麻醉-覺醒周期中非洲爪蟾的大腦復雜度研究
2018-10-29 08:04:10劉衍素范艷珠方光戰
四川動物 2018年5期
關鍵詞:實驗
劉衍素, 范艷珠, 方光戰*
(1.四川護理職業學院,成都610100; 2. 中國科學院成都生物研究所,成都610041)
全身麻醉是指藥物誘導下中樞神經系統被抑制的狀態,該狀態能保證長時間的手術操作和有創實驗研究的順利進行(Antogninietal.,2005;Goddard & Smith,2013)。全身麻醉以痛覺消失、靜止不動和意識喪失為特征,是麻醉藥品劑量和被麻醉對象意識狀態之間的平衡(Pocock & Richards,1993;Brownetal.,2010)。麻醉不充分可導致術中覺醒,而麻醉過量則增加并發癥風險(Zhangetal.,2001;Loepke & Soriano,2008)。因此,精確的麻醉深度監測對手術安全至關重要(Goddard & Smith,2013)。麻醉深度可用大腦活動的復雜度來刻畫(Bruhnetal.,2000;Zhangetal.,2001;Ferenetsetal.,2006;Liangetal.,2015;Hudetzetal.,2016):清醒時復雜度最高,而麻醉時復雜度最低。研究發現,不同腦區的大腦活動復雜度變化不盡相同;同時,與其他腦區相比,中腦網狀結構、丘腦、頂葉聯合皮層和額葉聯合皮層等特定區域對麻醉藥物更為敏感(Heinke & Koelsch,2005)。有研究認為,丘腦-皮層環路(特別是丘腦)可能在麻醉-覺醒調控中起著重要的調節作用(Ries & Puil,1999;Franks,2008),但是在麻醉-覺醒過程中,丘腦活動復雜度的動態變化特征依然不夠明確,同時尚不清楚丘腦是否為最重要的調控腦區。
大腦功能偏側性普遍存在于動物中(Rogers & Vallortigara,2008;Samara & Tsangaris,2011;Satoetal.,2011;Roussigneetal.,2012;Salvaetal.,2012;Frasnelli,2013;Rogersetal.,2013;Rogers,2014;Guoetal.,2016;薛飛等,2016)。當信息傳入大腦時,偏側性可確保大腦左右半球并行處理信息,從而提高信息處理能力(Vallortigara & Rogers,2005;Daddaetal.,2009;Fangetal.,2014)。可見,偏側性具有適應性意義,即通過提高動物感知信息和行為響應的效率來提高適合度。意識是大腦功能之一,所以麻醉-覺醒的調控(即意識的保持與恢復)可能亦存在偏側性,但需實驗證實。
腦電(electroencephalogram,EEG)是大量錐體細胞同步產生的突觸后電位總和在大腦皮層或頭皮表面的反映(Muthuswamyetal.,1996)。EEG是評估不同麻醉狀態下大腦活動情況最直接的方法,但是通過傳統的線性方法對原始EEG信號進行分析得到的結果往往難以解釋大腦在不同麻醉狀態下的各種特征(Billardetal.,1997;Katohetal.,1998;Bruhnetal.,2006;Mahonetal.,2008)。