脂氧素A4通過p38 MAPK及Nrf2通路調(diào)控氣道炎癥反應(yīng)
2018-06-21 13:19:38卓樂盈吳鎮(zhèn)杰于祥周美茜李成業(yè)歐陽(yáng)金生林琪斌蔡暢
溫州醫(yī)科大學(xué)學(xué)報(bào) 2018年6期
關(guān)鍵詞:水平
卓樂盈,吳鎮(zhèn)杰,于祥,周美茜,李成業(yè),歐陽(yáng)金生,林琪斌,蔡暢
(1.溫州醫(yī)科大學(xué) 第一臨床醫(yī)學(xué)院,浙江 溫州 325035;2.永嘉縣中醫(yī)院 呼吸科,浙江 溫州 325105;3.溫州醫(yī)科大學(xué)附屬第一醫(yī)院 呼吸科,浙江 溫州 325015)
氣道慢性炎癥是大多數(shù)呼吸道疾病的重要臨床病理生理特征。近年來(lái),隨著環(huán)境污染的逐漸加重,氣道慢性炎癥性疾病的發(fā)生率逐年增長(zhǎng)。人體氣道與外界環(huán)境相通,直接接觸各種氧化劑和毒性物質(zhì)、吸入性過敏原、病原微生物以及空氣污染物等,均可通過不同的機(jī)制激發(fā)炎癥反應(yīng)。
脂氧素(lipoxins,LXs)是一種重要的內(nèi)源性脂質(zhì)抗炎介質(zhì),能夠促進(jìn)炎癥反應(yīng)的及時(shí)消退,被譽(yù)為炎癥反應(yīng)的“剎車信號(hào)”或“停止信號(hào)”[1]。研究發(fā)現(xiàn)LXA4及其類似物在小鼠哮喘模型中具有抑制炎癥及促進(jìn)炎癥消退的作用,包括增強(qiáng)自然殺傷細(xì)胞功能,刺激巨噬細(xì)胞表達(dá)CD206[2],還可以促進(jìn)肺損傷的修復(fù)[3]。但目前LXs在氣道炎癥進(jìn)程中參與的信號(hào)通路及其分子調(diào)控機(jī)制尚未完全清楚。本研究通過脂多糖(lipopolysaccharide,LPS)刺激BEAS-2B細(xì)胞,探討LXA4對(duì)氣道炎癥反應(yīng)的抑制作用及其所涉及的信號(hào)通路。
1 材料和方法
1.1 材料 BEAS-2B細(xì)胞株購(gòu)自中國(guó)醫(yī)學(xué)科學(xué)院昆明細(xì)胞庫(kù)。LPS、Trizol和DCFH-DA試劑購(gòu)于美國(guó)Sigma公司;LXA4購(gòu)自美國(guó)Cayman Chemical公司;BEGM培養(yǎng)基購(gòu)自美國(guó)Lonza公司;RPMI 1640培養(yǎng)基、胎牛血清購(gòu)自美國(guó)Gibco公司;谷胱甘肽(Glutathione,GSH)檢測(cè)試劑盒購(gòu)于南京建成生物工程研究所;BCA試劑盒、細(xì)胞漿和細(xì)胞核蛋白抽提試劑盒購(gòu)自上海碧云天生物技術(shù)有限公司;人p38、p-p38抗體購(gòu)自美國(guó)Cell Signal Technology公司;人核因子E2相關(guān)因子(nuclear factor erythroidderived 2-like 2,Nrf2)、p-Nrf2、核纖層蛋白B1(Lamin B1)抗體購(gòu)自美國(guó)Abcam公司;人β-tubulin抗體購(gòu)自杭州聯(lián)科生物公司;SYBR Green realtime PCR Master Mix、反轉(zhuǎn)錄-聚合酶鏈反應(yīng)試劑盒購(gòu)自日本Takara公司;引物由上海生工生物工程有限公司合成。
1.2 方法
1.2.1 試劑配制:LPS配成貯存濃度為100μg/mL 的溶液,分裝后-20℃保存;為保持試劑穩(wěn)定性,LXA4用無(wú)菌無(wú)水乙醇溶解,配成貯存濃度為100 μmol/L 的溶液,分裝后-80℃保存?zhèn)溆茫褂脮r(shí)取適量溶液,待無(wú)水乙醇蒸發(fā)完畢后,加入培養(yǎng)基重新溶解。
