干旱脅迫對八寶景天葉片生理的影響
2018-04-19 09:50:59劉紫娟李炳言賈斯淳宗毓錚郝興宇
山西農業科學 2018年4期
劉紫娟,袁 蕊,王 娜,李炳言,賈斯淳,宗毓錚,李 萍,郝興宇
(山西農業大學農學院,山西太谷 030801)
工業革命以來,全球氣溫持續上升,氣候變暖同時也影響全球水資源的分布、降水量的變化,導致部分地區干旱[1]。水分是植物生長的必需底物,植物的基因表達和新陳代謝都會受到干旱的影響。我國是缺水國家,在干旱和半干旱地區,由于降水量少、土壤水分缺乏,不僅造成作物減產,也嚴重影響了林業發展和生態環境[2]。因此,探究植物適應干旱脅迫的生理機制對干旱區植物生長和生態發展具有一定意義。
八寶景天(Hylotelephium erythrostictum)又名八寶、活血三七、對葉景天,為景天科八寶屬多年生草本植物[3]。八寶景天是景天酸代謝(CAM)植物,葉肉質,喜強光和干燥,管理方式粗放,病蟲害較少,是一種常見的植物,由于其具有較高的觀賞價值和較低的養護成本,在城市建設中被廣泛應用于園林綠化,特別是近些年快速發展的屋頂綠化中[4]。CAM植物由于其代謝方式特殊,均有一定的抗旱性,國內外學者針對CAM植物已有許多研究,多探討CAM植物的光合代謝方式[5-7],針對其抗旱性的研究主要集中在鐵皮石斛、垂盆草、東南景天、細葉景天、紅景天等[8-10],對八寶景天的相關研究較少,特別是一天中白天和夜晚生理變化的研究還未見報道。
本試驗擬探究干旱脅迫條件下八寶景天的葉片生理變化特征,旨在為CAM植物的抗旱機理研究奠定基礎,為八寶景天今后的應用和發展提供理論依據。
1 材料和方法
1.1 試驗材料
供試景天品種為八寶,由山西農業大學園藝學院提供。
1.2 試驗設計
試驗在山西農業大學試驗基地(37.42°N,112.58°E)旱棚中進行,旱棚高3.5 m,面積為4 m×4 m。八寶景天于2016年4月24日移植于長×寬×高為60 cm×40 cm×35 cm塑料箱中,箱子底部打5個孔用于排水,箱內裝土約28cm高,2016年10月20日割除八寶景天的地上部,留根于土中,2017年3月25日發芽。共栽植10盆八寶景天,前期正常灌水。5盆作對照處理(CK),植株發芽后控制土壤含水量為土壤飽和含水量的70%~80%;另外5盆作干旱處理(T),植株發芽30 d后(之前無水分脅迫)控制土壤含水量為土壤飽和含水量的30%~40%,其他管理措施一致。
1.3 生育期的確定
八寶景天開始發芽至發芽后107 d為營養生長期,期間植株莖伸長增粗,葉片增多,發芽后107~118 d陸續有植株長出花蕾,發芽后第119天有80%以上植株長出花蕾,進入現蕾期。
1.4 測定項目及方法
