芍藥苷對高糖刺激的小鼠骨髓來源的巨噬細胞TLR4信號通路的影響
2018-07-04 11:02:52楊雯雯段分分邵云俠吳永貴
安徽醫科大學學報 2018年7期
楊雯雯,段分分,邵云俠,王 坤, 吳永貴
隨著糖尿病發病率的上升,糖尿病腎病已成為全球終末期腎病的主要原因之一。從實驗和臨床研究中獲得的證據表明,腎臟炎癥在糖尿病期間腎臟損傷的進展中起關鍵作用[1]。Toll樣受體4(Toll-like receptor 4,TLR4)在糖尿病腎病中的作用機制已相當明確,Lin et al[2]發現在糖尿病腎病患者的腎活檢中TLR4表達升高,與之相關的腎組織中巨噬細胞的浸潤程度也隨之增加。白芍總苷(total glucosides of paeony, TGP)是我國重要傳統中藥白芍根的主要成分。前期研究[3-4]表明TGP可以抑制STZ誘導的糖尿病腎組織巨噬細胞激活,其機制部分與抑制腎組織中巨噬細胞的TLR4及STAT3信號通路有關。TLR4可以通過髓樣分化因子 88(myeloid differentiation primary response gene 88,MyD88)依賴途徑和MyD88非依賴途徑激活下游信號通路,參與核因子-κB(nuclear factor κB,NF-κB)的活化[5]。而芍藥苷(paeoniflorin, PF)作為TGP的主要有效成分,一直深受廣泛關注。該研究旨在探討PF是否通過TLR4信號通路抑制巨噬細胞的激活,為糖尿病腎病提供新的防治思路。
1 材料與方法
1.1材料6~8周齡雄性SPF級C57BL/6J小鼠和TLR4-/-小鼠(南京大學模式動物研究所);PF(南京廣潤公司);CCK-8檢測試劑盒(南京諾唯贊公司);兔抗TLR4、TIR結構域銜接蛋白(TIR-domain-containing adapter-inducing interferon-β,Trif)、MyD88、誘生型一氧化氮合酶(inducible nitric oxide synthase,iNOS)、腫瘤壞死因子α(tumor necrosis factor-α,TNF-α)、白細胞介素1β(interleukin-1β,IL-1β)、單核細胞趨化因子-1(monocyte chemoattractant protein 1,MCP-1)抗體(美國Abcam公司);兔抗磷酸化白介素-1受體相關激酶(phospho-IL-1 receptor-associated kinase-1,p-IRAK1)、磷酸化干擾素調節因子3(phospho-interferon regulatory factor 3,p-IRF3)、NF-κBp65、NF-κBp-p65抗體(美國Cell Signaling Technology公司);小鼠抗β-actin抗體、山羊抗兔IgG抗體、山羊抗小鼠IgG抗體(武漢三鷹公司);TNF-α、IL-1β和MCP-1 ELISA檢測試劑盒(美國R&D Systems公司);LeicaTCS SP5共聚焦掃描顯微鏡(德國Leica公司)。
1.2研究方法
1.2.1提取骨髓來源的巨噬細胞(bone marrow-derived macrophages,BMDMs)及鑒定 處死小鼠后,分離股骨和脛骨,在超凈工作臺中剪斷骨的兩端,用含有2%胎牛血清的無菌PBS溶液沖洗骨髓腔細胞,離心后棄去上清液,用配制好的DMEM培養基輕輕吹打重懸細胞,于37 ℃、5%CO2細胞培養箱中培養;7 d后收集細胞,用巨噬細胞表面抗原F4/80和CD11b抗體標記細胞,鑒定巨噬細胞的純度和成熟度。
1.2.2優化實驗條件 使用不用濃度的葡萄糖刺激BMDMs,同時不同濃度的甘露醇補充入各刺激組,以排除高糖產生的滲透壓對實驗的影響[5]。然后收集細胞,提取總蛋白,選取TLR4和iNOS蛋白表達最高時的糖濃度,作為實驗的刺激濃度;在確定刺激濃度后,選取不同刺激時間收集BMDMs,以確定高糖刺激的最佳時間。最后采用不同濃度的PF干預高糖刺激的BMDMs,選擇抑制蛋白表達上調的最適PF濃度。將BMDMs分為7組:正常糖對照組(LG組)、正常糖對照+PF組(LG+PF組)、高糖刺激組(HG組)、高糖刺激+PF組(HG+PF組)、TLR4-/-對照組(TLR4-/-組)、TLR4-/-對照+高糖刺激組(TLR4-/-+HG組)、TLR4-/-對照+高糖刺激+PF組(TLR4-/-+HG+PF組)。
1.2.3PF干預及高糖刺激對巨噬細胞活力的影響 將BMDMs制成細胞懸液接種于96孔板中,設無細胞組、無藥物對照組、空白對照組、不同濃度PF的高糖刺激組,24 h后每孔加入CCK-8溶液,用酶標儀進行光密度(optical density,OD)值測定,算出細胞活力。
1.2.4PF干預對高糖刺激的巨噬細胞遷徙能力的影響 將BMDMs接種于Transwell小室中,按照實驗分組,每組分別加入高糖刺激和PF干預,后在各組的Transwell下室中分別加入重組小鼠MCP-1培養后棄取上清液,Transwell上室乙醇固定20 min后,切去底部薄膜,結晶紫染色,輕輕鋪在載玻片上,顯微鏡下拍照后用PBS漂洗2次,薄膜置于33%冰醋酸中,回收結晶紫染色液,測定各組OD值后比較分析。
1.2.5實時熒光定量PCR法測定各分組細胞中的TNF-α、IL-1β、MCP-1及iNOS的mRNA的轉錄表達 提取每組BMDMs的總RNA后,立即檢測其純度,并反轉錄成cDNA。引物序列見表1。SYBR Green PCR 試劑盒被用來進行 PCR,基因的相對GAPDH表達采用 2-ΔΔCt法分析。

