抗壞血酸引發對NaCl脅迫燕麥種子活力的影響
2018-04-18 07:51:18董秋麗夏方山李曉禹王明亞毛培勝朱慧森佟莉蓉杜利霞
草業學報 2018年4期
關鍵詞:生長
董秋麗,夏方山,李曉禹,王明亞,毛培勝,朱慧森,佟莉蓉,杜利霞
(1.山西農業大學林學院,山西 太谷 030801;2.山西農業大學動物科技學院,山西 太谷 030801; 3.中國農業大學動物科技學院,草業科學北京市重點實驗室,北京 100193)
燕麥(Avenasativa)具有耐瘠薄及抗逆性強等優點,因其產草量高、營養價值高及適口性好,已成為解決草地畜牧業生產中冷季缺草問題的重要飼草料來源[1]。此外,燕麥籽實因富含可溶性膳食纖維β-葡聚糖、均衡蛋白質及油脂等對人類健康至關重要的營養成分,是對人類具有綠色營養保健作用的重要小雜糧作物[2]。所以,燕麥成為我國北方地區廣泛種植的優質高產糧飼兼用作物,其播種面積和總產量僅次于小麥(Triticumaestivum)、玉米(Zeamays)及水稻(Oryzasativa)[3-5]。因此,探討燕麥高效種植途徑對于農牧業可持續發展及解決飼糧安全問題具有重要意義。
耐鹽植物品種選育被認為是有效抑制土壤鹽漬化,并科學利用鹽堿地資源最有效、最經濟的途徑之一[6]。燕麥被認為是改良鹽漬化土壤的主要先鋒作物之一,種植于含鹽量0.2%的土壤上能有效提高其出苗和產量[7-8]。但土壤鹽漬化依然是制約燕麥生產的關鍵因素,尤其春季返鹽季節對其種子萌發及幼苗生長發育的損害最大[9]。此外,由于燕麥籽實不飽和脂肪酸含量達80%以上,而脂肪衍生物容易酸敗或劣變,致使其種子萌發過程更容易遭受逆境損傷,從而降低其種用價值[2]。因此,探究如何提高燕麥種子萌發及其幼苗生長能力是其充分利用并改良鹽漬化土地的最有效途徑。引發是指控制種子的水合狀態,使其避免胚根突破種皮而足以發生萌發前代謝,從而改善種子在不同生態條件下的活力水平及幼苗生長潛能[10]。利用適當的小分子生理活性物質對種子進行引發處理,不僅能啟動其萌發前的代謝反應,還能參與其萌發過程的代謝反應[11]。這既能有效地增強種子活力及幼苗的抗逆性,又有助于其市場流通和應用于農牧業生產。因此,如何采用生理活性物質引發提高種子活力及其抗逆性,是當下種子科學領域研究的主要熱點問題之一。抗壞血酸(ascorbic acid,AsA)作為種胚細胞內普遍存在的重要小分子生理活性物質,其生物學功能的研究日益受到種子科學的重視[12]。AsA可參與種胚細胞內許多氧化還原反應及物質代謝活動,從而維持其活力水平及促進其細胞分裂與伸長[11,13]。外源AsA提高種子耐鹽性的研究已在菜豆(Phaseolusvulgaris)[14]、甘蔗(Saccharum)[15]、鷹嘴豆(Cicerarietinum)[16]、小麥[17]及油菜(Brassicacampestris)[18]等植物中報道。然而,關于外源AsA引發對鹽脅迫種子活力的影響研究較少。因此,試驗以燕麥種子為材料,探索AsA引發對其鹽脅迫條件下萌發的活力影響,為外源AsA引發促進植物種子耐鹽性萌發機理的研究提供參考依據。
1 材料與方法
1.1 材料來源
供試燕麥(品種:太陽神)種子由北京正道生態科技有限公司提供,2016年進口于惠特蘭種子有限責任公司(Wheatland Seed LLC),在-20 ℃保存至進行試驗。種子自然含水量為7.8%,初始發芽率為100%。
1.2 含水量的測定