1.4.1 葉片表面氣孔的測定 在營養生長期(發芽后102 d)10:00和22:00,每個處理分別選取有代表性的八寶景天5株(每箱1株),每株選取倒數第7片完全展開葉,用指甲油粘片法測量并計算氣孔密度[11];在40倍物鏡下選取視野內8個氣孔,測量其長度、寬度,并測量氣孔開度的長度和的寬度,計算氣孔開度。氣孔開度(c)=π·ab,其中,a=1/2 氣孔開度的長度,b=1/2氣孔開度的寬度[12]。
1.4.2 葉片抗逆指標的測定 在營養生長期(發芽后 102 d)和現蕾期(發芽后 119 d)的 10:00和22:00,每個處理分別選取有代表性的植株5株(每箱1株),每株選取其倒數第5片完全展開葉,用于測定抗逆生理指標。采用愈創木酚比色法測定過氧化物酶(POD)活性,采用硫代巴比妥酸顯色法測定丙二醛(MDA)含量,采用酸性茚三酮比色法測定脯氨酸(Pro)含量[13]。
1.4.3 葉片含水量的測定 利用1.4.2選取的葉片,剪成0.5 cm小段后測鮮質量,然后于60℃烘箱中12 h烘干,測干質量。含水量=(鮮質量-干質量)/鮮質量×100%。
1.4.4 葉片蘋果酸含量的測定 利用1.4.2選取的葉片,采用Goodban法進行蘋果酸含量的測定[7]。
1.4.5 葉片色素含量的測定 利用1.4.2選取的葉片,用丙酮∶乙醇∶蒸餾水為4.5∶4.5∶1的混合液作為葉綠素提取液,再用分光光度計測定吸光值,并計算光合色素含量。
1.4.6 葉片糖類物質含量的測定 利用1.4.2選取的葉片,采用3,5-二硝基水楊酸法測定還原糖含量,采用蒽酮-硫酸法測定可溶性糖、淀粉和纖維素含量[14]。
1.4.7 各項形態指標的測定 于植株現蕾期將地上部分收獲,對未取樣植株分別測量其株高、莖粗、葉片數,于60℃烘箱48h烘干,測定單株總生物量。
1.5 數據處理
利用Excel 2007軟件對數據進行統計分析和作圖。用SPSS19軟件對數據進行顯著性分析(α=0.05)。圖表中數據均為平均值±標準差。
2 結果與分析
2.1 干旱脅迫對八寶景天葉片氣孔的影響
干旱脅迫使八寶景天葉片上下表皮細胞變小,上下表皮的氣孔長度和開度減小,但氣孔密度增加(圖1)。與對照相比,干旱脅迫使八寶景天葉片上表皮的氣孔長度在10:00和22:00分別顯著降低19.9%和17.3%,10:00和22:00的氣孔開度分別顯著降低30.7%和16.6%,但是氣孔密度分別顯著增加36.8%和28.8%;下表皮的氣孔長度在10:00和22:00分別顯著降低23.7%和20.4%,氣孔開度在10:00和22:00分別顯著降低49.2%和11.5%,但 氣孔密度增加(表1)。

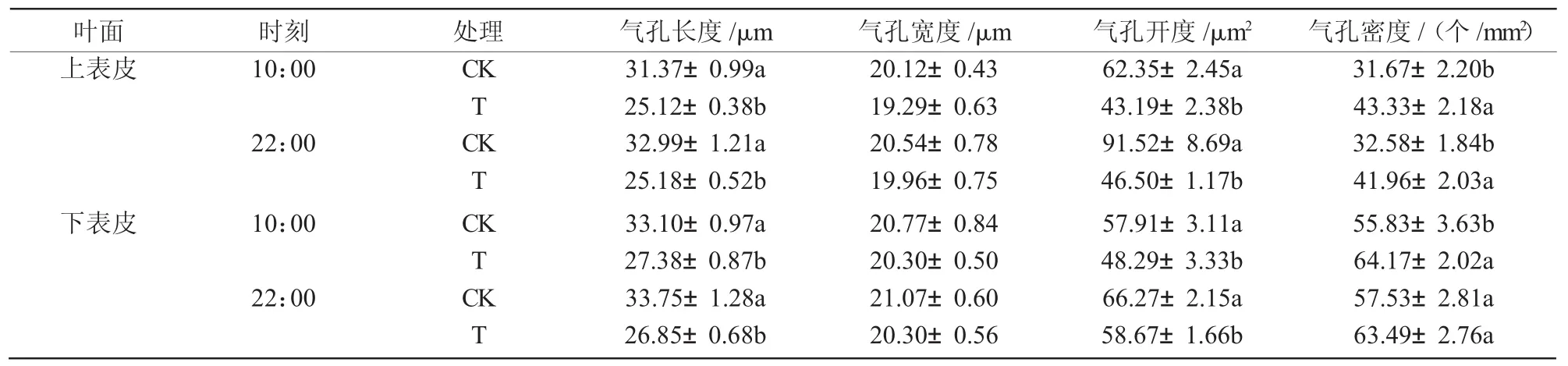
表1 干旱脅迫對八寶景天氣孔的影響
2.2 干旱脅迫對八寶景天葉片含水量的影響