1.2.2 BEAS-2B細(xì)胞培養(yǎng):BEAS-2B細(xì)胞培養(yǎng)于BEGM培養(yǎng)基,按1×105個(gè)細(xì)胞/mL密度接種到培養(yǎng)瓶,置于37℃,5% CO2培養(yǎng)箱中培養(yǎng),每2~3 d換液1次,BEAS-2B細(xì)胞為貼壁生長(zhǎng),細(xì)胞匯集至70%~80%時(shí),按1:3傳代。
1.2.3 實(shí)驗(yàn)分組及干預(yù):將對(duì)數(shù)期生長(zhǎng)的BEAS-2B細(xì)胞分為3組。對(duì)照組:不做任何處理;LPS組:100 ng/mL LPS刺激24h;LPS+LXA4組:100 nmol/L LXA4預(yù)處理30min,加入100 ng/mL LPS刺激24h。收集細(xì)胞或培養(yǎng)上清進(jìn)行后續(xù)實(shí)驗(yàn)。
1.2.4 實(shí)時(shí)熒光定量PCR:以Trizol法提取各組細(xì)胞總RNA,并測(cè)定RNA濃度及純度(2.0>A260/A280>1.8)。按照反轉(zhuǎn)錄試劑盒操作說明將RNA反轉(zhuǎn)錄為cDNA。取cDNA產(chǎn)物用SYBR Green real-time PCR Master Mix進(jìn)行實(shí)時(shí)熒光定量PCR,每個(gè)樣本設(shè)3個(gè)復(fù)孔。采用2-ΔΔCt法進(jìn)行數(shù)據(jù)處理,計(jì)算實(shí)驗(yàn)組相對(duì)于對(duì)照組的基因表達(dá)倍數(shù)變化。IL-1β上游引物序列:5’-CCAGGAGAATGACCTGAGCA-3’,下游引物序列:5’-GGAGCGTGCAGTTCAGTGAT-3’;IL-6上游引物序列:5’-TGAGGAGACTTGCCTGGTGA-3’,下游引物序列:5’-TGCAGGAACTGGATCAGGAC-3’;血紅素加氧酶-1(heme oxygenase,HO-1)上游引物序列:5’-CAGTGCCACCAAGTTCAAGC-3’,下游引物序列:5’-CTGGATGTTGAGCAGGAACG-3’;醌氧化還原酶[NAD(P)H:quinone oxidoreductase,NQO-1]上游引物序列:5’-AAGCCGCAGACCTTGTGATA-3’,下游引物序列:5’-TGGCAGCGTAAGTGTAAGCA-3’。GAPDH直接從上海生工生物工程公司購(gòu)買。
1.2.5 流式細(xì)胞術(shù):收集各組細(xì)胞制備單細(xì)胞懸液[細(xì)胞數(shù)量為(5~6)×106],用PBS清洗2~3次,用0.4mL含1% FBS的PBS重懸細(xì)胞,分別加入流式管。各組加入DCFH-DA,37℃孵育20~30min。PBS清洗2~3次,加入0.3mL含1% FBS的PBS重懸細(xì)胞,注意避光。用流式細(xì)胞儀上機(jī)檢測(cè),F(xiàn)lowJo.10軟件分析實(shí)驗(yàn)數(shù)據(jù)。
1.2.6 GSH含量檢測(cè):收集各組細(xì)胞[細(xì)胞數(shù)量為(5~6)×106],用PBS作為勻漿介質(zhì)破碎后離心取上清,檢測(cè)細(xì)胞內(nèi)GSH表達(dá)水平,測(cè)定方法根據(jù)南京建成生物工程研究所試劑盒說明書操作。各組設(shè)置3個(gè)復(fù)孔以減少誤差。
1.2.7 Western blot:BCA法測(cè)定蛋白濃度。抗體濃度分別為:兔抗p38抗體、兔抗p-p38抗體、兔抗Lamin B1抗體、兔抗Nrf2抗體和兔抗p-Nrf2抗體 1:1 000;鼠抗β-tubulin抗體1:4 000,HRP標(biāo)記的羊抗兔IgG抗體、羊抗鼠IgG抗體1:8 000。用Bio-Rad Image Lab軟件分析條帶光密度,半定量比值測(cè)定分析。