準確稱取4.5 g燕麥種子,放入樣品盒中稱重(精確到0.001 g),設2次重復。稱取后保持樣品盒開啟,放入130~133 ℃烘箱內烘干1 h,烘干后取出,蓋好盒蓋,放入干燥器里冷卻30 min,按公式計算含水量:種子含水量=[(M2-M3)/(M2-M1)]×100%,式中:M1為樣品盒和蓋的重量(g);M2為樣品盒、蓋及樣品的烘前重量(g);M3為樣品盒、蓋及樣品的烘后重量(g)。
1.3 AsA引發處理
在20 ℃黑暗條件下,將燕麥種子分別用濃度為0、0.5、1.0、1.5和2.0 mmol·L-1的AsA引發處理0.5 h后,蒸餾水沖洗2次,用濾紙吸干表層水分,然后25 ℃室內風干至含水量為10%,每個處理重復4次。
1.4 發芽試驗及指標測定
選取均勻飽滿的處理種子100粒放入培養皿中,分別加入8 mL濃度為0(CK)、50、100和200 mmol·L-1的NaCl溶液,在20 ℃恒溫條件下培養,每天更換NaCl溶液,設4次重復。發芽條件參照國際種子檢驗協會(International Seed Testing Association,ISTA)的種子檢驗規程(2015)[19]規定進行。每天統計種子發芽數,以第10天為末次計數,并最終測定其平均苗長,發芽率(germination percentage,Gp)按照ISTA的種子檢驗規程(2015)[19]進行計算,發芽指數(germination index,Gi)按照Abdul-Baki等[20]的方法進行計算,平均發芽時間(mean germination time,MGT)按照Ellis等[21]的方法進行計算。
1.5 統計分析
通過Excel 2010和SAS 8.0統計分析軟件處理試驗數據,采用Duncans法進行多重比較,結果以平均值±標準誤表示。
2 結果與分析
2.1 AsA引發對NaCl脅迫燕麥種子發芽率(Gp)的影響
隨NaCl濃度的升高,AsA濃度為1.5 mmol·L-1時,燕麥種子Gp呈先升后降的趨勢,在NaCl濃度為50 mmol·L-1時顯著(P<0.05)高于其濃度為0(CK)時(表1);AsA濃度為0、0.5、1.0和2.0 mmol·L-1時,燕麥種子Gp在NaCl濃度為0(CK)和50 mmol·L-1時差異不顯著(P>0.05),而在其濃度為100和200 mmol·L-1時顯著(P<0.05)下降。隨AsA濃度的升高,燕麥種子Gp在NaCl濃度為0(CK)和50 mmol·L-1時差異不顯著(P>0.05);在NaCl濃度為100和200 mmol·L-1時呈先升后降的趨勢,均在AsA濃度為1.0 mmol·L-1時最高。
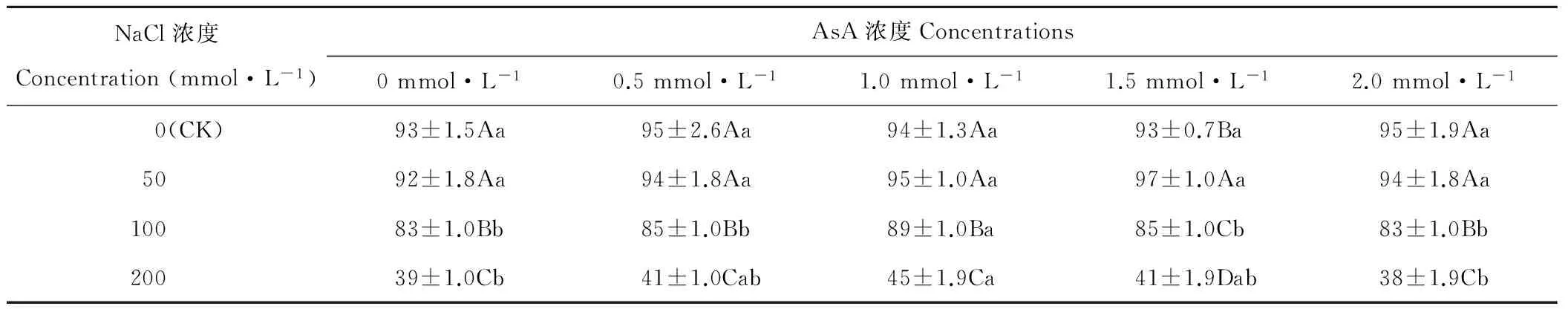
表1 AsA引發對NaCl脅迫燕麥種子發芽率的影響Table 1 Effect of AsA treatments on germination percentage in oat seeds under NaCl stress (%)
注:同列不同大寫字母表示差異顯著(P<0.05),同行不同小寫字母表示差異顯著(P<0.05)。下同。
Note: Means in the same column with different capital letters are significant differences (P<0.05), in the same row with different small letters are significant differences (P<0.05). The same below.
