公式:GSI=100%×(性腺質量/體質量),雌魚GSI≥5%時為繁殖期(其卵巢發育至Ⅲ~Ⅳ期),其余為非繁殖期。雄魚直接判別,繁殖期雄魚頭部隆起,頭呈褐黑色、體色變黃,腹部赤紅。本研究共使用樣品253尾,雌魚135尾(其中120尾分析形態和性腺指數,另7尾僅分析形態,8尾僅分析性腺指數),雄魚118尾(其中3尾不作形態分析,只用于雌雄魚逐月體長對比分析),故用于形態分析的雌魚共127尾,雄魚115尾(繁殖期:雌魚50尾,雄魚56尾;非繁殖期:雌魚77尾,雄魚59尾),用于性腺分析的雌魚樣品為128尾,具體見各部分標注。繁殖世代間體長分布的比較分別使用其中的2017年3—5月混合樣品(n=30)與6月樣品(n=80)。
小黃黝魚傳統可量性狀指標測量方法主要參考《中國動物志 硬骨魚綱 鱸形目(五) 蝦虎魚亞目》(伍漢霖,鐘俊生,2008),使用電子數顯游標卡尺測量樣品的全長、體長、體高、頭長、吻長、尾柄長、尾柄高、眼徑、體寬等(精度為0.01 mm,計算平均值時四舍五入至小數點后第3位)(圖1),并用電子天平測量樣品的體質量、卵巢質量(精度為0.001 g),同時記錄性別。

圖1 小黃黝魚可量性狀及框架測量示意Fig. 1 Morphometric measurements of Micropercops swinhonis
A. 吻端, B. 第一背鰭起點, C. 第一背鰭基部末端, D. 第二背鰭起點, E. 第二背鰭基部末端, F. 尾鰭背部起點, G. 腹鰭起點, H. 臀鰭起點, I. 臀鰭基部末端, J. 尾鰭腹部起點, K. 胸鰭起點; A至B, A至C, ……, G至K之間的距離分別用AB, AC, ……, GK來表示; 1. 全長, 2. 體長, 3. 頭長, 4. 吻長, 5. 眼徑, 6. 體高, 7. 尾柄高, 8. 尾柄長, 9. 體寬