干旱脅迫下,八寶景天葉片含水量在營養生長期和現蕾期的10:00和22:00均無顯著變化(圖 2)。

2.3 干旱脅迫對八寶景天葉片MDA含量的影響
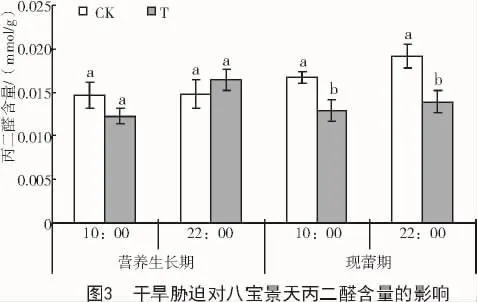
干旱脅迫使八寶景天葉片MDA含量在營養生長期 10:00,現蕾期 10:00 和 22:00 均有降低,在營養生長期22:00有所升高。其中,現蕾期10:00和22:00分別比對照顯著降低22.6%和27.5%(圖3)。
2.4 干旱脅迫對八寶景天葉片POD活性的影響
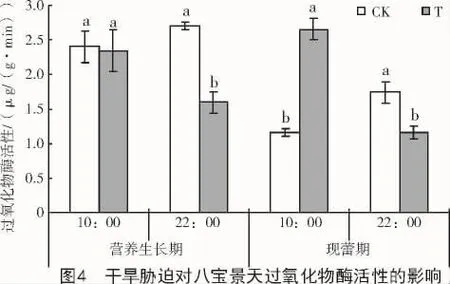
與對照相比,干旱脅迫使八寶景天葉片POD活性在營養生長期和現蕾期的22:00分別顯著降低41.0%和33.7%,在現蕾期10:00顯著升高128.3%,但在營養生長期10:00無明顯變化(圖4)。
2.5 干旱脅迫對八寶景天葉片脯氨酸含量的影響
與對照相比,干旱脅迫使八寶景天脯氨酸含量在營養生長期10:00和22:00分別顯著升高3.5%和1.8%,但在現蕾期脯氨酸含量變化不顯著(圖5)。

2.6 干旱脅迫對八寶景天葉片蘋果酸含量的影響
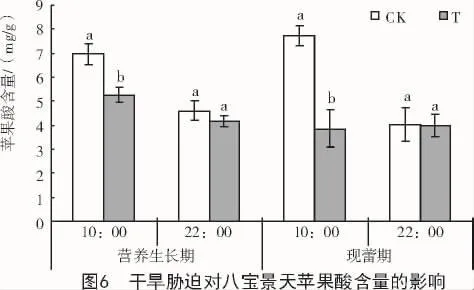
干旱脅迫條件下,八寶景天葉片蘋果酸含量在營養生長期和現蕾期10:00分別比對照顯著降低24.7%和50.1%,但在2個時期22:00均無明顯變化(圖 6)。
2.7 干旱脅迫對八寶景天葉片光合色素的影響
在干旱脅迫條件下,八寶景天葉片的葉綠素a、葉綠素b和類胡蘿卜素含量在營養生長期和現蕾期的10:00均為升高趨勢,其中,葉綠素a含量在營養生長期和現蕾期10:00分別比對照顯著升高36.4%和32.5%;而在2個時期的22:00均呈下降趨勢,其中,營養生長期22:00葉綠素b含量顯著降低24.4%,現蕾期22:00葉綠素a含量顯著降低23.8%(表 2)。
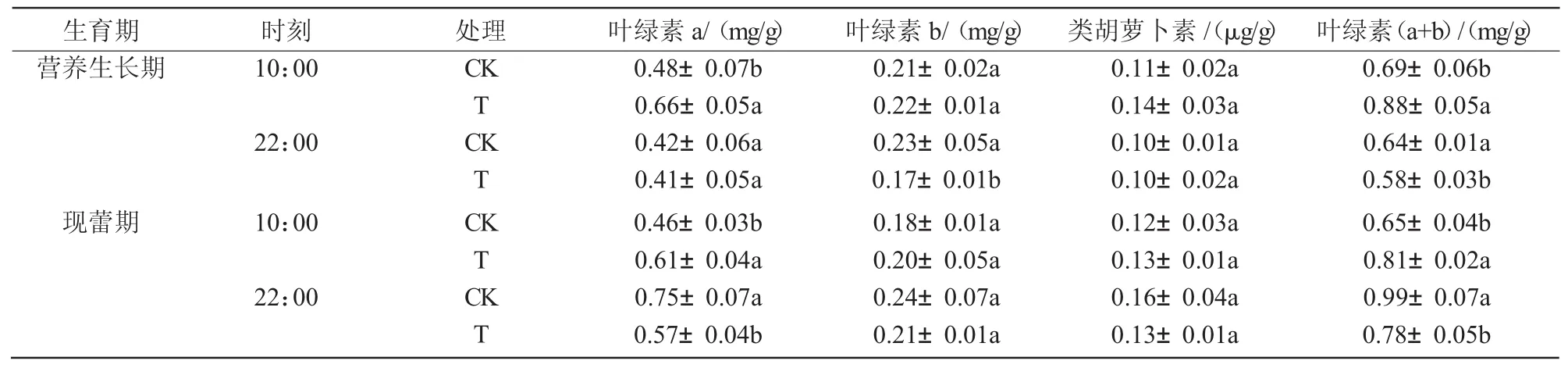
表2 干旱脅迫對八寶景天光合色素的影響
2.8 干旱脅迫對八寶景天葉片糖類物質含量的影響
與對照相比,干旱脅迫使八寶景天葉片中還原糖、可溶性糖、淀粉和纖維素在整個生育期全天總體上呈上升趨勢,其中,還原糖含量在2個時期的22:00分別升高24.6%和45.8%,可溶性總糖含量在現蕾期10:00和22:00分別顯著升高25.3%和24.7%,淀粉含量在營養生長期10:00,22:00和現蕾期10:00分別顯著升高40.3%,35.9%和147.1%,纖維素含量在營養生長期10:00和22:00分別顯著升高31.7%和25.2%,但在現蕾期卻呈下降趨勢,其中,10:00 時顯著下降 36.6%(表 3)。