2.2 AsA引發對NaCl脅迫燕麥種子發芽指數(Gi)的影響
隨NaCl濃度的升高,燕麥種子Gi逐漸下降(表2),但在NaCl濃度為0(CK)和50 mmol·L-1時差異不顯著(P>0.05)。隨AsA濃度的升高,燕麥種子Gi在NaCl濃度為0(CK)和50 mmol·L-1時呈下降趨勢;在NaCl濃度為100和200 mmol·L-1時呈先升后降的趨勢,均在AsA濃度為2.0時顯著(P<0.05)低于其他濃度;然而,NaCl濃度為100 mmol·L-1時,燕麥種子Gi在AsA濃度為0.5 mmol·L-1時顯著(P<0.05)高于1.0~2.0 mmol·L-1,但與其濃度為0時差異不顯著(P>0.05);NaCl濃度為200 mmol·L-1時,燕麥種子Gi在AsA濃度為1.0 mmol·L-1時顯著(P<0.05)高于其他濃度。
2.3 AsA引發對NaCl脅迫燕麥種子平均發芽時間(MGT)的影響
隨NaCl濃度的升高,燕麥種子MGT逐漸增加,除AsA濃度為0和2.0 mmol·L-1時,其MGT在NaCl濃度為0(CK)和50 mmol·L-1時差異不顯著(P>0.05)外,其他條件下均差異顯著(P<0.05)(表3)。NaCl濃度為0 mmol·L-1時,燕麥種子MGT隨AsA濃度的增加先降后升,并在AsA濃度為0.5 mmol·L-1時顯著(P<0.05)低于其他濃度;NaCl濃度為50、100和200 mmol·L-1時,燕麥種子MGT隨AsA濃度增加呈增加趨勢,2.0 mmol·L-1時顯著高于0~1.0 mmol·L-1。

表2 AsA處理對NaCl脅迫燕麥種子發芽指數的影響Table 2 Effect of AsA treatmens on germination index in oat seeds under NaCl stress
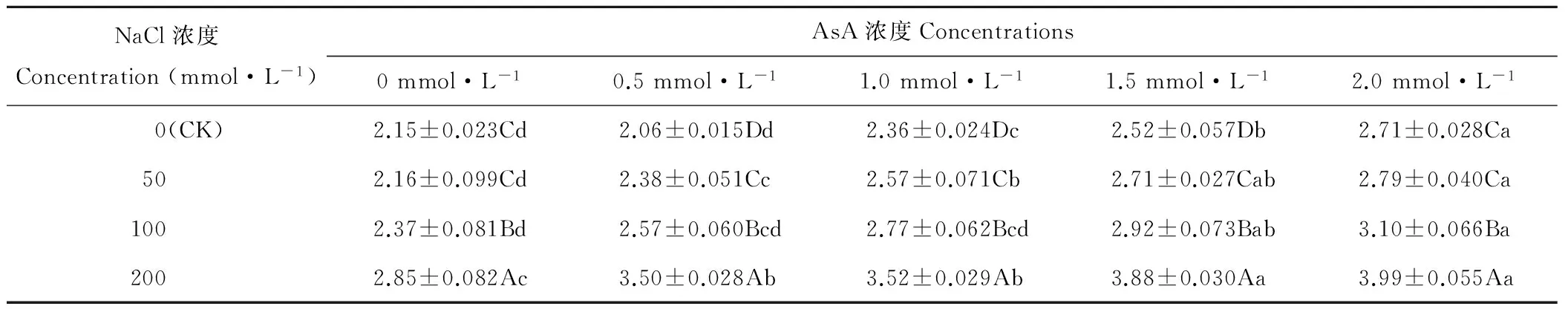
表3 AsA處理對NaCl脅迫燕麥種子平均發芽時間的影響Table 3 Effect of AsA treatments on mean germination time of oat seeds under stress (d)
