烏金豬、青峪豬和成華豬免疫器官與組織Toll樣受體 和抗菌肽基因表達比較研究
2018-10-29 08:11:02張敬梅顧以韌李江淩陳曉暉曾凱呂學斌高榮
四川動物 2018年5期
張敬梅, 顧以韌, 李江淩, 陳曉暉, 曾凱, 呂學斌*, 高榮
(1. 四川大學生命科學學院,生物資源與生態環境教育部重點實驗室,動物疫病防控和食品安全 四川省重點實驗室,成都610065; 2.四川省畜牧科學研究院,成都610066)
中國是世界上最大的豬肉生產國和消費國,隨著養豬業規模化與集約化程度的不斷提高,各種細菌和病毒傳染性疾病經常發生和流行,不僅嚴重危害我國養豬業的可持續發展和經濟效益,也威脅人民公共衛生和食品安全。豬Susscrofadomesticus的病原微生物種類多、變異快,導致疫苗和新藥研發成本高,防治效果受限。因此,培育抗病新豬種是目前解決豬病危害、提高養豬業經濟效益和保障公共衛生與食品安全經濟高效的新途徑。
Toll樣受體(toll-like receptors,TLRs)是1997年發現的介導天然免疫和獲得性免疫的病原模式識別受體家族,通過啟動天然免疫反應和激發適應性免疫反應的信號傳導,在宿主防御微生物病原體感染過程中發揮作用(Shaoetal.,2016)。TLRs屬于病原相關分子模式(pattern-associated molecular patterns,PAMPs)的識別受體,通過與病原的固有分子結合、激發信號轉導作用,誘導趨化因子和細胞因子的分泌,發揮先天免疫作用,并激活一系列適應性免疫反應(Kawai & Akira,2010;Zaidietal.,2016)。研究表明,TLRs是機體天然免疫系統的重要組成部分,在機體抗感染免疫中起著非常重要的作用(何小兵等,2011;Thakuretal.,2015)。TLRs能識別不同的PAMPs,包括脂多糖、雙鏈RNA病毒、脂蛋白、細菌CpG DNA及肽聚糖等(Leifer & Medvedev,2016;Luetal.,2018)。根據TLRs在細胞內的定位分為2個亞家族:細胞膜表面的TLR1、TLR2、TLR4、TLR5、TLR6、TLR11和細胞內膜的TLR3、TLR7、TLR8、TLR9(Yu & Feng,2018)。不同的TLRs識別不同的配體:細胞膜表面的TLRs主要識別病原的膜成分;TLR2與TLR1或TLR6形成異源二聚體,識別革蘭氏陽性菌的肽聚糖和磷壁酸;TLR4識別革蘭氏陰性菌表面的脂多糖;TLR5識別細菌的鞭毛蛋白;TLR11識別一些尿道細菌;細胞內膜的TLR3、TLR7、TLR8、TLR9主要識別病毒核酸成分(Hajishengallis & Lambris,2016)。養豬業常見的病毒性疾病包括豬瘟、口蹄疫、藍耳病等,這些病毒屬于單鏈RNA病毒。
抗菌肽是生物體內誘導產生的一種具有生物活性的小分子多肽,是抵御外界有害微生物或病毒入侵的重要屏障,也是生物體先天性免疫系統的重要組成部分(Andreu & Rivas,2015;Bechinger & Gorr,2017),在哺乳動物體內主要有防御素和Cathelicidin兩大抗菌肽家族(宴家友等,2013)。