1.3 統(tǒng)計(jì)學(xué)處理方法 應(yīng)用SPSS22.0統(tǒng)計(jì)學(xué)軟件進(jìn)行統(tǒng)計(jì)分析。計(jì)量資料以±s表示,用Shapiro-Wilk法行正態(tài)性檢驗(yàn),用Levene法行方差齊性檢驗(yàn),各組間比較采用單因素方差分析,組間兩兩比較用LSD法檢驗(yàn),方差不齊采用秩和檢驗(yàn)。P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 LXA4對(duì)LPS誘導(dǎo)BEAS-2B細(xì)胞產(chǎn)生IL-6和IL-1β的影響 與對(duì)照組比,LPS組BEAS-2B細(xì)胞IL-6和IL-1β mRNA水平明顯升高(P<0.05)。與LPS組比,LXA4+LPS組BEAS-2B細(xì)胞IL-6和IL-1β mRNA水平顯著降低(P<0.05),見圖1。
2.2 LXA4對(duì)LPS誘導(dǎo)BEAS-2B細(xì)胞氧化應(yīng)激的影響 炎癥反應(yīng)的發(fā)生發(fā)展常伴隨著大量氧自由基的產(chǎn)生。在LPS的刺激下,BEAS-2B細(xì)胞內(nèi)ROS水平顯著上調(diào),差異有統(tǒng)計(jì)學(xué)意義(P<0.05)。LXA4預(yù)處理后,ROS水平明顯下降,而GSH水平明顯升高,差異均有統(tǒng)計(jì)學(xué)意義(P<0.05),見圖2。
2.3 LXA4對(duì)LPS誘導(dǎo)BEAS-2B細(xì)胞p38 MAPK通路的影響 p38 MAPK通路在炎癥細(xì)胞因子的產(chǎn)生過程中有著重要的作用。與對(duì)照組比,LPS可顯著上調(diào)p-p38/p38水平(P<0.01),而LXA4能夠明顯抑制p-p38的蛋白表達(dá),差異均有統(tǒng)計(jì)學(xué)意義(P<0.01),見圖3。
2.4 LXA4對(duì)LPS誘導(dǎo)BEAS-2B細(xì)胞Nrf2通路的影響
在LPS的刺激下,BEAS-2B細(xì)胞核內(nèi)Nrf2表達(dá)水平顯著下降(P<0.01),胞漿Nrf2表達(dá)水平上升(P<0.05),Nrf2磷酸化水平下降(P<0.05)。而經(jīng)LXA4預(yù)處理后,核內(nèi)Nrf2表達(dá)水平顯著升高(P<0.01),胞漿Nrf2水平明顯下降(P<0.05),Nrf2磷酸化水平顯著上調(diào)(P<0.01),見圖4。

圖1 各組BEAS-2B細(xì)胞IL-6和IL-1β mRNA水平的比較
2.5 LXA4對(duì)LPS誘導(dǎo)BEAS-2B細(xì)胞Nrf2下游分子表達(dá)的影響 與對(duì)照組比,LPS組HO-1 mRNA表達(dá)水平顯著下降(P<0.01),NQO-1 mRNA水平差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)。LXA4預(yù)處理后,HO-1和NQO-1 mRNA水平出現(xiàn)顯著的上調(diào)(P<0.01),見圖5。
3 討論