大腦通常表現出非線性和混沌行為(Bruhnetal.,2000;Buriokaetal.,2005),其特征可通過非線性參數加以描述;復雜度是眾多非線性參數中的一種,與事物的本質特征有關,而與事物的大小、多少等物理量沒有必然聯系。Lempel-Ziv復雜度(Lempel-Ziv complexity,LZC)是用來度量信號復雜程度,算法簡單快速且能有效刻畫腦電信號復雜度的物理量(Zhangetal.,2001;Abásoloetal.,2015),可用來研究大腦功能狀態及麻醉劑對中樞神經系統的抑制作用(Schartneretal.,2015)。
從宏觀解剖結構看,兩棲類動物的大腦與爬行類、鳥類和哺乳類(含人類)相似,均分為端腦、間腦、中腦、后腦和延髓等部分(Wilczynski & Endepols,2007),其間腦由背側的丘腦和腹側的下丘腦組成,且直接位于顱骨下方,而不像其他高等動物被大腦皮層覆蓋。這樣的結構有利于電極植入,更有利于采集到高信噪比的丘腦活動信號。非洲爪蟾Xenopuslaevis是兩棲類中應用最廣的動物模型,在發育、遺傳和神經系統功能等研究領域有著廣泛應用(Kay & Peng,1991;Schultz & Dawson,2003;Guénetteetal.,2013)。魚安定(triciane methanesulfonate,MS-222)是一種安全有效的水溶性麻醉劑,常用于魚類和蛙類等變溫動物的麻醉。
本研究以非洲爪蟾為對象,在其端腦、間腦、中腦左右兩側分別埋植電極,通過MS-222麻醉,連續記錄動物在“麻醉前清醒-給藥-恢復-麻醉后清醒”整個過程中的EEG信號。通過計算不同時期各腦區的LZC,探討麻醉-覺醒過程中大腦活動復雜度的動態特征,及可能存在的麻醉-覺醒調控偏側性。
1 實驗材料和方法
1.1 實驗動物
以14只成年非洲爪蟾(雌雄各半)為研究對象,按性別飼養于2個透明玻璃缸(長120 cm×寬50 cm×高60 cm)中,水深20 cm。每周投食、換水1次。養殖房室溫20 ℃±1 ℃,光周期12L∶12D(08∶00開燈)。手術時動物體長為8.1 cm±1.1 cm、體質量為67.1 g±22.2 g。所有手術及實驗流程均符合中國科學院成都生物研究所動物福利倫理委員會的相關要求。
1.2 動物手術
將動物浸入3.5 g·L-1MS-222溶液中進行麻醉,通過夾趾反應判斷麻醉程度,無夾趾反應時立即停止麻醉。局部消毒,去除手術區皮膚,剝離頭骨表面肌肉暴露頭骨。根據圖1所示坐標,分別在左右端腦(LT和RT)、左右間腦(LD和RD)、左右中腦(LM和RM)埋植不銹鋼電極(φ=0.8 mm),參考電極埋植在小腦上方(P)。LT和RT位于人字縫前方6.4 mm,旁開1 mm;LD和RD位于人字縫前方3.4 mm,旁開1 mm;LM和RM位于人字縫前方1.4 mm,旁開1 mm;P位于人字縫中線后1 mm處。電極擰入顱骨深度約1 mm,再用牙托水泥固定并覆蓋整個手術創口,用局部消炎止痛藥膏均勻涂抹創口。最后用自封膜包裹電極接插件以防水。術后,動物置于水深約10 cm的塑料盒中單獨飼養,休息1 d后進行數據采集。
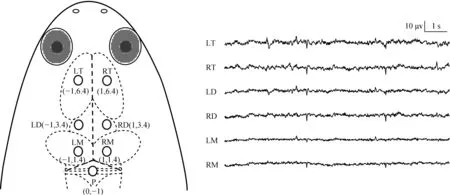