有關豬源抗菌肽的研究較全面:β-防御素1(poricine-β-defensin 1,pBD-1)是廣泛分布于呼吸道、消化道、胸腺、脾臟等組織內的一類活性多肽(江學斌等,2016),通過作用于病原體的細胞膜發揮抗菌作用,不會誘發細菌耐藥性,具有廣譜抗細菌、抗真菌及抗病毒活性,參與組成機體的先天性免疫系統,在抗感染中發揮重要作用(Aonoetal.,2006;Pruthvirajetal.,2016);PR-39是Cathecilidin家族中富含脯氨酸和精氨酸的小分子肽,具有較強的抗菌、抗病毒活性,對革蘭氏陰性菌和部分革蘭氏陽性菌具有很好抑菌效果,在動物機體先天免疫和抗感染免疫方面也發揮重要作用(Chenetal.,2015;盧順等,2015)。
成華豬是四川省著名的地方優良豬種,是四川推廣數量較多、覆蓋地域較廣的地方豬種,具有早熟易肥、肌纖維較短等優點,但也有生長周期長、養殖期耗料高等缺點。烏金豬具有耐粗飼、適應性強、肉質優良、抗逆性強及肌內脂肪豐富等優良性狀,但繁殖力低、生長慢、瘦肉率低。青峪豬作為四川省通江縣的地方豬種之一,具有耐粗性好、繁殖力強、適應性強、肉味香濃、口感細膩等優點。約克夏豬是我國引進的國外豬種,具有繁殖力高、生長快、瘦肉率高等優點,但抗病力和耐逆性較我國地方豬種弱(吳圣龍等,2008)。
雖有一些文獻報道了不同豬種TLRs、pBD-1和PR-39的表達水平(安沙等,2011;高彥華,2014;Holanietal.,2016),但目前尚無成華豬、烏金豬和青峪豬先天免疫分子特征的研究報道,因此,本實驗通過實時熒光定量PCR檢測TLRs(TLR1、TLR2、TLR4、TLR7、TLR9)、pBD-1和PR-39 mRNA在成華豬、烏金豬、青峪豬和約克夏豬的5種免疫器官和組織(胸腺、脾臟為系統免疫器官,扁桃體、腸系膜淋巴結為消化道局部免疫組織,肺門淋巴結為呼吸道局部免疫組織)中的表達差異,旨在揭示這些地方豬種之間TLRs和抗菌肽基因表達水平與約克夏豬的差異及特點,有助于從先天免疫分子層面深入闡明地方豬種抗病力較強的分子基礎,為今后利用地方豬種雜交培育抗病品系和篩選抗病分子標記提供科學依據和指導。
1 材料與方法
1.1 實驗動物與采樣
實驗用成華豬、烏金豬、青峪豬和約克夏豬來自四川省畜牧科學研究院簡陽種豬場,同組實驗豬均為同品種不同窩的母豬(非半同胞,父母不同),各組實驗豬采用相同營養水平和飼喂方式,飼養管理條件相同,8月齡時地方豬種體質量75~90 kg,約克夏豬125~135 kg。每組隨機選取5頭健康個體屠宰,取胸腺、脾臟、扁桃體、腸系膜淋巴結和肺門淋巴結樣品,無菌生理鹽水漂洗,迅速剪切后于液氮中速凍,-80 ℃保存。
1.2 總RNA的提取及cDNA的合成
RNA提取試劑用Trizol(TaKaRa),組織樣品100 mg,剪碎,加1 mL 1×磷酸緩沖鹽溶液(PBS)(成都奧克生物公司),使用TL2010S中通量組織研磨破碎儀(北京鼎昊源科技有限公司)研磨組織,12 000 r·min-14 ℃離心4 min;吸取上清0.2 mL,加入0.8 mL Trizol,劇烈震蕩,靜置10 min;加0.2 mL氯仿,劇烈振蕩15 s,室溫靜置10 min,12 000 r·min-14 ℃離心15 min;吸取上清液0.4 mL轉入新的EP管中,加入-20 ℃預冷15 min的異丙醇0.4 mL,混勻,室溫靜置10 min,12 000 r·min-14 ℃離心15 min;棄上清,加入1 mL 75%乙醇洗滌,7 500 r·min-14 ℃離心6 min,棄上清,干燥后加入30 μL無酶水溶解。