A. tip of snout, B. origin of first dorsal fin, C. tremunus of first dorsal fin, D. origin of second dorsal fin, E. tremunus of second dorsal fin, F. dorsal origin of caudal fin, G. origin of pelvic fin, H. origin of anal fin, I. tremunus of anal fin, J. ventral origin of caudal fin, K. origin of pectoral fin; AB, AC, …, GK indicate the distance of A to B, A to C, …, G to K; 1. total length, 2. standard body length, 3. head length, 4. lip length, 5. eye diameter, 6. body depth, 7. caudal peduncle depth, 8. caudal peduncle length, 9. body width
1.3 數據處理
所有數據均利用Excel和SPSS 19.0進行處理和分析。數值以平均數±標準差表示,顯著性水平設置為α=0.05。
為消除異速生長及體長差異對分析結果的影響,將除體長外的其他形態參數值與體長或頭長的比值作為統計值進行統計分析。利用單因素方差分析(One-Way ANOVA)對傳統形態性狀測量值及框架性狀測量值在雌雄小黃黝魚間的差異進行分析。基于上述傳統測量性狀及框架測量性狀共計33組可量性狀及比例參數,利用多元統計方法:主成分分析、判別分析等方法對雌雄魚的形態性狀差異進一步分析,以確認性別形態性狀差異。
2 結果
2.1 外部形態的兩性異形描述
