鉛脅迫對金絲草生長及生理生化的影響
2018-04-18 07:48:40韓航陳順鈺薛凌云侯曉龍蔡麗平劉愛琴周垂帆
草業學報 2018年4期
關鍵詞:植物
韓航,陳順鈺,薛凌云,侯曉龍, 2, 3*,蔡麗平,3,劉愛琴,3,周垂帆,3
(1.福建農林大學林學院, 福建 福州350002;2.福建農林大學資源與環境學院,福建 福州350002;3.海峽兩岸紅壤區水土保持協同創新中心,福建 福州350002)
近年來,由于我國礦產資源大規模開采,富含重金屬污染物無序排放,農藥化肥大量使用等因素,導致土壤重金屬污染嚴重。據環境保護部和國土資源部聯合發布全國土壤污染狀況調查顯示,全國土壤總的點位超標率達16.1%,部分工礦業地區土壤重金屬超標范圍較大,土壤環境質量堪憂[1]。其中,鉛污染因其毒害作用強、影響范圍廣等而備受關注,成為當前急需解決的環境問題[2]。
土壤重金屬污染治理傳統的工程、化學等方法存在需擾動土壤、費用昂貴等缺點,難以大規模推廣應用。植物修復技術因其綠色環保、經濟、操作性強等特點,在土壤重金屬污染治理方面有廣闊應用前景[3-4],植物修復技術的應用前提是篩選出重金屬超富集植物。多年生禾本科植物金絲草(Pogonatherumcrinitum),又名落蘇、黃毛草、毛毛草,是課題組在福建尤溪鉛鋅礦區發現,并通過室內試驗驗證的一種Pb的超富集植物,可在Pb濃度高達17000 mg·kg-1的鉛鋅礦區正常生長,對Pb具有較強耐性[5]。金絲草的鉛耐性是其長期適應Pb脅迫環境的演化結果,其中可能存在特殊的適應機制。大量研究表明,抗氧化系統及滲透調節物質的調節響應很大程度影響著植物的拮抗能力[6]。黑麥草可通過提高SOD、POD活性和可溶性糖及脯氨酸含量,增強體內清除活性氧和滲透調節能力,降低鉛毒害[7]。旱柳在Pb處理(<600 mg·kg-1)下,則通過根長和生物量調節,同時誘導POD、CAT和SOD等酶活響應來適應土壤鉛逆境[8]。但目前對金絲草適應土壤鉛脅迫的機理尚不清楚,特別是抗氧化系統和滲透調節系統在適應過程中發揮的作用仍有待驗證。
鑒于此,以金絲草為研究對象,測定不同梯度鉛脅迫下金絲草形態、生物量、抗氧化系統、滲透調節物質等指標,比較金絲草葉片和根系響應差異,探討逆境下金絲草抗氧化系統及滲透調節物質的響應規律,以期為耐性植物逆境響應機理研究提供科學參考。
1 材料與方法