用1%瓊脂糖凝膠電泳檢測總RNA的完整性,核酸分析儀NanoDrop2000(Thermo,美國)檢測總RNA的濃度和純度。第一鏈cDNA的合成參照反轉錄試劑盒TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix(TransGen Biotech,北京)說明書進行。
1.3 引物設計與合成
根據GenBank中報道的基因序列,使用Primer Premier 5.0設計實時熒光定量PCR引物,其中,肽基脯氨酰異構酶A(peptidylprolyl isomer-ase A,PPIA)為內參基因,引物序列及退火溫度見表1。

表1 實時熒光定量PCR引物序列Table 1 Primer sequences for real-time quantitative PCR
1.4 實時熒光定量PCR反應體系及反應條件
以PPIA為內參基因,在Bio-Rad IQ5實時熒光定量PCR儀(Bio-Rad,美國)上進行實時熒光定量PCR。擴增體系如下:3 μL模板cDNA,上、下游引物各0.5 μL,7.5 μL 2×SsoAdvanced Universal SYBR Green Supermix,加水補至15 μL。反應條件:95 ℃預變性30 s;95 ℃變性5 s,按表1中所列的退火溫度退火/延伸30 s,讀板收集熒光,共40個循環;最后從65 ℃開始以每秒0.5 ℃遞增至95 ℃進行熔解曲線特異性分析。每個樣品重復3次,同時設置無模板對照。
1.5 基因片段擴增效率的比較
以PCR產物為模板進行10倍濃度梯度稀釋(10-1~10-7),對所有引物進行實時定量PCR反應,輸入濃度梯度的數值,由Bio-Rad IQ5根據反應所得數據作出標準曲線,并得到相應的擴增效率(E)和可信度(R2)。
1.6 數據處理與分析
結果以PPIA為內參基因,以約克夏豬的組織作為對照組,用2-△△Ct法進行相對定量,數據用平均值±標準誤(Mean±SE)表示。數據使用Grapad Prism 6.01,各組織間的基因表達采用Two-Way ANOVA分析,并用Tukey法進行多重比較,顯著性水平設置為α=0.05。
2 結果
2.1 總RNA的純度與完整性
RNA純度檢測結果顯示,樣品的A260/280為 1.9~2.1,A260/230為2.0~2.5,說明RNA提取的質量較高。提取的各組織總RNA經1%瓊脂糖凝膠電泳,28S、18S和5.8S條帶均清晰可見,說明提取的RNA完整性較好,符合實驗要求。
2.2 實時熒光定量PCR引物特異性及擴增效率
從實時熒光定量PCR產物熔解曲線可以看出,各基因的熔解曲線均為單峰,無引物二聚體及非特異性產物形成。標準曲線分析結果表明,所有引物對經優化退火溫度后基因擴增效率均接近100%,可信度高(R2>0.98),表明該體系可用2-△△Ct法進行相對定量。
2.3 TLRs mRNA的表達比較
由圖1可知,成華豬胸腺中TLR7,脾臟中TLR4、TLR7,腸系膜淋巴結中TLR4、TLR9,肺門淋巴結中TLR1、TLR4、TLR7、TLR9 mRNA表達水平顯著高于其他豬種(P<0.05);烏金豬胸腺中TLR1、TLR9,扁桃體中TLR1、TLR2、TLR4、TLR7、TLR9和腸系膜淋巴結中TLR7 mRNA表達水平顯著高于其他豬種(P<0.05)。