GSI結果顯示,小黃黝魚3—9月繁殖,10月至次年2月停止繁殖(圖2)。體長分布顯示,小黃黝魚種群5—6月出現世代交替,可以劃分出5月前的世代Ⅰ(1+齡及以上)和6月后新成熟的世代Ⅱ(0+齡)2個繁殖群體(本文僅作指代,是未依據細分年齡劃分的2個群體,后者為前者春季繁殖仔魚當夏成熟所形成),2個群體的個體大小差異有統計學意義(圖3)。另一方面,小黃黝魚為分批產卵類型魚類,在繁殖季節中同時存在繁殖期個體及非繁殖期個體,本研究作兩性異形分析時,分別比較繁殖期與非繁殖期樣品。
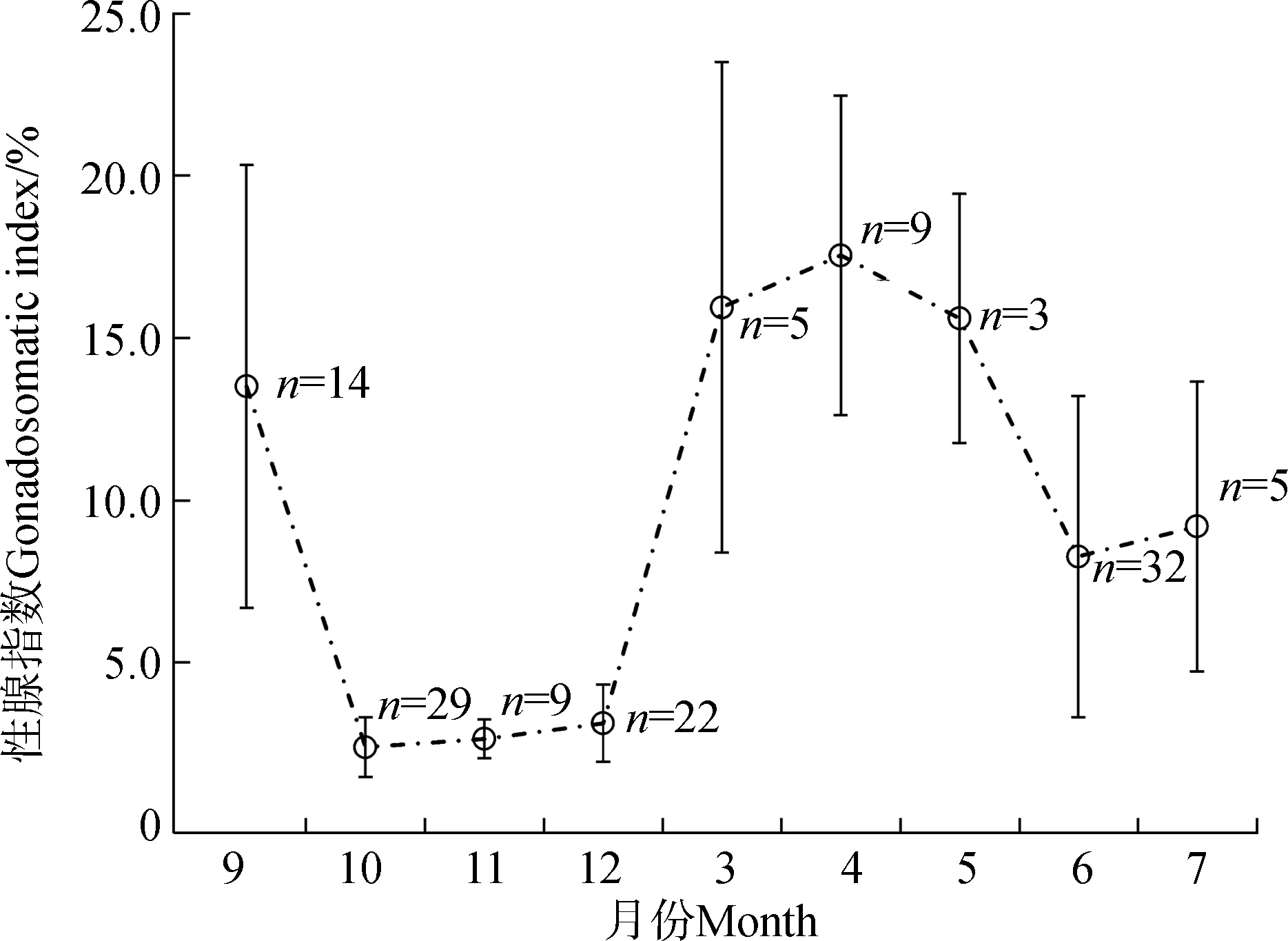
圖2 小黃黝魚研究期間雌魚性腺指數變化(n=128)Fig. 2 Changes of gonadosomatic index of female Micropercops swinhonis during the study period (n=128)
小黃黝魚雄魚在繁殖期與非繁殖期的外形特征存在差異,與雌魚的差異也十分明顯(表1,圖4)。其他特征差異與前人的研究(易伯魯,1982;陳詠霞,管敏,2011)一致。
小黃黝魚雄魚在繁殖期與非繁殖期的形態存在差異(圖4:a,b)。雄魚世代Ⅰ繁殖群體個體顯著大于世代Ⅱ繁殖群體(圖4:c)。雄魚能夠迅速從非繁殖期進入繁殖期,形態存在連續變化:非繁殖期頭部無隆起,曲線平滑;繁殖期初期,雄魚頭額部開始隆起;進入護卵期,頭額部、枕部隆起呈肉瘤狀(圖4:d)。
2.2 形態特征兩性異形單因素方差分析
雌雄小黃黝魚在多個形態參數差異有統計學意義。無論是世代Ⅰ還是世代Ⅱ繁殖群體,在繁殖期,雄魚體長均顯著大于雌魚(P<0.05)(表2,表3);而在非繁殖期,雄魚與雌魚個體大小差異無統計學意義(P>0.05)(表2,表3)。進一步分析顯示,雌雄個體大小差異在逐月分析的樣品中存在變化,以雄魚個體大于雌魚的月份居多(圖5)。
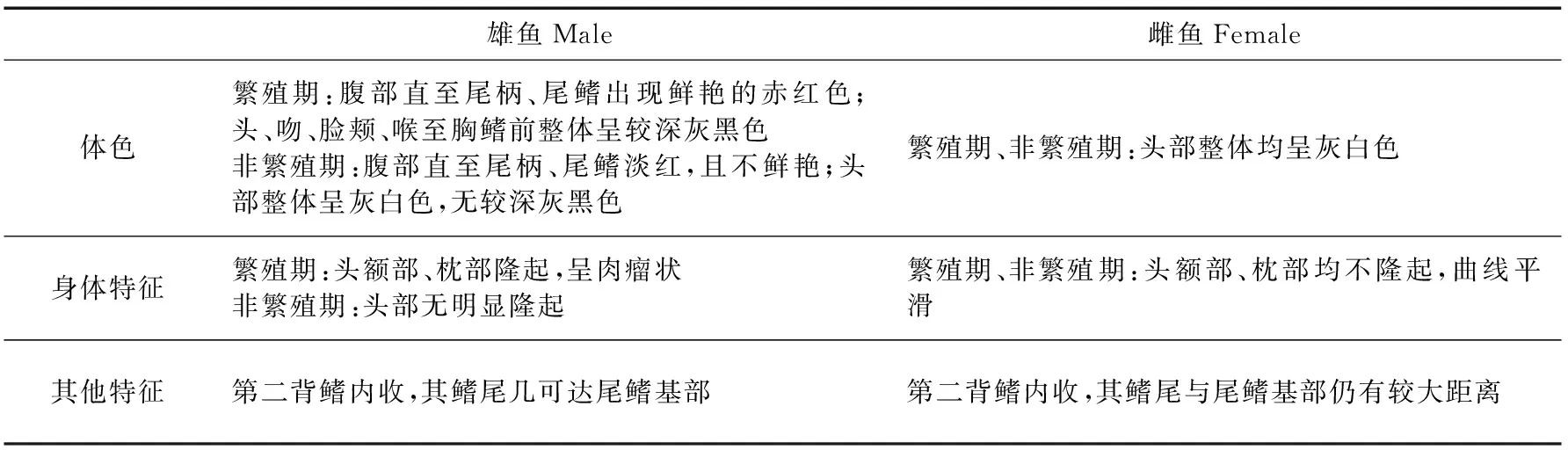
表1 雌雄小黃黝魚外形特征差異比較Table 1 Morphological characteristics between the male and female Micropercops swinhonis