表1 各引物序列
1.2.6Western blot法檢測各組巨噬細胞的蛋白表達 提取各組細胞的總蛋白后,測定濃度。加樣后經電泳,轉膜,脫脂牛奶室溫封閉后,TBST洗膜3次,將膜放入一抗TLR4(1 ∶500)、MyD88(1 ∶100)、Trif(1 ∶1 000)、p-IRAK1(1 ∶200)、p-IRF3(1 ∶1 000)、IRF3(1 ∶1 000)、NF-κBp65(1 ∶1 000)、NF-κBp-p65(1 ∶1 000)、iNOS(1 ∶1 000)、β-actin(1 ∶35 000) 中4 ℃過夜,TBST洗膜3次,二抗室溫下孵育45 min后,再次洗膜3次,最后用ECL發光試劑盒進行顯影,Image J軟件對條帶進行分析。
1.2.7ELISA法檢測各組巨噬細胞的培養上清液中細胞因子的分泌量 實驗結束后收集各組培養上清液,離心,用ELISA試劑盒檢測各組BMDMs培養上清液中TNF-α、IL-1β 和MCP-1的含量。
1.2.8激光共聚焦顯微鏡分析各組巨噬細胞TLR4和iNOS的協同表達 細胞接種于Petri皿中,固定10 min后,PBS漂洗3次, 每次5 min, 5%驢血清封閉0.5 h,PBS再次漂洗3次,加入抗TLR4和抗iNOS抗體,4 ℃條件下孵育過夜,PBS漂洗3次,加入相應的熒光二抗,室溫下避光孵育2 h。再次PBS漂洗,DAPI染核10 min,PBS漂洗后進行封片,最后用激光共聚焦顯微鏡拍照。
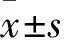
2 結果
2.1BMDMs純度及成熟度的鑒定流式細胞術鑒定結果顯示FITC及CD11b雙標的BMDMs達93.5%。見圖1。