圖1 電極位置分布及相應的10 s腦電特征波形Fig. 1 Electrode placements and corresponding typical electroencephalogram tracings of 10 s
LT.左端腦, RT. 右端腦, LD. 左間腦, RD. 右間腦, LM. 左中腦, RM. 右中腦, P. 小腦上方; 下同
LT and RT. the left and right telencephalon, LD and RD. the left and right diencephalon, LM and RM. the left and right mesencephalon, P. above the cerebellum; the same below
1.3 數據采集
在電磁屏蔽的隔音室(背景噪音24.3 dB±0.7 dB)內進行實驗。將動物放在透明實驗盒(底面積18 cm×11 cm,開口處20 cm×13 cm,高12 cm)內吸滿水的海綿(厚度為1 cm、吸水量約150 mL)上,以保證動物皮膚濕潤,實驗盒上方加通氣透明的蓋子。光照和溫度與養殖房一致。在實驗盒上方約40 cm處安裝具有運動偵測功能的紅外攝像機,記錄動物運動行為。
實驗開始前,將動物連接至信號采集系統(RM6280,成都儀器廠),并設定采樣頻率為1 000 Hz,記錄動物麻醉-覺醒周期的EEG和行為數據:(1)麻醉前清醒期(Stage Ⅰ):記錄EEG 30 min,此時動物頭部通常朝向實驗盒一角并保持不動;(2)給藥期(Stage Ⅱ):從將動物換入裝有200 mL 3.5 g·L-1MS-222溶液的、與實驗盒同規格的塑料盒中進行麻醉開始,至無夾趾反應止;(3)恢復期(Stage Ⅲ):從將動物放入原記錄盒開始,至輕觸即出現自主運動止;(4)麻醉后清醒期(Stage Ⅳ):恢復期后連續記錄30 min。實驗結束后,通過過量麻醉對動物進行安樂死,并在電極的相應位點注射蘇木精染料,檢查電極位置是否與預期一致。
1.4 LZC
LZC是由Lempel和Ziv(1976)提出的一種時間序列復雜性測度分析方法,即先對時間序列進行符號化(粗粒化)處理后,通過計算其符號序列出現新模式的概率來表征時間序列的復雜度。計算步驟為:
(1)對時間序列進行符號化(粗粒化)處理:對于時間序列xt(t=1,2,3,…,n),根據選取的閾值(大于閾值時取1,否則取0),將其轉化成(0,1)符號序列,即St(t=1,2,3,…,n)。
(2)令St(t=1,2,3,…,n)和Qt(t=1,2,3,…,m)為2個由(0,1)序列組成的字符串;SQ表示S和Q 2個字符串的級聯,即SQ={S1,S2,…,Sn,Q1,Q2,…,Qm};SQπ表示把SQ中最后一個字符刪去后所得的字符串,即SQπ=(S1,S2,…,Sn,Q1,Q2,…,Qm-1}。令V(SQπ)表示SQπ中所有不同子串的集合,并設C(n)為符號序列St的復雜度。
(3)初始化:令C(n)=1,S={S1},Q={S2},則SQπ={S1}。