2.3 形態特征兩性異形多元統計分析
世代Ⅰ繁殖群體:繁殖期共提取7個主成分,共解釋75.536%的變異,其中前5個主成分累計共解釋70.024%的變異:PC1=26.455%、PC2=17.762%、PC3=11.457%、PC4=8.553%、PC5=5.797%。非繁殖期共提取8個主成分,共解釋71.984%的變異,其中前4個主成分累計共解釋55.295%的變異:PC1=25.949%、PC2=14.086%、PC3=9.232%、PC4=6.028%。
世代Ⅱ繁殖群體:繁殖期共提取9個主成分,共解釋79.909%的變異,其中前5個主成分累計共解釋64.381%的變異:PC1=25.245%、PC2=17.550%、PC3=9.425%、PC4=6.793%、PC5=5.368%。非繁殖期共提取10個主成分,共解釋85.981%的變異,其中前4個主成分累計共解釋55.355%的變異:PC1=19.696%、PC2=14.157%、PC3=13.202%、PC4=8.300%。
以主成分PC1對主成分PC2繪制散點圖(圖6):2個繁殖群體中,繁殖期小黃黝魚雌雄之間空間位置明顯分離,非繁殖期雌雄魚高度重疊。在繁殖期,世代Ⅰ繁殖群體雌雄魚在PC2軸上顯著分離,尾柄高/體高(0.804)和腹鰭起點-臀鰭起點(GH)/體長(0.736)2個參數對主成分PC2有主要貢獻(>0.700),它們是影響兩者形態差異的主要因素;而世代Ⅱ繁殖群體在PC1軸上顯著分離,全長(0.707)、體長(0.755)、尾柄長/頭長(0.729)、尾柄高/體長(0.750)、頭寬/體長(0.814)、胸鰭起點-臀鰭起點(KH)/體長(0.764)6個參數對主成分PC1有主要貢獻(>0.700),它們是影響兩者形態差異的主要因素。