1.1 試驗材料
試驗于2016年5月展開,供試金絲草苗來自福建農林大學森環所,選取10 cm高、長勢一致的新生幼苗待用。基質采用洗凈河沙和黃心土,風干后過2 mm篩,按1∶3混合(參考砂壤土質地)。基本理化性質:pH 5.2,全碳、全氮、全磷、全鉀含量分別為:3.37、0.46、0.17和37.62 g·kg-1,速效磷、速效鉀含量分別為:0.44、38.19 mg·kg-1,重金屬Pb、Zn、Cu含量分別為:1.03、2.14和0.35 mg·kg-1。
1.2 試驗設計
試驗用盆直徑30 cm,高25 cm,基質填充8 kg。參考國家土壤質量標準(GB 15618-1995),設計4個Pb濃度,0、1000、2000和3000 mg·kg-1(文中分別以CK、Pb 1000、Pb 2000、Pb 3000表示)。采用(CH3COO)2Pb溶液進行施加,并鈍化30 d。移植金絲草前,一次性施加底肥,每kg土肥料用量N為100 mg、P2O2為80 mg,K2O為100 mg,施加形態為KH2PO4和尿素[9]。每組處理5個重復,每盆10株金絲草幼苗。試驗在玻璃溫室中進行,溫度(25±2) ℃,濕度65%~75%。試驗過程中,使用土壤溫濕度儀監測土壤水分情況,及時補充土壤水分,保持含水量在田間持水量的70%左右。脅迫試驗60 d時,取樣測定金絲草葉片和根系相關形態和生理指標。
1.3 測定方法
株高、葉片指標和生物量測定:在脅迫60 d收獲時,每盆隨機選取5株金絲草使用直尺測量株高,以金絲草最長葉尖計,計算均值后為一次重復,每個處理組共測定5盆;葉長、葉寬和葉面積采用北京Yaxin-1241葉面積儀測定,每盆隨機選取5株金絲草,選取金絲草自上至下第3~9片順序中任一葉片,共5片,其中葉長以葉身計算,葉寬以葉身中部最寬處計算,計算均值后為一次重復,每個處理組共測定5盆;收獲金絲草洗凈后,將地上部和根系分開,分別測定鮮重。然后105 ℃殺青30 min,75 ℃烘干至恒重后分別記錄地上和根系部分干重,并計算根冠比。
