BMP-2對間充質干細胞促進造血干細胞增殖的影響及其機制
2018-04-20 06:40:37李書壇黃遠瓊汪姝玥林凡莉黃純蘭
山東醫藥 2018年12期
李書壇,黃遠瓊,汪姝玥,林凡莉,黃純蘭,3
(1西南醫科大學附屬醫院,四川滬州646000;2瀘州市中醫醫院;3成都市第三人民醫院)
骨髓中的造血干細胞(HSC)具有自我更新和分化為各種成熟血細胞的能力,但其數量少。隨著干細胞移植(HSCT)技術的飛速發展,對HSC的需求增加,使得體外擴增HSC成為當前研究的熱點。HSC增殖、分化和歸巢受骨髓微環境即微龕“Niche”的調控。Niche由基質細胞、其前體間充質干細胞(MSC)以及它們表達和分泌的黏附分子、細胞因子和細胞外基質組成,包括以成骨細胞為主體的骨內膜龕和以內皮細胞為主體的血管微龕。研究顯示,MSC可促進HSC增殖[1~3],但作用機制復雜、尚未闡明。骨形態發生蛋白(BMP)/TGF-β家族在造血調控過程中發揮重要作用,該家族成員形成復雜的信號網絡,調控HSC增殖[4,5]。BMP-2是該家族中的一員,作為一種多功能蛋白,其能促進新骨及血管的形成。近年來研究顯示,BMP-2協同MSC可以促進血管生成[6]。2014年7月~2016年3月我們進行本研究,旨在從成血管角度探討BMP-2協同MSC對HSC增殖的影響及可能的機制。
1 材料與方法
1.1材料LG-DMEM培養基、FBS、青霉素、鏈霉素、0.25%胰蛋白酶(Hyclone公司,美國),鼠抗人ECD-CD34、鼠抗人PEcy7-CD45、鼠抗人FITC-CD105、鼠抗人IgG抗體(Pharmingen公司,美國),重組人干細胞因子(rhSCF)、重組人白介素3(rhIL-3)、rhBMP-2(RD公司,美國);Mini MACS免疫磁性吸附柱分離裝置和CD34+細胞選擇試劑盒(Mitenyi Biotec公司,德國),反轉錄試劑盒(TOYOBO公司,日本),熒光PCR試劑盒(QIAGEN公司,德國),TRIzol試劑盒(Invitrogen 公司,美國),Transwell小室(微網孔徑0.4 μm,Costar公司,美國)。β-actin、Ki67、血管生成素受體2(Tie2)、血管內皮生長因子(VEGF)、血管生成素1(Ang1)引物,均由上海生工生物工程股份有限公司合成。骨髓來自西南醫科大學附屬醫院健康志愿者,其中男9例、女4例,年齡21~47(32.72±2.01)歲。本研究經西南醫科大學附屬醫院倫理委員會批準,健康志愿者知情同意并簽字。
1.2MSC擴增和鑒定取健康志愿者髂骨骨髓,肝素抗凝管收集,按1∶4體積與完全培養基重置(完全培養基成分為89% LD-DMEM +10% FBS+1%青/鏈霉素),混合后接種于25 cm2塑料培養瓶中,于37 ℃、5% CO2孵箱中培養。每3天換液一次,待細胞融合達80%后用0.25%胰蛋白酶消化傳代,傳至第3代備用。倒置相差顯微鏡觀察MSC形態及貼壁、融合等生長情況。顯微鏡下顯示,全骨髓法原代培養MSC 72 h可見散在的梭形貼壁細胞;第14天時梭形細胞融合度可達80%,排列成放射狀、旋渦狀;第3代MSC在培養6天時融合度可達80%。原代培養細胞生長周期偏長,傳代細胞增殖較快且背景干凈。采用流式細胞儀檢測第3代MSC表面抗原CD34、CD45、CD105表達。結果顯示, CD34、CD45、CD105陽性表達率分別為2.2%、0.3%、52.4%。提示MSC分離培養成功。流式細胞儀檢測MSC的純度為52.4%。
1.3HSC分選及純度鑒定取健康志愿者髂骨骨髓,經預冷的PBS稀釋后輕緩地加入Ficoll分離液上層,4 ℃下2 000 r/min離心20 min,收集白色細胞層,分選出單個核細胞,采用Mini MACS免疫磁性吸附柱分離裝置,用CD34+細胞(即HSC)選擇試劑盒進行CD34+細胞分離,并用抗CD34-ECD抗體檢測分選細胞的純度。臺盼藍染色顯示HSC呈大小均勻的圓形,活性大于95%;流式細胞儀檢測顯示,HSC純度為81.9%。