研究表明支氣管上皮細(xì)胞在氣道慢性炎癥以及氣管重塑的過程中發(fā)揮著重要作用。支氣管上皮細(xì)胞不僅僅是一層屏障,它可以通過模式識(shí)別受體(Toll-like receptor,TLR)識(shí)別環(huán)境中的刺激物,分泌上皮細(xì)胞來(lái)源的細(xì)胞因子,橋接固有免疫和適應(yīng)性免疫[4]。活化的支氣管上皮細(xì)胞可促進(jìn)Th2型細(xì)胞因子的釋放,募集并激活其他炎性細(xì)胞,發(fā)揮主動(dòng)調(diào)節(jié)作用,同時(shí)誘導(dǎo)上皮下纖維的沉積,促進(jìn)肺部慢性炎癥的發(fā)生發(fā)展[5]。有研究發(fā)現(xiàn),LXA4在哮喘發(fā)病過程中,可以促進(jìn)NK細(xì)胞所介導(dǎo)的粒細(xì)胞凋亡,并且可影響NK細(xì)胞的細(xì)胞毒性作用[6]。本研究發(fā)現(xiàn)LXA4具有抑制支氣管上皮細(xì)胞的炎癥反應(yīng)以及促進(jìn)炎癥消退的作用。在BEAS-2B細(xì)胞中,LXA4預(yù)處理顯著抑制了由LPS引起的促炎因子和ROS的產(chǎn)生,同時(shí)LXA4還誘導(dǎo)抗氧化酶和GSH的表達(dá)顯著上調(diào),并且該作用在一定程度上依賴p38的磷酸化水平和Nrf2的核轉(zhuǎn)位及磷酸化水平。因此,LXA4在氣道炎癥性疾病中發(fā)揮抗炎和抗氧化作用,對(duì)肺部炎癥的預(yù)防和治療具有一定的作用。

圖2 各組BEAS-2B細(xì)胞內(nèi)ROS和GSH水平的比較
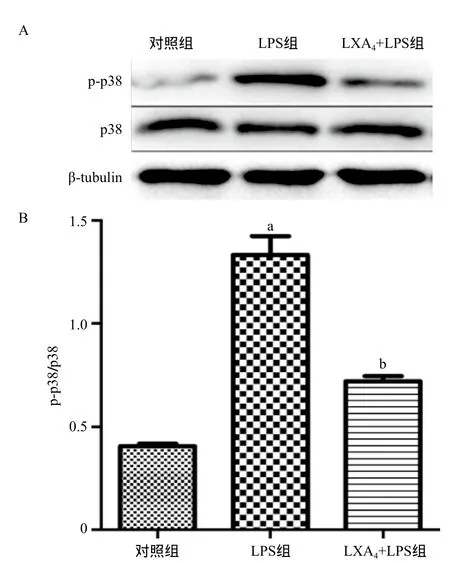
圖3 各組BEAS-2B細(xì)胞內(nèi)p-p38和p38表達(dá)水平的比較
在過敏性哮喘患者中,由氣道上皮細(xì)胞合成與釋放的IL-6可調(diào)節(jié)效應(yīng)性CD4+T細(xì)胞的功能,誘導(dǎo)Th1型細(xì)胞向Th2型細(xì)胞的分化,促進(jìn)炎癥的發(fā)生發(fā)展,造成肺組織損傷[7-9]。IL-1β是肺部炎性級(jí)聯(lián) 反應(yīng)中的關(guān)鍵因子,它可刺激細(xì)胞黏附分子的產(chǎn) 生,選擇性募集炎性細(xì)胞,誘導(dǎo)多種細(xì)胞因子的表達(dá)[10]。因此,抑制促炎細(xì)胞因子的產(chǎn)生是控制肺部炎癥發(fā)生發(fā)展的一個(gè)重要手段。本研究發(fā)現(xiàn)LXA4預(yù)處理可顯著抑制LPS誘導(dǎo)支氣管上皮細(xì)胞IL-1β和IL-6 mRNA的表達(dá)。這與以往在巨噬細(xì)胞、內(nèi)皮細(xì)胞和人視網(wǎng)膜上皮細(xì)胞中的研究[11-12]結(jié)果一致。

圖4 各組BEAS-2B細(xì)胞內(nèi)Nrf2核轉(zhuǎn)位以及磷酸化水平的比較
另外,氣道上皮細(xì)胞也是直接接觸外部環(huán)境刺激,產(chǎn)生ROS引起氣道氧化應(yīng)激反應(yīng)的主要細(xì)胞之一[13]。靜息狀態(tài)下,機(jī)體內(nèi)ROS的水平處于動(dòng)態(tài)平衡,并且維持在極低的水平。過量的ROS也能夠促進(jìn)炎癥因子以及其他炎癥相關(guān)調(diào)節(jié)因子的表達(dá),及時(shí)清除ROS可有效抑制炎癥的發(fā)生發(fā)展。LPS可刺激巨噬細(xì)胞、中性粒細(xì)胞和氣道上皮細(xì)胞ROS的生成,本研究發(fā)現(xiàn),LXA4能夠明顯抑制LPS誘導(dǎo)BEAS-2B細(xì)胞內(nèi)的ROS的產(chǎn)生。另外,GSH是重要的內(nèi)源性抗氧化劑,可直接清除自由基和ROS,保護(hù)過氧化誘導(dǎo)的細(xì)胞損傷。本研究發(fā)現(xiàn)LXA4顯著上調(diào)了LPS誘導(dǎo)的BEAS-2B細(xì)胞胞內(nèi)GSH的合成與表達(dá)。p38 MAPK通路是各種轉(zhuǎn)錄因子的上游信號(hào)調(diào)節(jié)器,如NF-κB、AP-1[14],參與炎癥反應(yīng)、細(xì)胞增殖、細(xì)胞分化和細(xì)胞死亡等過程。