抗氧化酶與MDA含量、抗氧化能力、滲透調節物質測定:每盆金絲草收獲后,分別取葉片和根系,洗凈混勻后各稱取0.2 g金絲草葉片和根系,加入5 mL預冷的pH 7.0的磷酸緩沖液,研磨成勻漿,于4 ℃下10000 r·min-1冷凍離心20 min,取上清液放4 ℃冰箱冷藏待測。超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)和丙二醛(MDA)含量分別采用氮藍四唑法、愈創木酚法、紫外吸收法測定和硫代巴比妥酸法測定[10]。抗超氧陰離子自由基活力(anti superoxide anion radical activity, ASAFR)和總抗氧化能力(total antioxidant capacity, T-AOC)均使用南京建成生物工程研究所提供的試劑盒(準確稱取組織重量,按重量體積比加入9倍的PBS制成10%的組織勻漿,2500 r·min-1,離心10 min,分別按試劑盒步驟操作加入各試劑后,1 cm光徑,波長550和520 nm處分別測定吸光度得ASAFR、T-AOC);可溶性蛋白(soluble protein, SP)含量采用考馬斯亮藍法測定[11],可溶性糖(soluble sugar, SS)含量采用蒽酮比色法測定[11],脯氨酸(proline, Pro)含量采用茚三酮比色法測定[12]。
1.4 數據統計分析
數據均為5次重復試驗結果,使用SPSS 20.0進行處理,采用單因素方差分析(One-way ANOVA)對試驗數據進行分析比較。采用Origin 8.5軟件制圖。
2 結果與分析
2.1 鉛脅迫對金絲草生長的影響
由圖1可知,Pb 1000處理下,金絲草株高、葉長和葉寬較對照無顯著變化。隨脅迫濃度增加,金絲草株高、葉長、葉寬均低于對照,抑制效果顯著(P<0.05)。其中,Pb 2000和Pb 3000處理下金絲草株高分別降低了13.4%和26.8%,葉長分別降低22.6%和26.9%,葉寬分別降低23.3%和18.4%。此外,鉛處理下金絲草葉面積均顯著低于對照(P<0.05)。
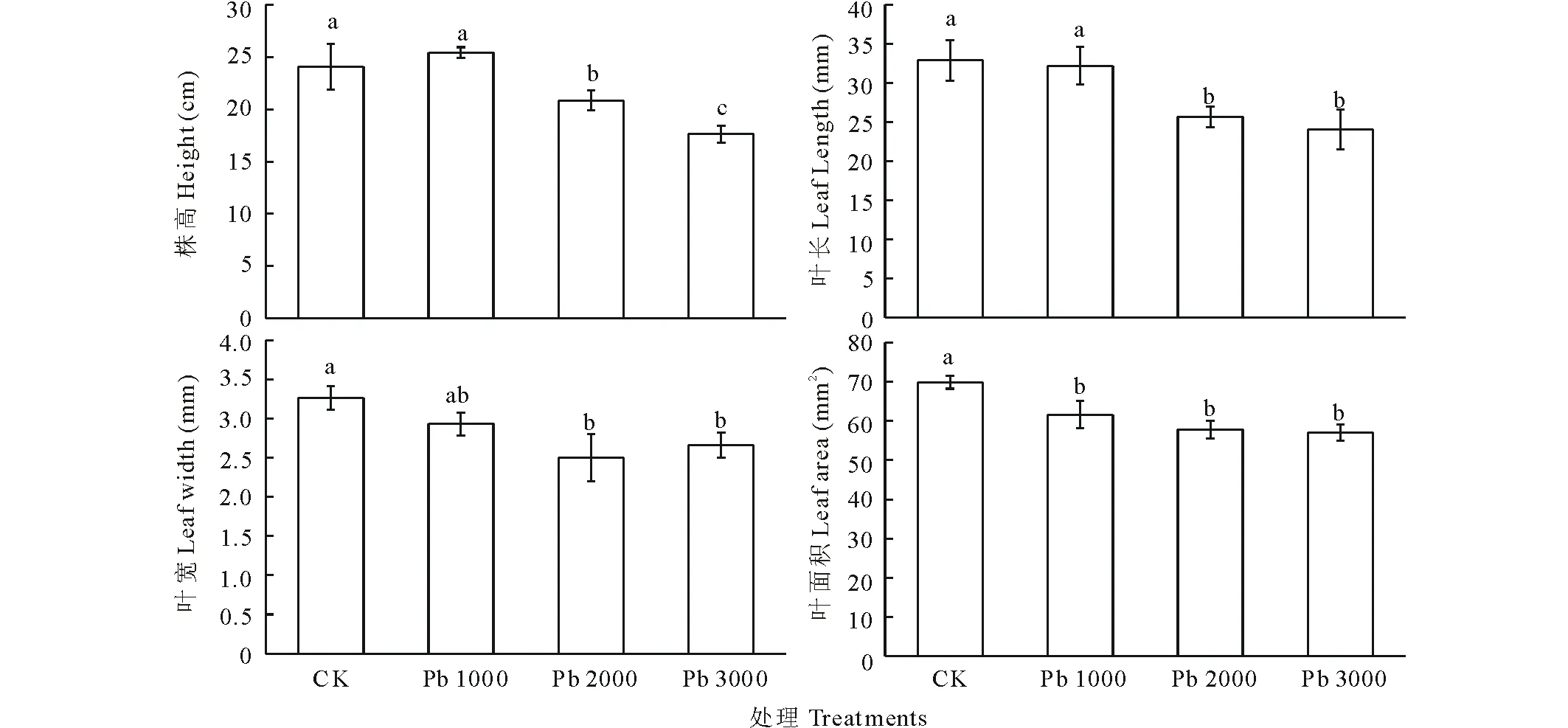
圖1 不同濃度鉛脅迫對金絲草地上部生長的影響Fig.1 The growth of aboveground parts of P. crinitum under Pb stress 不同小寫字母表示差異顯著(P<0.05),下同。The different letters mean the significant differences at P<0.05, the same below.