(5)重復步驟(4),直到待求序列中所有的字符被取完。這樣就把符號序列St(t=1,2,3,…,n)分成了C(n)個不同的子串,即得到復雜度。
由于對任意足夠長的序列,進行二進制符號化后所得的符號序列的復雜度趨向定值:
因此,復雜度可歸一化為:LZC=C(n)/b(n)。LZC值為0~1,反映時間序列的復雜程度。LZC值越小,說明時間序列的規律性越明顯;LZC值越大,則表示其復雜性越高,說明出現新模式的概率越高,即隨機性越強。
1.5 數據處理
EEG信號經50 Hz陷波、0.5~45 Hz帶通濾波和256 Hz降采樣后,以2 s為長度對數據進行分段。若數據最大幅度大于100 μV,即視為偽跡剔除;由于中值對奇異值不敏感,所以對無偽跡數據段,取其中值為閾值對數據進行符號化(Abásoloetal.,2015);然后計算每段的LZC值;最后每只動物按時段和腦區對LZC值進行平均。后續統計分析均基于上述均值。
1.6 統計分析
用Shapiro-WilkW檢驗和Levene’s檢驗對LZC值進行正態性及方差齊性檢驗,采用三因素(時段、腦區和性別)重復測量ANOVA進行統計分析,同時檢測主效應和交互效應。必要時使用Greehouse-Geisser校正;若存在交互效應則進行簡單效應分析。利用最小顯著性差異法(LSD)進行事后檢驗;效應度通過partialη2估計。利用Pearson相關分析計算各時段LZC值與給藥時長的相關性。所有統計在SPSS 21.0中完成,顯著性水平設置為α=0.05。
2 結果
在麻醉-覺醒過程中,不同腦區腦電LZC值的動態變化見圖2,在清醒階段(Stages Ⅰ/Ⅳ),LZC值最大;給藥時,LZC值急劇下降到最小;恢復時,LZC值逐漸上升。
2.1 LZC值的統計結果
ANOVA分析結果顯示(表1),時段[F(2, 24)=20.802,partialη2=0.634,P<0.001]和腦區[F(5, 60)=21.115,ε=0.547,partialη2=0.638,P<0.001]的主效應極顯著,而性別[F(1, 12)=4.342,partialη2=0.266,P=0.059]無主效應,且時段和腦區交互效應顯著[F(10, 120)=3.506,ε=0.394,partialη2=0.226,P=0.014]。
簡單效應分析顯示(表2),麻醉-覺醒周期中左端腦的LZC值最大,并且清醒階段(Stages Ⅰ/Ⅳ)端腦的LZC值極顯著大于間腦[Stage Ⅰ:F(5, 65)=36.252,ε=0.434,partialη2=0.736,P<0.001;Stage Ⅳ:F(5, 65)=8.668,partialη2=0.400,P<0.001],但在恢復階段,它們的差異無統計學意義。對于每個腦區而言,清醒階段的LZC值均極顯著大于恢復階段[左端腦:F(2, 26)=17.695,partialη2=0.576,P<0.001;右端腦:F(2, 26)=20.450,partialη2=0.611,P<0.001;左間腦:F(2, 26)=13.502,partialη2=0.509,P<0.001;右間腦:F(2, 26)=12.980,partialη2=0.500,P<0.001;左中腦:F(2, 26)=8.616,partialη2=0.399,P=0.001;右中腦:F(2, 26)=16.977,partialη2=0.566,P<0.001]。