有研究發(fā)現(xiàn),在中性粒細(xì)胞中,LXA4可以降低AP-1與DNA的結(jié)合能力,抑制下游炎癥相關(guān)基因的轉(zhuǎn)錄[15]。在氣道上皮細(xì)胞中,抑制p38 MAPK通路可抑制IL-6的合成與分 泌[16-18]。NF-κB通路的激活能夠上調(diào)IL-1β的分泌,后者又能夠進(jìn)一步促進(jìn)NF-κB通路的激活。LPS通過與TLR4結(jié)合可快速活化p38 MAPK途徑,本研究發(fā)現(xiàn)氣道上皮細(xì)胞在LPS的刺激下,p-p38表達(dá)增加,這與氣道炎癥條件下磷酸化的p38表達(dá)水平增高具有一致性,而LXA4可以抑制p38 MAPK途徑的激活。因此,LXA4可能通過調(diào)控p38 MAPK信號(hào)通路抑制促炎因子的表達(dá)。該作用與小膠質(zhì)細(xì)胞中的表達(dá)一致,而在心肌細(xì)胞系H9c2中,LXA4上調(diào)了p-p38的表達(dá)水平[19-20],造成這種差異的主要原因可能跟細(xì)胞類型有關(guān)。

圖5 各組BEAS-2B細(xì)胞內(nèi)HO-1及NQO-1 mRNA水平的比較
研究發(fā)現(xiàn),通過激活Nrf2通路能夠有效抑制氧化應(yīng)激反應(yīng)引起的肺損傷[21]。Nrf2是人體普遍存在的一種抗氧化轉(zhuǎn)錄因子,在抑制LPS誘導(dǎo)的氧化損傷中發(fā)揮著重要的保護(hù)作用[22-23]。在正常狀態(tài)下,Nrf2主要表達(dá)于細(xì)胞質(zhì)內(nèi),與細(xì)胞質(zhì)中肌動(dòng)蛋白結(jié)合蛋白KEAP1相互結(jié)合,保持在一個(gè)低表達(dá)狀態(tài),且易被泛素蛋白酶體途徑迅速降解。當(dāng)細(xì)胞受到氧化劑或毒性物質(zhì)的刺激時(shí),KEAP1的分子構(gòu)象發(fā)生改變,Nrf2可從KEAP1上解離后活化,而活化后的Nrf2轉(zhuǎn)位進(jìn)入細(xì)胞核內(nèi)并與抗氧化反應(yīng)原件(antioxidant response element,ARE)的啟動(dòng)子部位結(jié)合,從而促進(jìn)一系列細(xì)胞保護(hù)基因和抗氧化酶的表達(dá)。Nrf2通路的激活是抗氧化應(yīng)激反應(yīng)有效的保護(hù)機(jī)制。研究發(fā)現(xiàn),COPD患者肺泡巨噬細(xì)胞內(nèi)的HO-1表達(dá)下降[24]。NQO-1和HO-1具有顯著的抗炎和抗氧化作用,兩者的表達(dá)受到Nrf2通路下游ARE的調(diào)控[25]。本研究發(fā)現(xiàn)LXA4能夠促進(jìn)Nrf2的核轉(zhuǎn)位以及磷酸化,同時(shí)ARE下游基因NQO-1和HO-1的表達(dá)增高。因此,LXA4的抗氧化作用可能與Nrf2通路激活,使得抗氧化酶NQO-1和HO-1的表達(dá)水平升高有關(guān)。
綜上所述,LXA4可減輕LPS引起的BEAS-2B細(xì)胞炎癥反應(yīng)并促進(jìn)炎癥消退,其機(jī)制可能一方面與抑制p38 MAPK通路、減少促炎因子的分泌有關(guān);另一方面與增強(qiáng)Nrf2的核轉(zhuǎn)位以及磷酸化、減輕氧化應(yīng)激損傷有關(guān)。
[1] 李春來(lái), 張冰緣, 劉再英. 脂氧素在呼吸系統(tǒng)疾病中的研究進(jìn)展[J]. 中華臨床醫(yī)師雜志(電子版), 2016(11): 197-198.
[2] BORGESON E, JOHNSON A M, LEE Y S, et al. Lipoxin A4 attenuates obesity-induced adipose in flammation and associated liver and kidney disease[J]. Cell Metab, 2015, 22(1): 125-137.