2.2 鉛脅迫對金絲草生物量的影響
由表1可知,低濃度鉛脅迫下(Pb 1000),金絲草地上和地下部分鮮重、干重及全株總質量均高于對照,其中地上部分干重增長13.4%,根干重增長14.7%。隨脅迫濃度增加,金絲草生物量逐漸受到抑制。Pb 3000處理下,金絲草地上和地下部鮮重、干重及全株質量均顯著低于對照(P<0.05),全株質量下降約62.8%。且鉛處理下金絲草根冠比均大于對照,金絲草根系生物量占比逐漸增加。綜上,金絲草生物量指標在鉛脅迫下總體呈“低促高抑”趨勢。
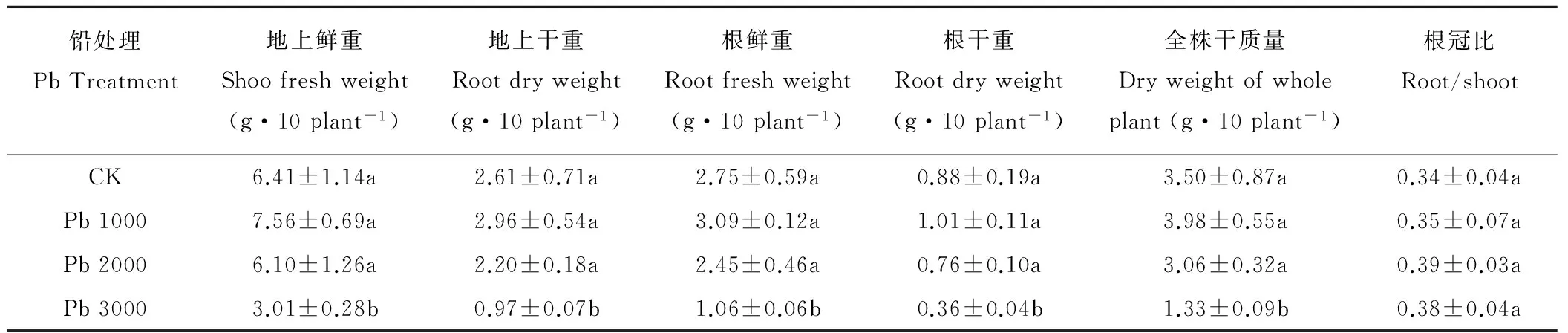
表1 不同濃度鉛脅迫對金絲草生物量的影響Table 1 The biomass of P. crinitum under Pb stress
注:同列不同小寫字母表示差異顯著(P<0.05),下同。
Note:The different letters within the same column mean the significant differences atP<0.05, the same below.
2.3 鉛脅迫對金絲草抗氧化酶活性的影響
由圖2(A)可知,鉛脅迫對金絲草葉片SOD活性抑制作用顯著(P<0.05),但脅迫濃度的增加對葉片SOD活性影響較小;金絲草根系SOD活性呈先增后減趨勢,Pb 2000處理下達最大值。且對照和Pb處理下金絲草葉片SOD活性均高于根系。由圖2(B)可知,鉛處理對金絲草葉片POD活性有一定促進作用,Pb 1000和Pb 2000處理下,葉片POD活性分別比對照增長17.9%和19.0%,但與對照無顯著差異(P>0.05);不同鉛脅迫處理金絲草根系POD活性均小于對照。同時,鉛脅迫條件下金絲草葉片POD活性均大于根系,而對照則相反。由圖2(C)可知,隨鉛脅迫濃度的增加,金絲草葉片和根系CAT活性均呈先減后增趨勢,Pb 1000和Pb 2000處理下,葉片和根系CAT活性低于對照,Pb 3000處理下較對照無顯著差異。此外,對照及Pb 1000處理下金絲草葉片CAT活性大于根系,而Pb 2000 和Pb 3000則相反。
2.4 鉛脅迫對金絲草抗氧化能力和MDA含量的影響
由圖3(A)可知,鉛脅迫下金絲草葉片抗超氧陰離子自由基活力(ASAFR)顯著低于對照(P<0.05),不同鉛處理間無顯著差異(P>0.05);金絲草根系在Pb 1000處理時高于對照,在Pb 3000處理時,顯著低于對照(P<0.05),呈先增后減趨勢。且除Pb 3000處理外,金絲草根系抗超氧陰離子自由基活力高于葉片。
植物總抗氧化能力(T-AOC)是植物體內酶促、非酶促等抗氧化物質總的抗氧化能力,受諸多因素影響,逆境下常表現為非線性、復雜性的規律[13]。由圖3(B)可知,土壤鉛脅迫對金絲草葉片總抗氧化能力(T-AOC)有一定促進作用,Pb 3000處理時比對照增長約76.3%,顯著高于對照及其他鉛處理(P<0.05);金絲草根系T-AOC在Pb 3000處理下顯著降低(P<0.05),Pb 1000和Pb 2000與對照無顯著差異(P>0.05);不同Pb處理和對照金絲草葉片T-AOC均大于根系。故推測,金絲草在Pb 1000~2000處理下,葉片和根系總抗氧化能力處于穩態,在Pb 3000處理下,由于葉片和根系抗氧化系統響應水平不同,導致根系T-AOC顯著下降,葉片則顯著增加。
由圖3(C)可知,鉛處理下葉片MDA含量呈增長趨勢,Pb 2000和Pb 3000處理時,顯著增加(P<0.05);根系MDA含量在鉛脅迫下均高于對照,其中Pb 1000處理顯著大于對照(P<0.05),而Pb 2000和Pb 3000與對照無顯著差異(P>0.05);對照及Pb處理下金絲草葉片MDA含量均高于根系。