2.2 LZC相關性分析結果
相關性分析結果顯示,在右端腦(r=0.665,P=0.009)和右間腦(r=0.625,P=0.017),Stage Ⅰ的LZC值與給藥時長顯著正相關;在右間腦(r=-0.542,P=0.045),Stage Ⅳ的LZC值與給藥時長顯著負相關(表3)。
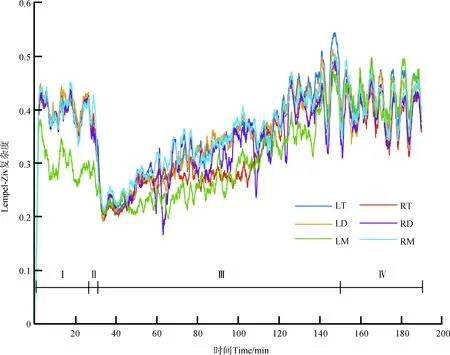
圖2 在麻醉-覺醒過程中非洲爪蟾不同腦區的腦電Lempel-Ziv復雜度Fig. 2 Lempel-Ziv complexity for different brain regions during the anesthesia-awakening cycle of Xenopus laevis
Ⅰ. 麻醉前清醒期, Ⅱ. 給藥期, Ⅲ. 恢復期, Ⅳ. 麻醉后清醒期; 下同
Ⅰ. Stage Ⅰ (pre-anesthesia stage), Ⅱ. Stage Ⅱ (administration stage), Ⅲ. StageⅢ (recovery stage), Ⅳ. Stage Ⅳ (post-anesthesia stage); the same below
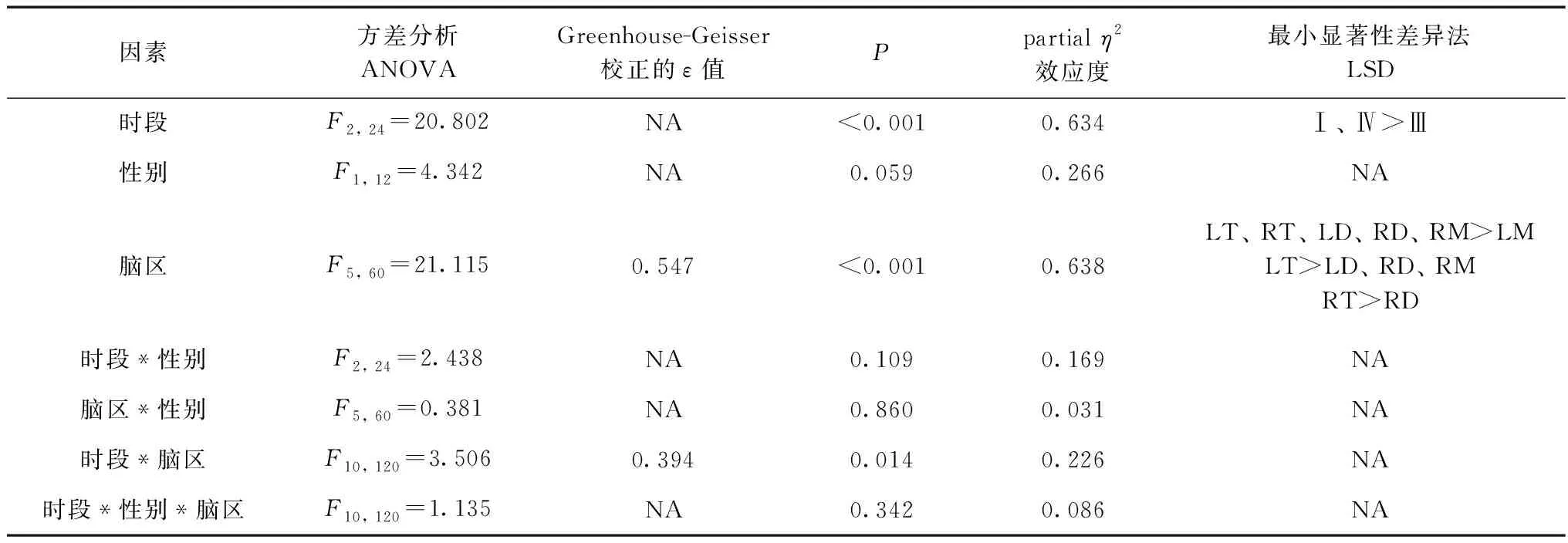
表1 Lempel-Ziv復雜度的ANOVA統計結果
注: >表示其左側相應條件下的LZC值大于右側, 同側內無差異; NA. 不適用; 下同
Notes: > denotes that Lempel-Ziv complexity values for the given conditions on the left side are significantly larger than those on the right side, and no significant difference exists among the corresponding conditions on the same side; NA. not applicable; the same below

表2 Lempel-Ziv復雜度的簡單效應分析Table 2 Simple effects analysis for Lempel-Ziv complexity
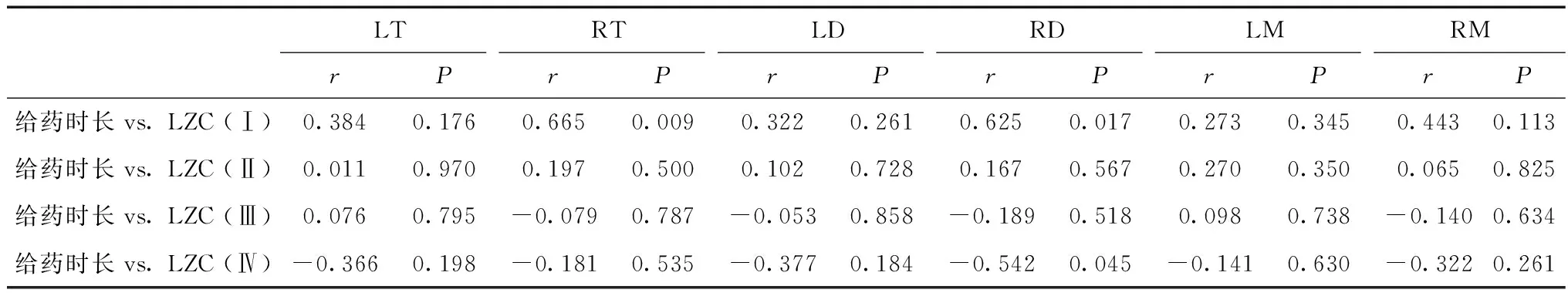
表3 Lempel-Ziv復雜度與給藥時長的相關性Table 3 Correlation analysis between Lempel-Ziv complexity and the duration of administration stage
3 討論
3.1 LZC隨麻醉-覺醒周期動態變化