2.4 pBD-1與PR-39 mRNA的表達比較
由圖2可知,成華豬胸腺中PR-39 mRNA表達水平顯著高于其他豬種(P<0.05);烏金豬扁桃體中pBD-1 mRNA表達水平顯著高于其他豬種(P<0.05);青峪豬扁桃體、腸系膜淋巴結中PR-39 mRNA表達水平顯著高于其他豬種(P<0.05);約克夏豬脾臟、肺門淋巴結中pBD-1和PR-39 mRNA表達水平顯著高于其他豬種(P<0.05)。
3 討論
先天性免疫系統是動物抵御病原入侵的第一道防線,PAMPs是先天性免疫的重要組成部分,在機體抵抗外界微生物入侵中起著重要的防御作用。TLR1、TLR2、TLR7、TLR9介導MyD88依賴性信號途徑;TLR4既可介導MyD88依賴途徑,也可介導TRIF依賴途徑。TLR7識別人工合成核酸的類似物以及來自單鏈病毒的ssRNA和siRNA,當宿主受到病原菌侵襲時,通過激活MyD88依賴的信號轉導通路產生Ⅰ型干擾素和炎癥細胞因子發揮抗病毒效應(Duetal.,2016;Petesetal.,2017);抗菌肽作為動物先天性免疫系統的重要防御物質,具有廣譜抗菌活性和抗腫瘤、抗病毒等生物活性。本文選擇了成華豬、烏金豬、青峪豬和約克夏豬的5種免疫器官和組織(胸腺、脾臟、扁桃體、腸系膜淋巴結、肺門淋巴結)進行了TLRs和抗菌肽基因mRNA的表達研究。
動物胸腺是免疫系統的中樞器官,在機體免疫,特別是在細胞免疫的演化、成熟和功能調節中起決定性作用,是參與免疫系統調控的主要內分泌腺;在T細胞的分化、發育、選擇和成熟過程中起關鍵作用,可促進T細胞發育成熟,表達不同的分化抗原,經胸腺選擇后獲得主要組織相容性復合體限制性(Sinkora & Butler,2009),維持機體正常的免疫平衡(龔非力,2000)。脾臟作為外周免疫器官,是機體最大的免疫器官,含有大量的淋巴細胞和巨噬細胞,是機體細胞免疫和體液免疫的中心(Cesta,2006)。
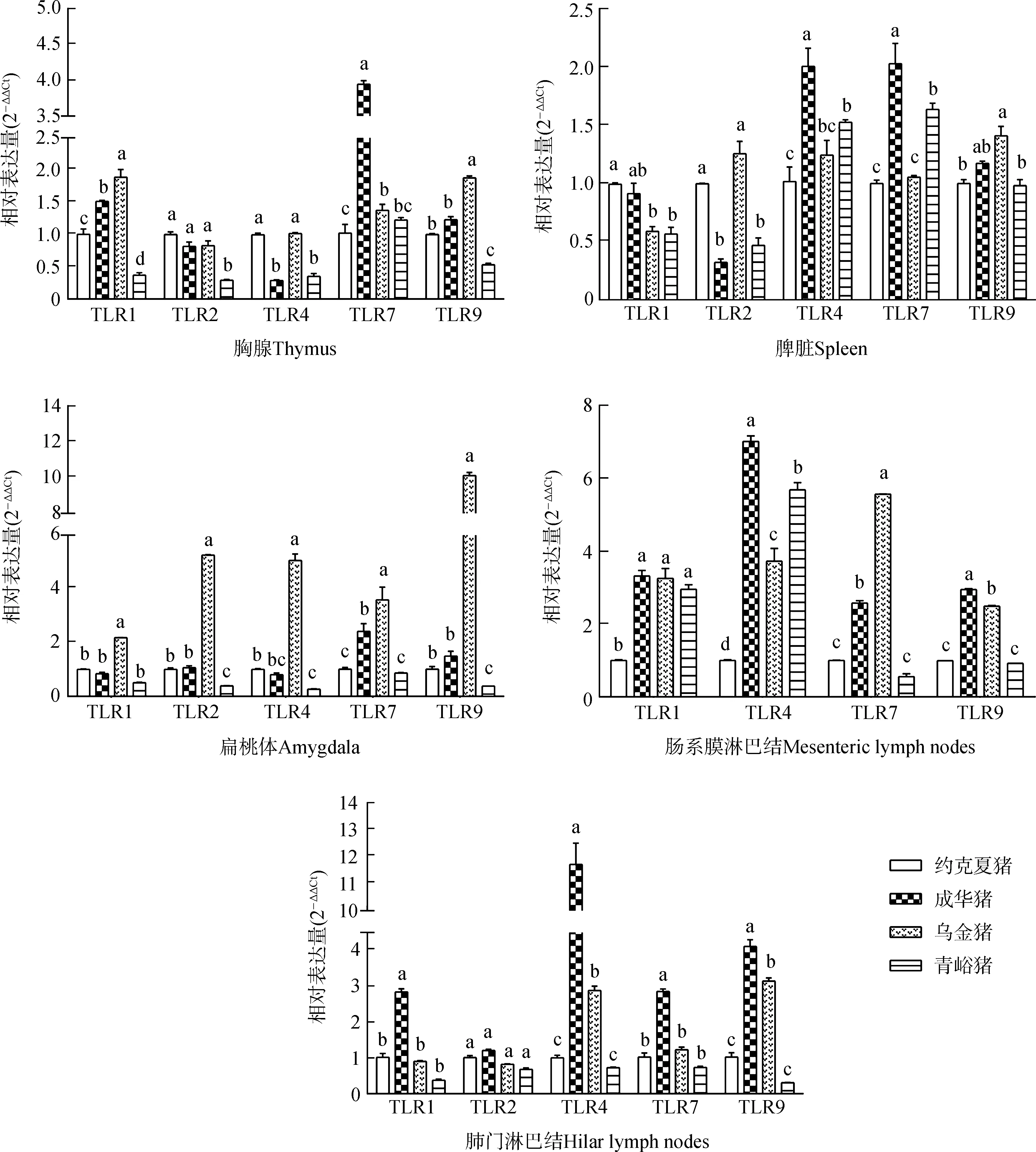
圖1 實驗豬免疫器官組織中TLRs mRNA的表達水平比較
Fig. 1 Comparison of the mRNA expression levels of TLRs in the immune organs and tissues of experimental pigs
不同字母表示組間數據的差異有統計學意義(P<0.05); 下圖同
Different letters indicate there is a significant difference between groups (P<0.05); the same below
本實驗發現,成華豬胸腺TLR7、PR-39 mRNA,脾臟TLR4、TLR7 mRNA表達水平較高,說明成華豬免疫器官的先天免疫水平明顯強于其他豬種;烏金豬胸腺中TLR1、TLR9 mRNA表達水平較高,表明烏金豬胸腺在先天免疫和抗感染免疫方面發揮重要作用。