判別分析。世代Ⅰ繁殖群體:繁殖期雌雄間及其與非繁殖期雌雄間組質心距離均明顯;非繁殖期兩性組質心極為接近,兩者分值點存在相當大程度的重疊(圖7)。判別函數對原始案例正確分別率為79.4%,交叉驗證分類正確率為66.1%,其中,對繁殖期雄魚、雌魚,非繁殖期雄魚、雌魚等各類群的正確分別率分別為95.2%、87.1%、68.8%及78.5%。世代Ⅱ繁殖群體:與世代Ⅰ繁殖群體類似,繁殖期雌雄間及其與非繁殖期雌雄間組質心距離均明顯。同時,非繁殖期雌性和雄性組質心距離也較為明顯,兩者分值點重疊不明顯。判別函數對原始案例正確分別率為97.4%,交叉驗證分類正確率為66.2%,其中對繁殖期雄魚、雌魚,非繁殖期雄魚、雌魚的正確分別率分別為97.1%、94.7%、100.0%及100.0%。
3 討論
3.1 小黃黝魚兩性異形形成機制
動物兩性異形表現形式多樣,最重要的表現之一是個體大小異形(廖灝泓等,2013;Estlanderetal.,2017)。繁殖期小黃黝魚雄魚體長顯著大于雌魚,屬于雄魚大于雌魚的兩性異形表現形式。雄魚個體大于雌魚的現象在蝦虎魚類中較為普遍,如同色韌蝦虎Lentipesconcolor(Maciolek,1977)、雙點蝦虎魚Gobiusculusflavescens(Pélabonetal.,2003)、沙塘鱧Odontobutisobscurus(樊曉麗等,2009)等。魚類兩性個體大小異形一般解釋為性選擇(Legrand & Morse,2000)和繁殖力選擇(Szekelyetal.,2000)等綜合因素作用的結果。性選擇主要依賴于交配體制發生作用(Henson & Warner,1997),如雄性護卵、一夫多妻的交配體制,雄性為提高競爭力、交配成功率和對后代的保護,形成大個體(Snelson,1972;Parker,1992;林植華等,2007)。對黃顙魚Pelteobagrusfulvidraco

圖4 小黃黝魚外部形態Fig. 4 Morphology of Micropercops swinhonis
a. 非繁殖期(12月標本, 上: 雄魚, 下: 雌魚), b. 繁殖期(3月標本, 上: 雄魚, 下: 雌魚), c. 世代Ⅰ繁殖群體雄魚(下)與世代Ⅱ繁殖群體雄魚(上)對比, d. 雄魚從非繁殖期向繁殖期過渡的形態變化: 下. 非繁殖期, 中. 求偶及交配期, 上. 交配及護卵期