1.4MSC對HSC增殖的影響將細胞分組,HSC組為HSC單獨培養,HSC+MSC組為HSC、MSC共培養;每組3個復孔。將融合達80%的第3代MSC接種于Transwell下室,CD34+HSC細胞接種于上室,前者接種細胞量為3.20×105個/孔,后者接種細胞量為1.25×104個/孔,培養基為含10% FBS、100 U/mL青霉素及0.1 mg/mL鏈霉素的IMDM完全培養基,每2天每孔添加rhSCF 50 ng/mL、rhIL-3 20 ng/mL,并換液。分別于培養第3、7、10天收集HSC并用計數板計數,確定HSC增殖最快的時間點。結果顯示,HSC增殖最快的時間點為共培養第7天。
1.5BMP-2對MSC促HSC增殖的影響將細胞分為六組,分別為HSC組、MSC組、HSC+BMP-2組、MSC+BMP-2組、HSC+MSC組、HSC+MSC+BMP-2組。HSC組、 MSC組單細胞培養,MSC+BMP-2組、HSC+MSC組分別加入100 ng/mL BMP-2培養,HSC+MSC組兩種細胞共培養,HSC+MSC+BMP-2組兩種細胞與100 ng/mL BMP-2共培養;細胞接種密度、培養條件同1.4。于培養第3、5天時更換培養基并添加細胞因子rhBMP-2,培養第7天時行以下指標檢測。
1.5.1HSC計數采用加樣槍分別吸取10 μL各組細胞懸液放入細胞計數板。顯微鏡下計數四角大方格內的細胞總數,壓線細胞只計左側和下方。計算公式:細胞數/mL=四角大方格內的細胞總數/4×104。
1.5.2增殖因子Ki67及促血管形成相關因子Tie2、VEGF、Ang1 mRNA表達采用實時熒光定量PCR法檢測。收集各組細胞,采用TRIzol法提取RNA,用分光光度計檢測RNA濃度、A260/A280。將RNA稀釋,采用模板量取1 ng,經逆轉錄酶轉錄成cDNA。以cDNA為擴增模板,采用熒光定量qRT-PCR法檢測各組CD34+細胞的Ki67、Tie2、VEGF、Ang1 mRNA。每組接種3個復孔,計算Ct值。采用2-ΔΔCt法計算上述因子mRNA相對表達量。Ki67、Tie2、VEGF、Ang1基因及內參基因β-actin的引物序列見表1。

表1 Ki67、Tie2、VEGF、Ang1及β-actin的引物序列
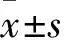
2 結果
2.1各組HSC數量比較HSC組、HSC+BMP-2組、HSC+MSC組、HSC+MSC+BMP-2組HSC數量分別為(15.37±0.42)、(37.81±1.07)、(117.00±2.53)、(129.06±5.13)×104個,組間比較P<0.05或<0.01。
2.2各組Ki67及Tie2、VEGF、Ang1 mRNA表達比較HSC組、HSC+BMP-2組、HSC+MSC組、HSC+MSC+BMP-2組Ki67及Tie2 mRNA相對表達量逐漸升高,MSC組、MSC+BMP-2組、HSC+MSC組、HSC+MSC+BMP-2組VEGF、Ang1 mRNA相對表達量逐漸升高,組間比較P<0.05或<0.01。見表2。
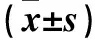
表2 各組Ki67及Tie2、VEGF、Ang1 mRNA相對表達量比較
注:“-”表示本組細胞未進行對應基因mRNA檢測。
3 討論
CD34是HSC表面重要的抗原,在臨床及科研工作中常用該抗原分選HSC。目前MSC的培養方法主要包括全骨髓黏附培養法、密度梯度離心法、免疫表型分選法等,全骨髓法因其經濟適用、所培養的細胞活性高,通過傳代可達到擴增純化的目的,是目前培養MSC的重要方法。本研究應用免疫磁珠法及全骨髓法獲得的HSC和MSC的純度分別為81.9%、52.4%,純度較高,能保證后續實驗研究的進行。HSC作為機體重要的干細胞,研究其增殖、凋亡等生理活動有利于探索造血以及腫瘤等疾病的發生機制。隨著干細胞移植技術的發展,HSC已成為治療血液系統惡性疾病、一些遺傳性和免疫性疾病以及部分惡性實體腫瘤的有效途徑之一。然而HSC數量少,體外擴增HSC勢在必行。研究發現,如前所述MSC可促進HSC增殖,這種擴增效應依賴于細胞間的相互作用和分泌的細胞因子[7,8],de Silva等[8]將HSC與MSC分別進行非接觸共培養、接觸共培養、單獨培養,結果顯示在接觸與非接觸兩種情況下MSC均可以促進HSC增殖,且MSC與HSC接觸培養對HSC的增殖促進作用最強。本研究應用Transwell將MSC與HSC進行非接觸共培養,結果發現MSC可以非接觸促進HSC增殖。