[3] KE Y, ZEBDA N, OSKOLKOVA O, et al. Anti-in flammatory effects of OxPAPC involve endothelial cell-mediated generation of LXA4[J]. Circ Res, 2017, 121(3): 244-257.
[4] QIU Z, ZHOU J, LIU F, et al. Deletion of Shp2 in bronchial epithelial cells impairs IL-25 production in vitro, but has minor in fluence on asthmatic in flammation in vivo[J]. PLoS One, 2017, 12(5): e0177334.
[5] LEONARDI S, VITALITI G, MARSEGLIA G L, et al.Function of the airway epithelium in asthma[J]. J Biol Regul Homeost Agents, 2012, 26(1 Suppl): S41-48.
[6] DUVALL M G, BARNIG C, CERNADAS M, et al. Natural killer cell-mediated inflammation resolution is disabled in severe asthma[J]. Sci Immunol, 2017, 2(9): eaam5446.
[7] hEIJINK I H, NAWIJN M C, HACKETT T L. Airway epithelial barrier function regulates the pathogenesis of allergic asthma[J]. Clin Exp Allergy, 2014, 44(5): 620-630.
[8] KAUR D, BRIGHTLING C. OX40/OX40 ligand interactions in T-cell regulation and asthma[J]. Chest, 2012, 141(2):494-499.
[9] RAJU K R, KUMAR M N, GUPTA S, et al. 5-Aminosalicylic acid attenuates allergen-induced airway inflammation and oxidative stress in asthma[J]. Pulm Pharmacol Ther,2014, 29(2): 209-216.
[10] LI H, YIN J, LI L, et al. Iso flurane postconditioning reduces ischemia-induced nuclear factor-κB activation and interleukin 1β production to provide neuroprotection in rats and mice[J]. Neurobiol Dis, 2013, 54: 216-224.
[11] KURE I, NISHIUMI S, NISHITANI Y, et al. Lipoxin A(4)reduces lipopolysaccharide-induced inflammation in macrophages and intestinal epithelial cells through inhibition of nuclear factor-kappa B activation[J]. J Pharmacol Exp Ther,2010, 332(2): 541-548.