2.5 鉛脅迫對金絲草滲透調節物質的影響
由圖4(A)可知,鉛處理下金絲草葉片可溶性蛋白含量均高于對照,Pb 2000處理下達最大值,較對照增加36.4%,顯著大于對照(P<0.05);金絲草根系可溶性蛋白含量則先減后增,在Pb 2000和Pb 3000處理高于對照;金絲草葉片可溶性蛋白含量在鉛處理及對照中均高于根系。由圖4(B)可知,金絲草葉片可溶性糖含量在鉛處理下呈先減后增趨勢,在Pb 3000處理時達最大值,顯著高于對照(P<0.05);Pb 1000和Pb 2000處理下金絲草根系可溶性糖含量分別比對照增加17.7%和19.0%,顯著增長(P<0.05),在Pb 3000處理下受明顯抑制。且葉片可溶性糖含量總體高于根系。由圖4(C)可知,金絲草葉片脯氨酸含量總體高于根系。鉛脅迫下金絲草葉片脯氨酸含量呈先減后增趨勢,在Pb 3000處理下達最大值,與對照無顯著差異(P<0.05);金絲草根系脯氨酸含量在Pb 1000處理時達最大值,隨鉛脅迫濃度增加,顯著降低(P<0.05)。

圖3 不同濃度鉛脅迫對金絲草抗氧化能力的影響Fig.3 Effect of Pb stress on the antioxidant capacity of P. crinitum
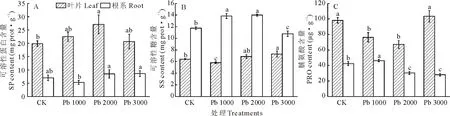
圖4 不同濃度鉛脅迫對金絲草滲透調節物質的影響Fig.4 Effect of Pb stress on the osmotic adjustment substances of P. crinitum
3 討論
植物生長形態和生物量變化是植物適應環境生長的最終結果。沈高峰[14]發現,鉛脅迫(≤1500 mg·kg-1)18 d時,高羊茅(Festucaarundinacea)和多年生黑麥草(Loliumperenne)鮮重分別為對照的67.44%和68.09%。李西等[15]也發現,土壤鉛脅迫會抑制假儉草(Eremochloaophiuroides)、狗牙根(Cynodondactylon)與結縷草(Zoysiajaponica)地上和根系生物量。本研究中,除Pb 1000處理外,其他鉛處理下金絲草株高、葉長、葉寬均顯著低于對照,抑制作用明顯,與前人研究基本一致。同時,金絲草地上與地下部分鮮重、干重及全株生物量在Pb 1000處理時達最大值。在Pb 2000處理時,與對照無顯著差異,在Pb 3000處理時,顯著降低,符合“低促高抑”規律。但鉛處理下金絲草根冠比逐漸增大,這可能是金絲草增強根系生物量占比,提高逆境下對養分、水分的汲取能力來主動適應鉛脅迫的結果。
重金屬鉛脅迫會抑制植物生長、導致植株矮化、葉片面積變小,降低植物鮮重和干重。同時,植物在鉛脅迫下體內會產生多種活性氧(ROS),造成膜脂質過氧化毒害、抑制酶活、削弱細胞正常滲透調節能力等[16]。為了避免和降低重金屬毒害作用,植物有一套復雜的防御機制。其中誘導抗氧化酶系統響應和提高滲透調節物質水平來清除植株體內活性氧自由基、維持細胞正常滲透壓和水勢,是耐性植物逆境生存的重要手段[17]。SOD、POD、CAT是植物抗氧化系統重要組成部分,具有清除O2-·、H2O2的作用,逆境下不同植物抗氧化酶響應敏感程度和響應規律略有不同[18]。有研究發現,香根草苗在水培Pb脅迫下,隨Pb2+濃度的增加(0~8 mmol·L-1),香根草苗SOD活性顯著增加,POD和CAT活性則在高濃度(>4 mmol·L-1)Pb處理時,逐漸受到抑制[19]。