a. non-reproduction period (samples of December, upper: male, bottom: female), b. reproduction period (samples of March, upper: male, bottom: female), c. males from generation Ⅰ reproduction group (bottom) and generation Ⅱ reproduction group (upper), d. morphological changes of males from non-reproduction period (bottom), courtship period (middle), and mating and guarding period (upper)
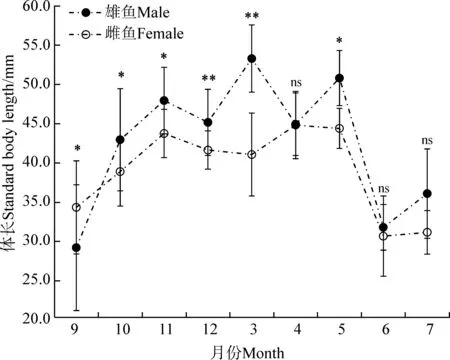
圖5 小黃黝魚雌雄逐月體長均值對比(n=253)
Fig. 5 Monthly changes of standard body length in the male and femaleMicropercopsswinhonis(n=253)
*P<0.05,**P<0.01, ns.P>0.05
(林植華,雷煥宗,2004)、棒花魚Abbottinarivularis(林植華等,2007)等的研究發現,雄性典型的筑巢、護巢和孵卵行為是促使雄性個體顯著大于雌性的重要影響因素。類似地,小黃黝魚在繁殖時,雄魚也具有典型的護巢行為。在護巢過程中,個體較大的雄性比小個體具有更強的護巢能力。小黃黝魚世代Ⅰ繁殖群體雄魚與雌魚形態出現差別的主要為尾柄高/體高、GH/體長等指標,且在雌雄間的差異有統計學意義,提示雄魚尾柄越大,運動能力和護巢能力越強,后代成活率就可能越高;而GH/體長與腹部容積增大有關,表明雌魚為獲得更大的懷卵量而演化出更大的腹腔。