圖1 檢測BMDMs純度及成熟度
2.2實驗條件的優化Western blot檢測顯示,高糖可導致TLR4表達上調,巨噬細胞活化;同時甘露醇作為高糖的滲透壓對照,對TLR4的表達及巨噬細胞的活化沒有影響。高糖濃度在30 mmol/L時,TLR4表達增加最明顯(0.55±0.01,F=379.480,P<0.01),iNOS表達也增加最明顯(0.37±0.01,F=283.612,P<0.01);高糖刺激的BMDMs在1 h TLR4開始過量表達,6 h開始巨噬細胞活化,產生iNOS, 與初始0 h比較:TLR4(1.1±0.002)和iNOS(0.82±0.02)均在24 h達到峰值(FTLR4=420.112,FiNOS=126.858,P<0.01);PF可以抑制TLR4表達及BMDMs的激活,并且抑制的程度與其劑量有關,在PF濃度為10-5mol/L時,TLR4(0.50±0.03)和iNOS(0.27±0.01)的表達抑制最為明顯(FTLR4=291.643,FiNOS=1818,P<0.01)。見圖2~4。
2.3PF干預及高糖刺激對巨噬細胞活力的影響高糖刺激對BMDMs活性無影響,PF濃度為10-3mol/L時對高糖刺激的BMDMs活性有影響(95.30±1.56,F=1.083,P<0.01)。見圖5。
2.4PF干預對高糖刺激巨噬細胞遷徙能力的影響高糖刺激可以增加MCP-1對巨噬細胞的趨化,導致巨噬細胞遷徙,而PF干預和TLR4基因敲除均可抑制高糖的刺激作用,各組間差異有統計學意義(F=49.39,P<0.01)。見圖6。

圖2 Western blot檢測高糖刺激BMDMs的TLR4及iNOS蛋白表達變化與正常糖濃度組(高糖5 mmol/L)比較:*P<0.05,**P<0.01

圖3 Western blot檢測高糖刺激BMDMs不同時間點TLR4及iNOS的蛋白表達變化與0 h比較:*P<0.05,**P<0.01

圖4 Western blot檢測不同濃度PF干預高糖刺激BMDMs對TLR4及iNOS的蛋白表達變化
A: LG組;B: HG組;C: HG+10-8mol/L組;D: HG+10-7mol/L組;E: HG+10-6mol/L組;F: HG+10-5mol/L組;G: HG+10-4mol/L組;與LG組比較:**P<0.01;與HG組比較:##P<0.01