灰綠藜和小藜幼苗SOD及POD活性則分別在200、100 mg·kg-1鉛濃度下達到最高,CAT的活性則在100 mg·kg-1鉛濃度時達到最大[20]。本研究中,除金絲草葉片SOD活性和根系POD活性受Pb脅迫明顯抑制外,金絲草葉片和根系其他酶活并未表現出單一增減規律。如金絲草根系SOD活性無顯著變化,葉片POD活性趨勢為先增后減。這可能是在一定閾值內鉛脅迫對金絲草抗氧化酶活性有刺激作用,但隨脅迫濃度增加,金絲草酶活系統受損,酶活降低,與李錚錚等[21]結論類似。且同一濃度鉛處理下,金絲草葉片與根系酶活響應水平不同,Pb 1000~3000處理下金絲草葉片SOD、POD活性均高于根系,推測是金絲草根系與污染土壤直接接觸,根系生長受阻、根系過氧化作用對酶活抑制更強造成的。綜上,金絲草主要通過葉片和根系抗氧化酶差異化響應,提高葉片和根系CAT活性,維持根系SOD活性、葉片POD活性穩態,協同配合來降低過氧化毒害。
另一方面,抗超氧陰離子自由基活力、總抗氧化能力和MDA含量高低反映了Pb脅迫下植物細胞膜脂過氧化程度,也體現出植物體內各類抗氧化物質總的抗氧化能力[13]。本研究中,隨鉛處理濃度增加,金絲草葉片和根系抗超氧陰離子自由基活力顯著低于對照(除Pb 1000處理下金絲草根系外),金絲草葉片和根系MDA含量均高于對照,鉛脅迫毒害漸顯。但金絲草葉片和根系T-AOC在Pb 1000~2000處理下處于穩態,葉片T-AOC在Pb 3000處理下顯著增長。這表明鉛脅迫在導致金絲草葉片和根系膜脂過氧化損傷的同時,也會誘導金絲草體內抗氧化酶、抗氧化物等抵御機制響應,使得金絲草葉片和根系總抗氧化能力處于高位水平,增強了金絲草整體拮抗能力。
此外,可溶性蛋白、可溶性糖和脯氨酸是植物體內重要的滲透調節物質,具有保持逆境下植物體正常含水量和膨壓的功能,是植物抗氧化系統中非酶物質重要的組成部分[22-23]。土壤鉛脅迫可能誘導了金絲草細胞內部基因表達等復雜機制的響應,使得鉛處理下葉片和根系的可溶性蛋白、可溶性糖含量處于較高水平,間接促進了金絲草體內活性氧清除,平衡了細胞質與液泡間滲透勢,使得金絲草體內細胞代謝等生理活動可正常運作,這可能是金絲草主要的抗逆機制之一。
4 結論
金絲草對土壤鉛脅迫有較強耐性,金絲草通過提高葉片POD活性、根系SOD活性等不同器官抗氧化酶差異化響應,促進金絲草葉片和根系可溶性蛋白和可溶性糖含量增加,維持金絲草體內總抗氧化能力處于較高水平,增強了金絲草抵御過氧化毒害和細胞逆境滲透調節能力,較好地適應了土壤鉛脅迫。但本研究同時發現,在Pb 3000脅迫下金絲草POD和可溶性蛋白等指標降低,MDA含量有增加趨勢,說明不同抗氧化酶響應Pb脅迫的作用有一定閾值,金絲草適應高濃度Pb脅迫可能還存在除抗氧化系統外的其他機理。
參考文獻References:
[1] Wang Y J, Liu C, Zhou D M,etal. A critical view on the status quo of the farmland soil environmental quality in China: discussion and suggestion of relevant issues on Report on the national general survey of soil contamination. Journal of Agro-Environment Science, 2014, 33(8): 1465-1473.