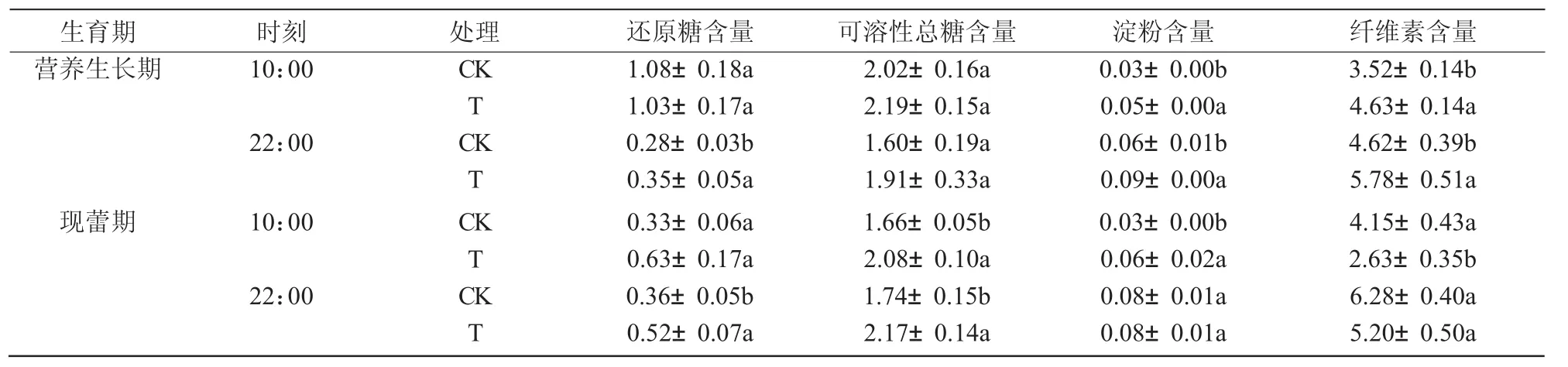
表3 干旱脅迫對八寶景天葉片糖類含量的影響 %
2.9 干旱脅迫對八寶景天各項形態指標的影響
與對照相比,干旱脅迫使八寶景天株高、莖粗、單株葉數和單株總生物量分別顯著降低了36.2%,32.3%,24.6%和35.2%(表4)。

表4 干旱脅迫對八寶景天各項形態指標的影響
3 討論
以往的研究通常認為,干旱脅迫會促使植物葉片氣孔關閉或密度減小,從而減少植物蒸騰作用中水分的散失,提高植物的抗旱性[15];但也有研究顯示,在干旱脅迫下,植物氣孔密度增大,但其長度、寬度和開度均減小[16]。本試驗中,八寶景天在干旱脅迫下葉片上下表皮氣孔的長度、寬度和開度均減小,但氣孔密度增加。氣孔密度增加的原因可能是由于干旱景天葉面積減小,使單位葉面積氣孔密度增加。而氣孔長度、寬度和開度減小的原因是由于干旱影響了氣孔的發生、分化和發育,使氣孔變小。氣孔變小使蒸騰減弱,提高水分利用效率[17]。
干旱脅迫會破壞植物的細胞膜系統,使植物體內自由基增加,從而引起膜脂過氧化作用,膜脂過氧化的產物是MDA,其含量的多少可表示膜脂的過氧化程度和植物的抗逆性[18]。本試驗中,干旱脅迫使八寶景天MDA含量在營養生長期10:00和現蕾期均降低,可能是由于八寶景天具有較強的抗旱性,干旱對葉片的膜脂過氧化作用影響不大。植物中的膜脂過氧化防御系統會保護細胞免遭傷害,POD是植物體內清除H2O2的主要酶類之一,能催化H2O2氧化后產生H2O[19]。本試驗中,干旱脅迫使八寶景天POD活性在22:00時均下降,現蕾期10:00升高,白天溫度較高,可能需要更多的POD防御膜脂的過氧化,夜間溫度降低,干旱脅迫有所緩解,所以,POD活性下降。Pro是一種滲透調節物,可以增強植物的抗旱能力,本試驗中,干旱脅迫下八寶景天葉片的Pro含量除現蕾期10:00外均增加,這也反映出八寶景天是一種抗旱植物。干旱脅迫下植物葉片的含水量可以反映植物保水能力的大小。本試驗中,干旱脅迫對八寶景天葉片的含水量并無顯著影響,說明八寶景天在通過氣孔調節和Pro、糖類物質等滲透調節物質的調節后,在干旱脅迫下有較好的保水能力。
八寶景天是CAM植物,光合方式有其特殊性,表現為:夜間氣孔開放,吸收CO2,以蘋果酸的形式貯存,白天蘋果酸分解,釋放CO2,進行光合作用[7]。本試驗中,干旱脅迫使八寶景天葉片蘋果酸含量在上午10:00顯著下降,說明干旱脅迫抑制了夜間蘋果酸的積累,這是因為干旱脅迫使氣孔開度減小,影響了CO2的吸收。