圖5 PF對高糖刺激的BMDMs活性的影響
A: LG組;B: HG組;C: HG+10-8mol/L組;D: HG+10-7mol/L組;E: HG+10-6mol/L組;F: HG+10-5mol/L組;G: HG+10-4mol/L組;H: HG+10-3mol/L組;與HG組比較:*P<0.05
2.5實時熒光定量PCR檢測各組巨噬細胞的iNOS、TNF-α、IL-1β和MCP-1mRNA的表達與LG組比較,LG+PF組iNOS、TNF-α、IL-1β和MCP-1的mRNA的表達差異無統計學意義,HG組的mRNA轉錄表達明顯增加(P<0.01),正常糖濃度下敲除TLR4組的mRNA表達明顯減少(P<0.05,P<0.01);與HG組比較,PF組和TLR4-/-+HG組mRNA表達明顯下降(P<0.01);與TLR4-/-+HG組比較,敲除TLR4之后,PF仍然可以進一步抑制高糖導致的mRNA轉錄表達的增加(P<0.01),各指標組間差異有統計學意義(FiNOS=297.8、FTNF-α=162.8、FIL-1β=170.9、FMCP-1=102.2,P<0.01)。見圖7。
2.6Westernblot檢測各組巨噬細胞的iNOS、TLR4、MyD88、p-IRAK-1、Trif、p-IRF3、NF-κBp65和NF-κBp-p65蛋白表達與LG組比較,LG+PF組iNOS、TLR4、MyD88、p-IRAK-1、Trif、p-IRF3、NF-κBp65和NF-κBp-p65的蛋白表達差異無統計學意義,HG組各蛋白表達明顯上調(P<0.01),在正常糖濃度下,TLR4-/-組各蛋白的表達明顯減少;與HG組比較,PF干預組和TLR4-/-+HG組的蛋白相對表達量受到抑制(P<0.01);與TLR4-/-+HG組比較,TLR4-/-+HG+PF組在敲除TLR4基礎上,PF仍可以進一步抑制高糖導致的各蛋白表達增加(P<0.01)。各指標每組間差異有統計學意義(FTLR4=227.3、FiNOS=112.99、FMyD88=55.89、Fp-IRAK-1=16.75、FTrif=32.75、Fp-IRF3=42.11、FNF-κBp65=37.92、FNF-κBp-p65=343.9,P<0.01)。見圖8。
2.7ELISA檢測各組巨噬細胞分泌的TNF-α、IL-1β和MCP-1水平與LG組比較,LG+PF組細胞培養基上清中所含TNF-α、IL-1β 和MCP-1水平差異無統計學意義,HG組明顯增高(P<0.01),TLR4-/-明顯減少(P<0.05,P<0.01);與HG組比較,HG+PF組和HG+TLR4-/-組明顯減少(P<0.01);與HG+TLR4-/-組比較,敲除TLR4基因后,PF仍可進一步抑制高糖刺激導致的培養基中TNF-α、IL-1β 及MCP-1分泌增加(P<0.01)。每種指標各組間差異具有統計學意義(FTNF-α=29.76、FIL-1β=60.96、FMCP-1=41.65,P<0.01)。見圖9。

圖6 PF干預對高糖刺激巨噬細胞遷徙能力的影響 ×400
A: LG組;B: LG+PF組;C: HG組;D: HG+PF組;E: TLR4-/-組;F: TLR4-/-+HG組;G: TLR4-/-+HG+PF組;與LG組比較:**P<0.01;與HG組比較:##P<0.01;與TLR4-/-+HG組比較:ΔΔP<0.01

圖7 實時定量PCR檢測高糖刺激對各組巨噬細胞iNOS、TNF-α、IL-1β和MCP-1的mRNA表達
1: LG組;2: LG+PF組;3: HG組;4: HG+PF組;5: TLR4-/-組;6: TLR4-/-+HG組;7: TLR4-/-+HG+PF組;與LG組比較:**P<0.01;與HG組比較:##P<0.01;與TLR4-/-+HG組比較:ΔΔP<0.01

圖8 Western blot檢測各組巨噬細胞的蛋白表達
A:TLR4、iNOS、MyD88、p-IRAK1、Trif、p-IRF3蛋白相對表達量,B:NF-κBp65和NF-κBp-p65的蛋白相對表達;1:LG組;2: LG+PF組;3: HG組;4: HG+PF組;5: TLR4-/-組;6: TLR4-/-+HG組;7: TLR4-/-+HG+PF組;與LG組比較:**P<0.01;與HG組比較:##P<0.01;與TLR4-/-+HG組比較:ΔΔP<0.01