王玉軍, 劉存, 周東美, 等. 客觀地看待我國耕地土壤環境質量的現狀——關于《全國土壤污染狀況調查公報》中有關問題的討論和建議. 農業環境科學學報, 2014, 33(8): 1465-1473.
[2] Nemati K, Abu Bakar N K, Abas M R,etal. Speciation of heavy metals by modified BCR sequential extraction procedure in different depths of sediments from Sungai Buloh, Selangor, Malaysia. Journal of Hazardous Materials, 2011, 192(1): 402-410.
[3] Sánchez-Chardi A. Biomonitoring potential of five sympatric Tillandsia, species for evaluating urban metal pollution (Cd, Hg and Pb). Atmospheric Environment, 2016, 131: 352-359.
[4] Hou X L, Cai L P, Han H,etal. Effect of lead stress on the chorophyll fluorescence characteristics and antioxidative enzyme activities ofPaspalumnotatum. Acta Prataculturae Sinica, 2017, 26(3): 142-148.
侯曉龍, 蔡麗平, 韓航, 等. 鉛脅迫對百喜草葉綠素熒光特性及酶活性的影響. 草業學報, 2017, 26(3): 142-148.
[5] Hou X L, Chang Q S, Liu G F,etal. Two lead-hyperaccumulator:PogonatherumcrinitumandLsacheglobosa. Chinese Journal of Environmental Engineering, 2012, 6(3): 989-994.
侯曉龍, 常青山, 劉國鋒, 等. Pb超富集植物金絲草 (Pogonatherumcrinitum), 柳葉箬(Lsacheglobosa). 環境工程學報, 2012, 6(3): 989-994.
[6] Mahar A, Wang P, Ali A,etal. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: A review. Ecotoxicology & Environmental Safety, 2016, 126: 111-121.
[7] Liu H Q, Han J C, Liu H P,etal. Influence of lead gradient stress on the physiological and biochemical characteristics of perennial ryegrass (Loliumperenne) seedlings. Acta Prataculturae Sinica, 2012, 21(6): 57-63.
劉慧芹, 韓巨才, 劉慧平, 等. 鉛梯度脅迫對多年生黑麥草幼苗生理生化特性影響. 草業學報, 2012, 21(6): 57-63.
[8] Zhu J, Wang P, Jia S S,etal. Tolerance, accumulation, translocation and stress response ofSalixmatsudanaKoidz to lead. Acta Scientiae Circumstantiae, 2016, 36(10): 3876-3886.
朱健, 王平, 夾書珊, 等. 旱柳(SalixmatsudanaKoidz)對Pb的耐性、富集、轉運與脅迫響應研究. 環境科學學報, 2016, 36(10): 3876-3886.
[9] Sun J Y. Varietal difference in Cd tolerance and mechanism in cabbage. Nanjing: Nanjing Agricultural University, 2011.
孫建云. 甘藍(BrassicaoleraceaL.)耐鎘性的品種差異及其機理研究. 南京: 南京農業大學, 2011.
[10] Zou Q. Experimental guidance of plant physiology and biochemistry. Beijing: China Agriculture Press, 1995.
鄒琦. 植物生理生化實驗指導. 北京: 中國農業出版社, 1995.
[11] Li H S. Plant physiological and biochemical experimental principles and techniques. Beijing: Higher Education Press, 2000.
李合生. 植物生理生化實驗原理和技術. 北京: 高等教育出版社, 2000.
[12] Wang X K, Huang J L. Principles and techniques of plant physiology and biochemistry experiment. Beijing: Higher Education Press, 2015.