研究發現,MSC能分泌干細胞因子(SCF)、IL、集落刺激因子(GSF)、基質細胞衍生因子1(SDF-1)、基質金屬蛋白酶組織抑制劑3(TIMP-3)等多種細胞因子、黏附分子,它們形成復雜的信號網絡可促進HSC增殖[7,9]。目前研究顯示,這些信號通路主要包括CXCLl2/CXCR4、Notch、Wnt、Tie2/Angl、BMP/TGF-β、Hedgehog[2,9,10]。其中,BMP/TGF-β家族調節多種細胞的增殖、凋亡、分化等生理活動。該家族包括TGF-β和BMP兩個亞家族,前者由TGF-β、Activin和Nodal等組成,后者由BMP、GDF和AMH 組成。現發現的20多種BMP蛋白分布于多種組織,目前研究較多的主要為BMP-2、BMP-4、BMP-7、BMP-10等,其受體BMPR包括BMPR-IA,BMPR-IB和BMPR-Ⅱ,它們屬于絲氨酸/蘇氨酸激酶家族。BMP-2可以與HSC的BMPRI結合并促進其下游轉錄子Smad-1、Smad-4、Smad-5表達進而調控HSC[11]。BMP蛋白是多功能蛋白,大多能促進骨重建及生長,是胚胎和器官發育的重要因子,在胚胎期能調控造血[3,12,13],其在成體中的造血調控作用目前研究較少。BMP對造血的調控機制復雜,不同的BMP具有不同的作用,如BMP-4抑制HSC向紅系分化,BMP-6抑制人B淋巴細胞形成,即使是同種BMP對HSC的同種生理功能也可能不同[14]。目前研究認為這可能與靶細胞的分化程度[12]、細胞因子亞型及濃度、甚至生長因子來源于旁分泌還是自分泌等有關[15]。BMP-2對HSC增殖的影響與培養模式、BMP-2濃度以及所聯合的細胞因子種類等具體環境相關。Bhatia等[16]發現,BMP-2可引起NOD/SCID小鼠HSC數目、集落形成、細胞表型和多系分化能力的改變,且影響程度與劑量有關;高濃度(50 ng/mL)BMP-2會抑制細胞增殖,低濃度(5 ng/mL)BMP-2則對HSC增殖影響較小。將BMP-2與不同的細胞因子聯合培養HSC,發現SCF聯合GM-CSF能促進HSC增殖[17],而BMP-2與EPO和IL-3[17]、SCF和SF3[18]、SCF和SF3以及TPO[19]聯合則無協同促進作用。 本研究結果顯示,濃度為100 ng/mL的BMP-2聯合IL-3、SCF可促進成人HSC增殖,且BMP-2協同MSC亦能促進HSC增殖。然而,BMP-2對造血的哪個階段進行調節,HSC增殖的具體機制以及增殖后的HSC是否分化目前尚不清楚,有待進一步研究。
增殖指數Ki67基因與細胞增殖相關,能反映細胞增殖的活躍程度,其核蛋白在細胞G1、S、G2和M期均有表達,G0期則不表達。Ki67陽性率越高,說明細胞增殖越活躍。血管生成受多種信號通路的調節,其中最重要的是VEGF/血管內皮生長因子受體(VEGFR)通路。VEGF包括VEGF-A、-B、-C、-D、-E和胎盤生長因子,其中VEGF-A和VEGF-B與血管生成相關[20,21]。VEGF具有高度特異性,通過刺激內皮細胞增殖促進血管生成[21]。VEGFR屬于酪氨酸激酶受體超家族(RTK),由跨膜區、七個免疫球蛋白樣結構域組成的胞外區和酪氨酸激酶胞內區三部分組成[22],與血管形成相關的主要有VEGFR-1、VEGFR-2、VEGFR-3,在內皮細胞均有表達。Ang/Tie2通路在血管形成中發揮重要作用。Ang主要包括Ang1和Ang2,它們共同的受體為Tie2。Ang1在正常組織中表達廣泛,參與血管新生及血管成熟[23]。Ang1啟動Tie2受體后使內皮周圍平滑肌細胞和血管周細胞募集至血管內皮細胞周圍,通過增加內皮細胞與周圍支持細胞之間的相互作用,維持血管的完整性與穩定性,并降低其通透性。Ang2是Ang1的天然拮抗劑,主要分布于子宮、卵巢和胎盤等部位。