葉綠素a、葉綠素b和類胡蘿卜素是植物進行光合作用所需的最主要的色素,其含量的多少會直接影響植物的生長發育[20]。本試驗中,干旱脅迫使八寶景天的葉綠素a、葉綠素b和類胡蘿卜素含量在營養生長期和現蕾期10:00升高,但22:00下降,這與之前的研究結果有所不同,可能是由于八寶景天在干旱條件下會產生更多光合色素促使光合作用的進行,但在夜間不進行光反應時,為維持自身正常生理活動,光合色素含量下降。但其調節機制有待進一步研究探索。
可溶性糖也是植物體內參與滲透調節的重要物質,植物在遭受脅迫時,可以通過調節可溶性糖的多少來維持正常的滲透壓,淀粉在植物遭受逆境時可以與可溶性糖相互轉化,纖維素是細胞壁的主要組成部分,可以使植物保持挺拔[21-23]。本試驗中,還原糖、可溶性糖和淀粉含量在不同時期均有增加的趨勢,這也是八寶景天抵抗干旱的一種形式。纖維素含量在營養生長期均增加,但在現蕾期降低,可能是由干旱后期植株缺水時間過長、碳素積累受限所致。
干旱脅迫會使植株的生長形態發生變化,表現在細胞、組織、器官個體等不同水平[24-25]。一般認為,干旱脅迫會使植物的株高、莖粗、生物量均降低,導致作物減產[26]。本試驗中,干旱脅迫使八寶景天株高、莖粗、單株葉片數和單株生物量都降低,葉片的減少和植株矮小可以減少植株水分的散失和植物對水分的需求,有利于植株抵抗干旱環境。
在投融資方案上充分發揮政府投資對社會資本的引導放大作用,吸引社會資本參與。要堅持以市場為導向,以制度建設為重點,形成市場融資為主導、政府投資為補充、民間投資健康發展的投融資格局。在旅游、醫療、教育、停車等社會化產業服務領域,堅持以政策導向引領,社會資本主導參與建設為原則,通過出讓特許經營權、第三方付費、使用者付費結合少量政府補助的方式,鼓勵社會資本加大此類應用的建設力度和速度。利用社會資本的靈活性和企業服務的創新性實現智慧城市的長期可持續發展。
4 結論
本研究表明,干旱脅迫下,八寶景天葉片氣孔變小,氣孔開度降低,能夠有效地減少水分散失,同時影響夜間CO2的吸收,降低了蘋果酸的積累;干旱脅迫使八寶景天MDA含量降低,使POD活性白天升高,夜間降低。現蕾期Pro含量、可溶性總糖和淀粉含量均升高,以提高其抗旱性;干旱脅迫下,八寶景天葉片光合色素含量在白天增多,夜晚降低,有利于其光合效率的提高。干旱使八寶景天株高、莖粗、單株葉片數和單株總生物量均降低,可以有效地減少水分消耗。總之,八寶景天在干旱脅迫下通過氣孔開度變小,POD活性增加進行膜脂過氧化防御,Pro和可溶性糖含量增加進行滲透調節,提高日間光合色素含量保證光合作用的有效進行等方式提高其干旱適應能力,表現了較強的抗旱性。
參考文獻:
[1]WALLACE J S.Increasing agricultural water use efficiency to meet future food production[J].Agriculture,Ecosystems and Environment,2000,82:105-119.