扁桃體位于消化道和呼吸道入口的交會處,是機體第一道防線的重要組成部分,在抵御外界病原微生物的入侵中發揮著重要的作用。豬pBD-1 mRNA在舌和口腔黏膜高轉錄水平有利于黏膜和系統性防御(Qietal.,2009;Jiaoetal.,2017)。腸系膜淋巴結作為腸道相關淋巴組織中的一個重要組分,是機體免疫系統與外來抗原以及微生物較早接觸的免疫器官,同時也是較早發生免疫反應的器官(Teixeiraetal.,2013;張秀林等,2017)。PR-39 mRNA在腸系膜淋巴結中的表達差異可能與腸道內多種病原微生物的聚集有關。pBD-1不僅在模擬生理條件下抗菌效果明顯,而且與PR-39等Cathelicidin家族抗菌肽協同抗菌和抗病毒的作用非常顯著,在機體先天性免疫和獲得性免疫中發揮一定的免疫調節作用(Yangetal.,2001;Sangetal.,2006)。有報道表明,咽鼓管扁桃體表達TLRs(Lesmeisteretal.,2006),在天然免疫中發揮著重要作用(Manssonetal.,2006),同時在機體抵抗外界病原微生物的應答中也發揮著重要作用。烏金豬扁桃體中TLR1、TLR2、TLR4、TLR7、TLR9、pBD-1和腸系膜淋巴結中TLR7 mRNA表達水平均高于其他豬種,表明其消化道局部黏膜先天性免疫具有優勢。TLRs可識別病原微生物模式分子,參與誘導調控哺乳動物上皮細胞分泌抗菌肽殺死入侵機體的有害微生物(全佳慧等,2016)。成華豬腸系膜淋巴結中TLR4、TLR9 mRNA有較高的表達水平,說明成華豬腸系膜淋巴結通過PAMPs識別特定的病原微生物發揮特定的先天性免疫水平較高。青峪豬扁桃體和腸系膜淋巴結中PR-39 mRNA有較高的表達水平,表明其在抗菌及抗病毒等方面具有優勢。成華豬肺門淋巴結中TLR1、TLR4、TLR7、TLR9 mRNA有較高的表達水平,表明在成華豬肺門淋巴結中通過抗原遞呈,借助TLRs識別PAMPs,激活宿主的先天性免疫應答反應,抵抗病原微生物的感染(Ignacioetal.,2005;Holtetal.,2008;Kuzemtsevaetal.,2014)。
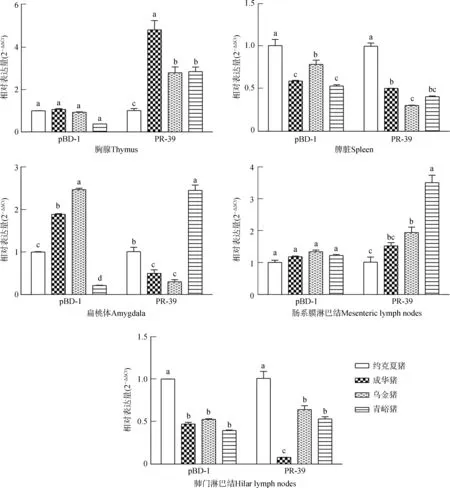
圖2 實驗豬不同免疫器官組織中pBD-1和PR-39 mRNA的表達水平比較Fig. 2 Comparison of the expression levels of pBD-1 and PR-39 mRNA in the immune organs and tissues of experimental pigs
呼吸道和消化道是各種致病微生物進入機體的主要通道,很多病原微生物由此侵害宿主,導致機體發病(Kirstil?etal.,1996)。上述結果表明,地方豬種相關局部免疫器官組織中TLRs高表達有助于機體發揮較強的先天性免疫反應,抵抗入侵的病原微生物,這為不同品種豬對于同一病原表現出不同抵抗力或敏感性提供了理論依據。
綜上所述,成華豬TLRs基因在系統免疫、消化道、呼吸道局部免疫組織以及PR-39基因在胸腺中高表達,表明成華豬在先天性免疫應答及抗病能力方面具有優勢。烏金豬TLRs基因在系統免疫器官、消化道局部免疫組織和pBD-1基因在扁桃體中高表達,表明其消化道有較強的先天免疫抗感染機能。本實驗揭示了地方豬種TLRs和抗菌肽基因在免疫器官組織中的表達水平及特點,可為利用地方豬種雜交選育抗病力強的新品種提供有益的線索和依據。