Tie2是RTK家族成員,在內皮細胞及某些造血祖細胞中特異性表達[24]。相比于成骨Niche,血管Niche能促進HSC增殖、分化[25]。研究發現,BMP可促進成骨細胞及BMP介導的MSC向成骨細胞分化時VEGF及Ang1表達[26]。本研究發現,HSC組、HSC+BMP-2組、HSC+MSC組、HSC+MSC+BMP-2組Ki67及Tie2 mRNA相對表達量逐漸升高,MSC組、MSC+BMP-2組、HSC+MSC組、HSC+MSC+BMP-2組VEGF、Ang1 mRNA相對表達量逐漸升高,說明BMP-2及BMP-2協同MSC均可通過促進血管形成而促進HSC增殖。
綜上所述,BMP-2協同MSC可促進HSC增殖,其機制可能與促血管形成相關因子VEGF、Ang1和Tie2表達增加相關。
參考文獻:
[1] Raynaud CM, Butler JM, Halabi NM, et al. Endothelial cells provide a niche for placental hematopoietic stem/progenitor cell expansion through broad transcriptomic modification[J]. Stem Cell Res, 2013,11(3):1074-1090.
[2] 朱小鳳,黃純蘭,李曉明.骨髓間充質干細胞聯合外源性Shh蛋白對骨髓造血干細胞增殖影響[J].山東醫藥,2017,57(37):31-33.
[3] Bai H, Xie YL, Gao YX, et al. The balance of positive and negative effects of TGF-β signaling regulates the development of hematopoietic and endothelial progenitors in human pluripotent stem cells[J]. Stem Cells Dev, 2013,22(20):2765-2776.
[4] Robin C, Durand C. The roles of BMP and IL-3 signaling pathways in the control of hematopoietic stem cells in the mouse embryo[J]. Int J Dev Biol, 2010,54(6):1189-2000.
[5] Francois S, Eder VV, Belmokhtar K, et al. Synergistic effect of human bone morphogenic protein-2 and mesenchymal stromal cells on chronic wounds through hypoxia-inducible factor-1α induction[J]. Sci Rep, 2017,7(1):42-72.
[6] Rodríguez-Pardo VM,Vernot JP. Mesenchymal stem cells promote a primitive phenotype CD34+c-kit+in human cord blood-derived hematopoietic stem cells during ex vivo expansion[J]. Cell Mol Biol Lett, 2013,18(1):11-33.
[7] Andrade PZ, Santos F, Almeida-Porada G, et al. Systematic delineation of optimal cytokine concentrations to expand hematopoietic stem/progenitor cells in co-culture with mesenchymal stem cells[J]. Mol Biosyst, 2010,6(7):1207-1215.