[2]魯松.干旱脅迫對植物生長及其生理的影響[J].江蘇林業科技,2012,39(4):51-54.
[3]中國科學院中國植物志編輯委員會.中國植物志 [M].北京:科學出版社,1984:54-55.
[4]陳江霞.八寶景天及應用領域[J].林木花卉,2009(11):46-47.
[5]李明啟.景天科酸代謝的碳途徑:植物代謝多條途徑觀點的一個典型例子[J].植物生理學通訊,1994,30(4):286-292.
[6]張澤錦.鐵皮石斛的光合碳同化途徑及其對環境變化的生理響應[D].北京:中國農業大學,2014.
[8]李云霞,張建生,吳永華,等.5種景天科地被植物抗旱性比較研究[J].干旱區資源與環境,2010,24(2):183-186.
[9]張斌,胡永紅,劉慶華,等.幾種屋頂綠化景天植物的耐旱性研究[J].中國農學通報,2008,24(5):272-276.
[10]李濱勝,周玉遷,潘杰,等.干旱脅迫下細葉景天生理生化指標的變化[J].北方園藝,2010(16):105-107.
[11]張立榮,牛海山,汪詩平,等.增溫與放牧對矮嵩草草甸4種植物氣孔密度和氣孔長度的影響 [J].生態學報,2010,30(24):6961-6969.
[12]徐萍,李進,呂海英,等.干旱脅迫下水楊酸對銀沙槐子葉表皮氣孔開度的影響[J].植物生理學報,2014,50(4):510-518.
[13]李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000:164-165,260-261.
[14]李強,唐薇,石園園,等.蒽酮-硫酸法和3-5-二硝基水楊酸法測定杜仲水提液多糖含量 [J].食品工業科技,2010(10):370-371,374.
[15]張歲岐,李金虎,山侖.干旱下植物氣孔運動的調控[J].西北植物學報,2011,21(6):1263-1270.
[16]于海秋,王曉磊,蔣春姬,等.土壤干旱下玉米幼苗解剖結構的傷害進程[J].干旱地區農業研究,2008,26(5):143-147.
[17]楊惠敏,王根軒.干旱和CO2濃度升高對干旱區春小麥氣孔密度及分布的影響[J].植物生態學報,2001,25(3):312-316.
[18]胡學儉,孫明高,夏陽,等.NaCl脅迫對無花果與海棠膜脂過氧化作用及保護酶活性的影響 [J].西北植物學報,2005,25(5):937-943.
[19]邵艷軍,山侖.植物耐旱機制研究進展[J].中國生態農業學報,2006,14(4):17-20.
[20]徐秀梅,張新華,王漢杰.四翅濱藜抗旱生理特性研究[J].南京林業大學學報(自然科學版),2004,28(5):54-58.
[21]牟鳳娟,陳麗萍,李軍萍,等.干旱脅迫下旱地油杉生理生化指標動態變化[J].干旱區資源與環境,2016,30(5):180-184.
[22] KLEIN T,HOCH G,YAKIR D,et al.Drought stress,growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest[J].Tree Physiology,2014,34(9):981-992.
[23] O'BREIN M J,LEUZINGER S,PHILIPSON C D,et al.Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels[J].Nature Climate Change,2014,4(8):710-714.
[24]叢建鷗,李寧,許映軍,等.干旱脅迫下冬小麥產量結構與生長、生理、光譜指標的關系 [J].中國生態農業學報,2010,18(1):67-71.
[25]何曉瑩.初花期干旱對油菜農藝性狀的影響 [J].山西農業科學,2016,44(9):1291-1293.
[26]王春乙,婁秀榮,王建林.中國農業氣象災害對作物產量的影響[J].自然災害學報,2007,16(5):38-43.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
今日農業(2021年11期)2021-08-13 08:53:34
當代陜西(2021年2期)2021-03-29 07:41:24
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
海峽姐妹(2016年5期)2016-02-27 15:20:20
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