圖9 ELISA法檢測各組巨噬細胞培養基中分泌的TNF-α、IL-1β 和MCP-1表達
1: LG組;2: LG+PF組;3: HG組;4: HG+PF組;5: TLR4-/-組;6: TLR4-/-+HG組;7: TLR4-/-+HG+PF組;與LG組比較:*P<0.05,**P<0.01;與HG組比較:##P<0.01;與TLR4-/-+HG組比較:ΔΔP<0.01
2.8共聚焦顯微鏡分析各組巨噬細胞TLR4和iNOS的熒光表達與LG組細胞比較,LG+PF組綠色及紅色熒光表達無差別,TLR4-/-組紅、綠熒光基本無表達,HG組熒光表達增強;與HG組比較,HG+PF組、HG+TLR4-/-組、HG+TLR4-/-+PF組熒光表達降低,提示高糖可同時激活TLR4和iNOS在巨噬細胞胞質內表達,PF干預及TLR4基因敲除可抑制高糖的誘導作用。見圖10。
3 討論
糖尿病腎病與全身炎癥和局部的腎臟炎癥都有關,在糖尿病腎病的早期階段血清或外周血細胞中可檢測到炎性細胞因子(如TNF-α、IL-1及MCP-1)和免疫介質水平的升高,其中一些因子隨疾病進展而增加[6]。本實驗的刺激因素是高血糖,同時高血糖也是糖尿病腎病發生的先決條件,巨噬細胞是糖尿病腎臟中發現的最普遍的浸潤性白細胞,有研究[7-9]表明,高糖可以刺激巨噬細胞遷移,本實驗使用MCP-1作為初始趨化因子,在高糖的刺激下,各組實驗細胞均發生了趨化遷移,但與正常糖濃度對照組比較,高糖刺激組巨噬細胞遷移明顯增加,其增加的程度可被PF干預及TLR4信號通路敲除抑制。