通過計算非洲爪蟾“麻醉前清醒-給藥-恢復-麻醉后清醒”整個周期中EEG信號的LZC值,發現全身麻醉能引起其顯著改變:在麻醉前清醒期,LZC值處于較高水平;給藥時,LZC值驟降;當動物處于深度麻醉時,LZC值最小;恢復階段,LZC值逐漸恢復到原始清醒狀態時的水平。這種大腦復雜度隨著麻醉-覺醒周期動態變化的結果與非洲爪蟾在麻醉-覺醒過程中EEG信號的熵的動態變化相似(Fanetal.,2018),同時與人類病人在全身麻醉時大腦復雜度的動態變化類似(Bruhnetal.,2000),表明利用LZC值可以較好地刻畫麻醉-覺醒周期大腦的動態變化和測量麻醉深度(Zhangetal.,2001;Fanetal.,2011;Schartneretal.,2015;Hudetzetal.,2016)。統計結果顯示,清醒期(麻醉前和麻醉后)的LZC值顯著高于恢復期。LZC值越大,表明大腦系統復雜度越高,EEG信號更難預測;相反,LZC值越小,表明大腦系統復雜度越低,EEG信號變化具有更強的規律性和可預測性。清醒期LZC值的高水平與此時動物需不停采集外界信息并進行分析處理的功能需求一致(Heinke & Koelsch,2005);而在恢復期,大腦意識喪失,對外界刺激無響應,此時LZC值較低。由此可見,LZC值的變化與動物大腦功能狀態的改變相對應,說明此方法適用于麻醉深度和大腦活動復雜度的監測。
3.2 右側丘腦在麻醉-覺醒調控中可能起著關鍵作用
相關性分析顯示,右端腦和右間腦(丘腦)在麻醉前清醒期的LZC值與給藥時長正相關,說明右端腦和右側丘腦在麻醉前清醒期越活躍,進入全身麻醉狀態所需的時間就越長,這一結果與用近似熵和排列熵的分析結果類似(Fanetal.,2018)。另外,右側丘腦在麻醉后清醒期的LZC值與給藥時長呈負相關,說明給藥時長越短,右側丘腦在麻醉后清醒期就越活躍;反之,給藥時間越長,右側丘腦就越處于抑制狀態。由于LZC值和給藥時長的相關性主要出現在右側丘腦,而且和清醒期相比,恢復期丘腦與端腦之間的LZC值差異無統計學意義,說明右側丘腦在麻醉-覺醒調控中可能起著重要作用。這一推論與既往的研究結果一致:首先,脊椎動物不同腦區對麻醉劑的敏感程度不同(Heinke & Koelsch,2005);其次,大腦皮層、邊緣系統和網狀結構及皮層-丘腦環路在意識調控過程中起著關鍵作用(Heinke & Schwarzbauer,2002;Heinke & Koelsch,2005),而丘腦是多種感覺信息投射到端腦的中繼站(Sherman & Guillery,2002;Béhuretetal.,2013),全身麻醉導致的無意識狀態通常伴隨著丘腦代謝或血流減少,提示丘腦起著意識開關的作用(Alkireetal.,1997;Fisetetal.,1999)。當前結果進一步提示,右側丘腦在麻醉-覺醒調控中可能起著更為重要的作用,即這種調控具有偏側性,這一結論與包括人在內的所有脊椎動物的神經系統都具有結構和功能偏側性這一事實相符(Fangetal.,2014;Rogers,2014;薛飛等,2016;Vallortigara & Versace,2017)。總體上,端腦的LZC值最大,這與蛙類端腦具有高級認知和信息處理的功能相匹配(Fangetal.,2015;Xueetal.,2016a,2016b;Yueetal.,2017)。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