[12] KAVIARASAN K, JITHU M, ARIF M M, et al. Low blood and vitreal BDNF, LXA4 and altered Th1/Th2 cytokine balance are potential risk factors for diabetic retinopa-thy[J].Metabolism, 2015, 64(9): 958-966.
[13] GUAN S P, TEE W, NG D S, et al. Andrographolide protects against cigarette smoke-induced oxidative lung injury via augmentation of Nrf2 activity[J]. Br J Pharmacol, 2013,168(7): 1707-1718.
[14] PARK J, KWAK C H, HA S H, et al. Ganglioside GM3 suppresses lipopolysaccharide-induced inflammatory responses in rAW 264.7 macrophage cells through NF-kappaB, AP-1, and MAPKs signaling[J]. J Cell Biochem, 2018, 119(1):1173-1182.
[15] QI W, LI H, CAI X H, et al. Lipoxin A4 activates alveolar epithelial sodium channel gamma via the microRNA-21/PTEN/AKT pathway in lipopolysaccharide-induced in flammatory lung injury[J]. Lab Invest, 2015, 95(11): 1258-1268.
[16] DOUILLET C D, ROBINSON W P, MILANO P M, et al.Nucleotides induce IL-6 release from human airway epithelia via P2Y2 and p38 MAPK-dependent pathways[J]. Am J Physiol Lung Cell Mol Physiol, 2006, 291(4): L734-746.
[17] FURUSAWA J, MORO K, MOTOMURA Y, et al. Critical role of p38 and GATA3 in natural helper cell function[J]. J Immunol, 2013, 191(4): 1818-1826.
[18] JIN M, KIM S R, YOON S J, et al. Suppressive effects of fructus of Magnolia denudata on IL-4 and IL-13 expression in T cells[J]. In Vitro Cell Dev Biol Anim, 2013, 49(10):805-814.
[19] 周昱, 吳升華, 陳筱青, 等. L X A 4通過p 38 M A P K/N r f 2信號(hào)通路誘導(dǎo)H 9 c 2心肌細(xì)胞H O-1高表達(dá)[J]. 南京醫(yī)科大學(xué)學(xué)報(bào)(自然科學(xué)版), 2012, 32(11): 1493-1498.
[20] WANG Y P, WU Y, LI L Y, et al. Aspirin-triggered lipoxin A4 attenuates LPS-induced pro-in flammatory responses by inhibiting activation of NF-kappaB and MAPKs in BV-2 microglial cells[J]. J Neuroin flammation, 2011, 8: 95.
[21] SMIRNOVA N A, HASKEW-LAYTON R E, BASSO M,et al. Development of Neh2-luciferase reporter and its application for high throughput screening and real-time monitoring of Nrf2 activators[J]. Chem Biol, 2011, 18(6): 752-765.
[22] PARK J, MIN J S, KIM B, et al. Mitochondrial ROS govern the LPS-induced pro-in flammatory response in microglia cells by regulating MAPK and NF-kappaB pathways[J].Neurosci Lett, 2015, 584: 191-196.
[23] YANG H L, LIN M W, KORIVI M, et al. Coenzyme Q0 regulates NFkappaB/AP-1 activation and enhances Nrf2 stabilization in attenuation of LPS-induced in flammation and redox imbalance: Evidence from in vitro and in vivo studies[J]. Biochim Biophys Acta, 2016, 1859(2): 246-261.
[24] FREDENBURGH L E, PERRELLA M A, MITSIALIS S A.The role of heme oxygenase-1 in pulmonary disease[J]. Am J Respir Cell Mol Biol, 2007, 36(2): 158-165.
[25] KIM J, CHA Y N, SURH Y J. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in in flammatory disorders[J]. Mutat Res, 2010, 690(1-2): 12-23.
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國(guó)火炬(2010年12期)2010-07-25 13:26:22
中國(guó)火炬(2010年8期)2010-07-25 11:34:30