圖10 共聚焦顯微鏡下各組巨噬細胞TLR4和iNOS的協同表達 ×400
Toll樣受體能識別危險相關分子模式的受體并促進先天性免疫反應,可通過兩種途徑激活下游信號分子:MyD88依賴途徑和MyD88非依賴途徑。這兩種途徑均可參與由TLR4激活的通路從而產生炎癥因子[10-11]。在未刺激的細胞中,NF-κB游離存在于胞質中;一旦受到刺激,NF-κB便移位于細胞核中并促進多種炎癥因子,如TNFα、IL-1β及MCP-1的釋放[12]。在本實驗中,高糖作為刺激因素,模擬了糖尿病的微環境,結果提示,在高糖刺激下,TLR4與高糖結合,作為內源性配體,激活下游的依賴MyD88信號途徑和非依賴MyD88信號途徑,使TLR4、MyD88、p-IRAK-1、Trif、p-IRF3、NF-κBp65及NF-κBp-p65表達均上調,與此同時啟動巨噬細胞的活化;表現為巨噬細胞活化產物iNOS,TNF-α、IL-1β及MCP-1的mRNA表達明顯上升,同時細胞的培養上清液中TNF-α、IL-1β及MCP-1水平也顯著升高。通過激光共聚焦顯微鏡中也可以看出高糖可以增加TLR4及iNOS的熒光強度,敲除TLR4基因后可以同時使iNOS熒光強度減弱,這提示TLR4及其下游的兩種信號通路可調節巨噬細胞激活。
PF作為TGP的主要生物活性成分,近幾年在臨床越來越多地被用來治療類風濕性關節炎、系統性紅斑狼瘡和系膜增生性腎炎等疾病[13-15],PF的藥理作用可能與抗炎、抗氧化和免疫調節有關,但是PF對腎臟保護作用的具體信號通路機制仍然是不明確的。本實驗室前期的研究[4]證實了TGP可抑制腎小球和腎小管-間質巨噬細胞TLR4的表達,在本實驗中,可以觀察到PF對體外高糖刺激的BMDMs激活起到抑制作用,可減少炎癥因子TNF-α、IL-1β、MCP-1及巨噬細胞表面活化標志物iNOS的產生;這些證實了PF具有抗炎作用;同時觀察到PF對正常糖濃度下的BMDMs及TLR4信號通路無影響,但是對高糖刺激下的BMDMs的TLR4及其下游信號通路有抑制作用,由此可以推測,PF是通過抑制高糖刺激下TLR4信號通路的上調而調節巨噬細胞活化的。另外,還可以觀察到,在敲除TLR4基因后,繼續給予PF干預,仍然可以進一步抑制BMDMs的激活,TNF-α、IL-1β、MCP-1及iNOS的產生也進一步減少,MyD88、p-IRAK-1、Trif、p-IRF3、NF-κBp65和NF-κBp-p65表達減弱,由此可以推測PF對巨噬細胞激活的抑制作用可能還與其他的信號通路有關,其具體的機制有待進一步研究。
綜上所述,高糖刺激可以導致巨噬細胞的TLR4信號通路激活,同時啟動巨噬細胞活化,PF可以通過抑制巨噬細胞TLR4信號通路的上調從而調節巨噬細胞的活化,但是PF的抑制作用是否同時與其他信號通路有關還需深入研究。
[1] Tesch G H. Diabetic nephropathy is this an immune disorder?[J].Clin Sci(Lond),2017,131(16):2183-99.
[2] Lin M, Yiu W H, Wu H J, et al. Toll-like receptor 4 promotes tubular inflammation in diabetic nephropathy[J]. J Am Soc Nephrol, 2012, 23(1):86-102.
[3] Wang K, Wu Y G, Su J, et al. Total glucosides of paeony regulates JAK2/STAT3 activation and macrophage proliferation in diabetic rat kidneys[J]. Am J Chi Med, 2012, 40(3):521-36.
[4] 趙 莉, 蘇雙全,夏 林. 白芍總苷對糖尿病大鼠腎組織TLR2、TLR4/NF-κB信號通路調節的影響[J]. 安徽醫科大學學報,2012, 47(5): 518-22.
[5] Cheng C I, Chen P H, Lin Y C. High glucose activates Raw2647 macrophages through RhoA kinase-mediated signaling pathway[J]. Cell Signal, 2015, 27(2):283-92.
[6] Perlman A S, Chevalier J M, Wilkinson P, et al. Serum inflammatory and immune mediators are elevated in early stage diabetic nephropathy[J]. Ann Clin Lab Sci, 2015, 45(3):256-63.
[7] Cheng C I, Chen P H, Lin Y C. High glucose activates Raw2647 macrophages through RhoA kinase-mediated signaling pathway[J]. Cell Signal, 2015, 27(2):283-92.
[8] Sato W, Kosugi T, Zhang L, et al. The pivotal role of VEGF on glomerular macrophage infiltration in advanced diabetic nephropathy[J]. Lab Invest, 2008, 88(9):949-61.
[9] Xu X, Qi X, Shao Y, et al. Blockade of TGF-β-activated kinase 1 prevents advanced glycation end products-induced inflammatory response in macrophages[J]. Cytokine,2016,78: 62-8.
[10] Akira S. Toll-like receptor signalling[J]. Nat Rev Immunol, 2004, 4(7):499-511.
[11] Medzhitov R. Toll-like receptors and innate immunity[J].Nat Rev Immunol, 2001, 1(2): 135-45.
[12] Mendes K L,Lelis D F.Nuclear sirtuins and inflammatory signaling pathways[J]. Cytokine Growth Factor Rev, 2017, 38: 98-105.
[13] Li P P, Liu D D, Liu Y J, et al. BAFF/BAFF-R involved in antibodies production of rats with collagen-induced arthritisviaPI3K-Akt-mTOR signaling and the regulation of paeoniflorin[J]. J Ethnopharmacol, 2012, 141(1):290-300.
[14] Wang Y N, Zhang Y, Wang Y, et al. The beneficial effect of total glucosides of paeony on psoriatic arthritis links to circulating Tregs and Th1 cell function[J]. Phytother Res, 2014, 28(3):372-81.
[15] Yun B R, Weon J B, Lee J, et al. Simultaneous determination of 11 bioactive compounds in Jaeumganghwa-tang by high performance liquid chromatographydiode array detection[J]. Pharmacogn Mag, 2014, 10(Suppl 2):S256-63.
猜你喜歡
中老年保健(2022年5期)2022-08-24 02:35:42
中老年保健(2022年1期)2022-08-17 06:14:56
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年5期)2021-08-24 07:07:20
中老年保健(2021年11期)2021-08-22 03:15:16
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
電子制作(2018年11期)2018-08-04 03:25:42