[8] de Silva CL, Gon?alves R, dos Santos F, et al. Dynamic cell-cell interactions between cord blood haematopoietic progenitors and the cellular niche are essential for the expansion of CD34+, CD34+CD38-and early lymphoid CD7+cells[J]. J Tissue Eng Regen Med, 2010,4(2):149-158.
[9] Moll NM, Ransohoff RM. CXCL12 and CXCR4 in bone marrow physiology[J]. Expert Rev Hematol, 2010,3(3):315-322.
[10] Chen LZ, Yin SM, Zhang XL, et al. Protective effects of human bone marrow mesenchymal stem cells on hematopoietic organs of irradiated mice[J]. Zhongguo Shi Yan Xue Ye Xue Za Zhi, 2012,20(6):1436-1441.
[11] Utsugisawa T, Moody JL, Aspling M, et al. A road map toward defining the role of Smad signaling in hematopoietic stem cells[J]. Stem Cells, 2006,24(4):1128-1136.
[12] Lempereur A, Canto PY, Richard C, et al. The TGFβ pathway is a key player for the endothelial-to-hematopoietic transition in the embryonic aorta[J]. Dev Biol, 2017,12(17):307-320.
[13] Crisan M, Solaimani Kartalaei P, Neagu A, et al. BMP and hedgehog regulate distinct AGM hematopoietic stem cells ex vivo[J]. Stem Cell Reports, 2016,6(3):383-395.
[14] S?derberg SS, Karlsson G, Karlsson S. Complex and context dependent regulation of hematopoiesis by TGF-β superfamily signaling[J]. Ann N Y Acad Sci, 2009(1176):55-69.
[15] Kiel MJ, Morrison SJ. Uncertainty in the niches that maintain haematopoietic stem cells[J]. Nat Rev Immunol, 2008,8(4):290-301.
[16] Bhatia M, Bonnet D, Wu D, et al. Bone morphogenetic proteins regulate the developmental program of human hematopoietic stem cells[J]. Exp Med, 1999,189(7):1139-1148.
[17] Detmer K, Walker AN. Bone morphogenetic proteins act synergistically with haematopoietic cytokines in the differentiation of haematopoietic progenitors[J]. Cytokine, 2002,17(1):36-42.
[18] Grassinger J, Simon M, Mueller G, et al. Bone morphogenetic protein (BMP)-7 but not BMP-2 and BMP-4 improves maintenance of primitive peripheral blood-derived hematopoietic progenitor cells (HPC) cultured in serum-free medium supplemented with early actin cytokines[J]. Cytokine, 2007,40(3):165-171.
[19] Su YH, Cai HB, Ye ZY, et al. BMP-7 improved and hematopoietic reconstitution potential of ex vivo expanded cord blood-derived CD34(+) cells[J]. Hum Cell, 2015,28(1):14-21.
[20] Yang XJ, Zhang Y, Yang YL, et al. Vascular endothelial growth factordependent spatiotemporal dual roles of placental growth factor in modulation of angiogenesis and tumor growth[J]. P Natl Acad Sci U S A, 2013,1(10):13932-13937.
[21] Leopol′d AV, Baklaushev VP, Korchagina AA, et al. Ligandreceptor assay for evaluation of functional activity of human recombinant VEGF and VEGFR-1 extracellular fragment[J]. B Exp Biol Med, 2012,152(6):707-711.
[22] Shibuya M. VEGF-VEGFR signals in health and disease[J]. Biomol Ther, 2014,22(1):1-9.
[23] Fagiani E, Christofori G. Angiopoietins in angiogenesis[J]. Cancer Lett, 2013,328(1):18-26.
[24] Makinde T, Agrawal DK. Intra and extravascular transmembrane signalling of angiopoietin-1-Tie2 receptor in health and disease[J]. J Cell Mol Med, 2008,12(3):810-828.
[25] Mercier FE, Ragu C, Scadden DT. The bone marrow at the crossroads of blood and immunity[J]. Nat Rev Immunol, 2012,12(1):49-60.
[26] Ellis SL, Grassinger J, Jones A, et al. The relationship between bone, hemopoieticstem cells, and vasculature[J]. Blood,2011(18):1516-1524.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