小黃黝魚世代Ⅱ繁殖群體雄魚與雌魚形態差別主要體現在與生長、繁殖有關的能量分配策略上。雄魚分配更多能量于生長以快速提升個體競爭力,而雌魚則分配更多能量于性腺發育,是導致兩者體長出現顯著差異的重要原因。2個世代兩性異形表現存在差異是由于兩者所在生長發育階段不同所致,同時也提示著兩性異形表現在小黃黝魚中具有較為豐富的可塑性。
繁殖力選擇更多出現在雌魚個體大于雄魚個體的物種中(Pincheira-Donoso & Hunt,2015),如尼羅河尖吻鱸(Dadeboetal.,2005)、食蚊魚(樊曉麗等,2016)等,雌魚通過身體的增大而增加繁殖輸出,是有利于提高種群增長率的一種繁殖策略。另一方面,因獲得的后代質量更高,雄魚傾向于選擇與個體較大的雌魚進行交配(Pélabonetal.,2003),也是促使雌魚個體在演化過程中向大個體發展以獲得更高繁殖力的重要原因。然而,繁殖力選擇壓力導致雌性大于雄性,但雄性小黃黝魚大于雌性的特點提示雄性護巢、競爭雌性能夠獲得更高的繁殖成功率,因此,性選擇壓力強于繁殖力選擇壓力是導致雄魚大于雌魚的重要原因。類似地,護巢及競爭配偶作用也被認為超過了繁殖力選擇壓力對雌雄棒花魚的形態塑造強度(林植華等,2007)。目前,偏向于雄性的兩性異形的物種中,雄魚護巢、爭奪配偶等策略是如何促進大個體雄性出現的細節有待闡明。
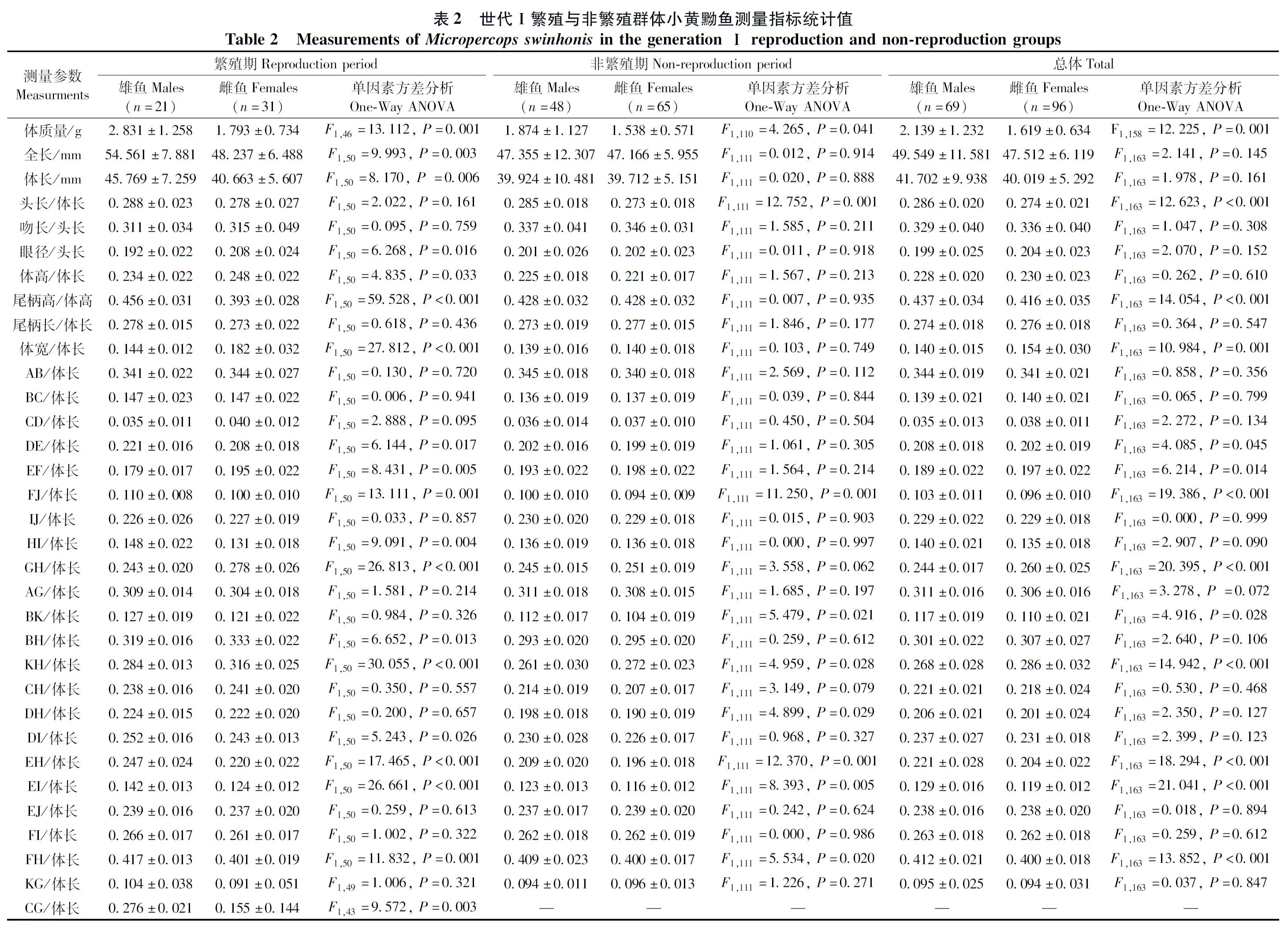
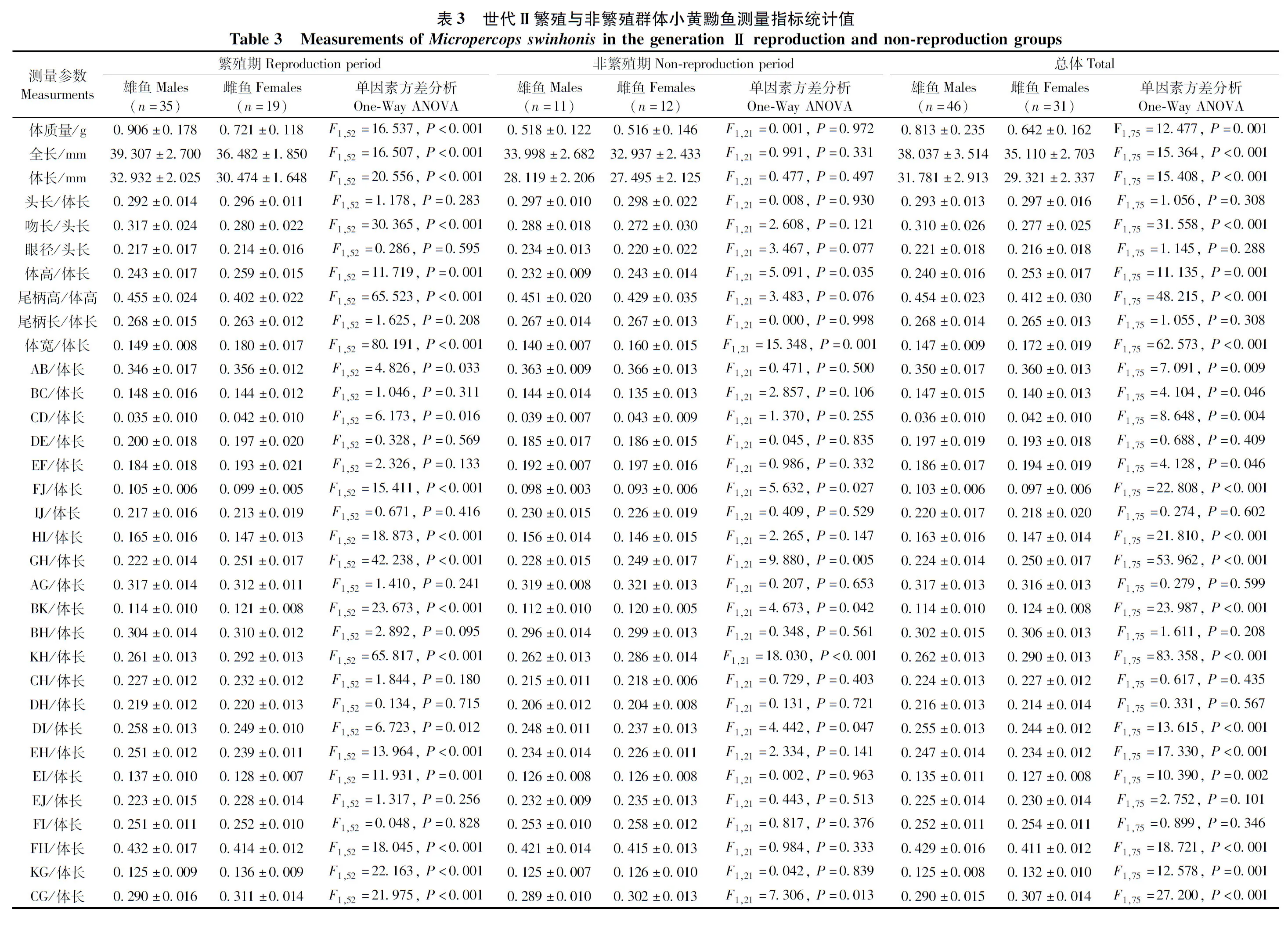
圖6 主成分PC1對PC2散點圖Fig. 6 Scatter plots for the principal component PC1 vs. PC2
a. 世代Ⅰ繁殖群體繁殖期, b. 世代Ⅰ繁殖群體非繁殖期, c. 世代Ⅱ繁殖群體繁殖期, d. 世代Ⅱ繁殖群體非繁殖期