王學奎, 黃見良. 植物生理生化實驗原理與技術. 北京: 高等教育出版社, 2015.
[13] Gül?in. Antioxidant activity of food constituents: an overview. Archives of Toxicology, 2012, 86(3): 345-391.
[14] Shen G F. Effects of simulated acid rain and Pb combined stress on growth and antioxidative physiology of two kinds of turf grass. Journal of Anhui Agricultural Sciences. 2012, 40(22): 11283-11286.
沈高峰. 模擬酸雨和Pb復合脅迫對2種草坪草生長及抗氧化生理的影響. 安徽農業科學, 2012, 40(22): 11283-11286.
[15] Li X, Wu Y J, Sun L X,etal. Growth and physiological responses of three warm-season turfgrasses to lead stress. Acta Prataculturae Sinica, 2014, 23(4): 171-180.
李西, 吳亞嬌, 孫凌霞. 鉛脅迫對三種暖季型草坪草生長和生理特性的影響. 草業學報, 2014, 23(4): 171-180.
[16] Bankaji I, Ca?ador I, Sleimi N. Physiological and biochemical responses ofSuaedafruticosato cadmium and copper stresses: growth, nutrient uptake, antioxidant enzymes, phytochelatin, and glutathione levels. Environmental Science and Pollution Research, 2015, 22(17): 13058-13069.
[17] Tian B H, Zhang Y J, Zhang L P,etal. Effects of cadmium or chromium on growth and NADPH oxidase and antioxidant enzyme system of foxtail millet seedlings. Journal of Agro-Environment Science, 2016, 35(2): 240-246.
田保華, 張彥潔, 張麗萍, 等. 鎘/鉻脅迫對谷子幼苗生長和NADPH氧化酶及抗氧化酶體系的影響. 農業環境科學學報, 2016, 35(2): 240-246.
[18] Chen J W. Study on response and potential phytoremediation ofBidenspilosaL. in cadmium and lead stress. Chongqing: Southwest University, 2013.
諶金吾. 三葉鬼針草(BidenspilosaL.)對重金屬Cd、Pb脅迫的響應與修復潛能研究. 重慶: 西南大學, 2013.
[19] Han L, Zhang X P, Liu B R,etal. Response ofVetiveriazizanioidesto Pb2+stress. Chinese Journal of Applied Ecology, 2005, 16(11): 2178-2181.
韓露, 張小平, 劉必融. 香根草對重金屬鉛離子的脅迫反應研究. 應用生態學報, 2005, 16(11): 2178-2181.
[20] Liu J H, Yao Z G. Effects of Pb stress on seed germination and seedling growth of two weed species seeds. Agricultural Science & Technology, 2016, 17(12): 2726-2731.
劉俊華, 姚志剛. 重金屬鉛脅迫對2種雜草種子萌發和幼苗生長的影響. 農業科學與技術 (英文版), 2016, 17(12): 2726-2731.
[21] Li Z Z, Wu J, Tang Y,etal. Effect of Pb, Zn and their interactions on the chlorophyll content and antioxidant enzyme systems ofHouttuyniacordatathumb. Acta Ecologica Sinica, 2007, 27(12): 5441-5445.
李錚錚, 伍鈞, 唐亞, 等. 鉛、鋅及其交互作用對魚腥草(Houttuyniacordata)葉綠素含量及抗氧化酶系統的影響. 生態學報, 2007, 27(12): 5441-5446.
[22] Marcu D, Damian G, Cosma C,etal. Gamma radiation impact on lettuce (Lactucasativavar.Capitata) growth and development, mitotic cell division and electron spin resonance analysis of induced free radicals. Environmental Engineering & Management Journal, 2014, DOI:10.1007/s10867-013-9322-z.
[23] Zhang J Y. The effect of Pb and Cd on physiological and biochemical indexes ofMonsteradeliciosaLiebm. in the short-term conditions. Journal of Soil and Water Conservation, 2016, 30(2): 340-345.
張家洋. 重金屬鉛鎘短期脅迫對蓬萊蕉生理生化指標的影響. 水土保持學報, 2016, 30(2): 340-345.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