Generation Ⅰ reproduction group: a. reproduction period, b. non-reproduction period; Generation Ⅱ reproduction group: c. reproduction period,d. non-reproduction period
3.2 小黃黝魚繁殖期與非繁殖期兩性異形表現差異

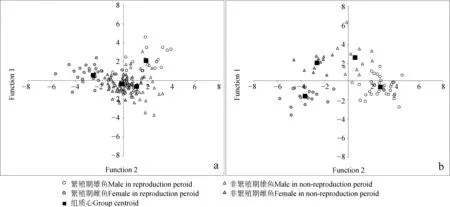
圖7 判別分析聯合分布Fig. 7 Combine plots for discriminant analysis
a. 世代Ⅰ繁殖群體generation Ⅰ reproduction group, b. 世代Ⅱ繁殖群體generation Ⅱ reproduction group
出兩性異形,如尼羅羅非魚Oreochromisniloticus(Dwivedietal.,2016)。在多數樣品采集月份中,小黃黝魚雄魚個體顯著或極顯著大于雌魚,這可能與兩者能量分配策略不同有關,雌魚將更多能量投入到性腺發育、繁殖輸出等活動中,而分配用于身體增長的能量比例較雄魚少,導致兩性體長增長產生差異而出現上述現象。
3.3 小黃黝魚兩性異形在其入侵中的作用
生態形態學理論(張堂林等,2008)指出,形態差異提示兩性生態位的差異。因此,小黃黝魚兩性異形現象可能在其入侵種群在入侵地成功建立種群過程中扮演著重要角色。雌雄形態上的差異對于降低兩者在種群內對某些資源如食物的競爭,特別是對于雌魚主要通過獲取更多食物以提高繁殖輸出的繁殖策略具有重要意義。而促進雄魚形態演化的護巢、競爭雌魚等行為能有效提高后代的質量與成活率,更有利于其種群擴張。類似現象在我國許多典型入侵種如黃顙魚(林植華,雷煥宗,2004)、棒花魚(林植華等,2007)、麥穗魚(Záhorskáetal.,2013)等研究中均有所揭示。盡管如此,這種貢獻對入侵種群在入侵初期快速增長過程中所起作用尚待深入分析。