2.4 AsA濃度影響NaCl脅迫下燕麥種子萌發的雙因素方差分析
雙因素方差分析結果表明(表4),不同NaCl濃度及AsA引發濃度對燕麥種子Gp、Gi和MGT的影響差異極顯著(P<0.01),不同NaCl濃度與AsA引發濃度的交互作用對燕麥種子Gp的影響差異顯著(P<0.05),對其Gi和MGT的影響均差異極顯著(P<0.01)。
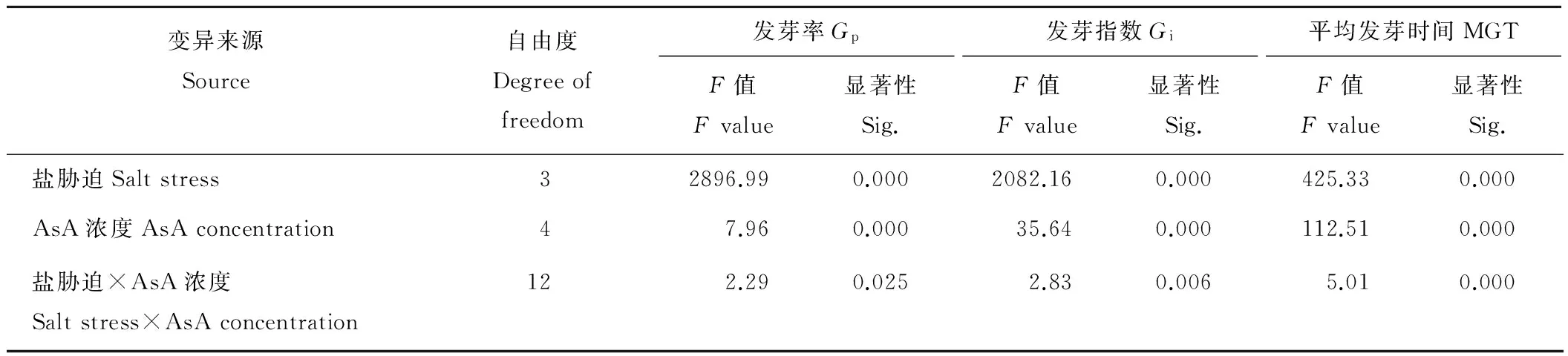
表4 AsA濃度和鹽脅迫對燕麥種子活力影響的雙因素方差分析Table 4 Variance analysis of AsA concentration and salt stress on oat seed vigor
Gp: Germination percentage;Gi: Germination index; MGT: Mean germination time.
3 討論
鹽脅迫是限制植物生長發育最主要、最常見的非生物脅迫,導致其產量和品質的下降,從而給農牧業生產造成巨大經濟損失。鹽脅迫不僅會抑制燕麥種子的萌發及幼苗生長,還會造成其產量及品質的降低[5,22]。本試驗中,與CK相比,燕麥種子Gp、Gi在50 mmol·L-1的NaCl脅迫下變化不顯著(P>0.05),其MGT顯著(P<0.05)增大;在100和200 mmol·L-1的NaCl脅迫下,燕麥種子Gp和Gi均顯著(P<0.05)下降,而其MGT則顯著(P<0.05)增大。這與前人的研究結果相似,即NaCl脅迫會降低燕麥種子的活力水平。劉鳳岐等[5]研究發現,NaCl脅迫顯著(P<0.05)降低了燕麥種子Gp、發芽勢、株高和根長,且這種抑制現象隨NaCl濃度的增大而增加。此外,復合鹽堿脅迫下也發現了燕麥種子Gp、發芽勢、胚根及胚芽生長的降低[23]。這說明燕麥種子活力下降是限制其在鹽堿地種植的關鍵因素。
AsA作為植物體內普遍存在的高豐度小分子生理活性物質,既能維持種子的活力水平,又能促進其萌發及幼苗生長[24]。此外,高含量的AsA會激活植物根系分生組織細胞的分裂,外源AsA處理促進植物根系的生長[25]。然而,本試驗中,在NaCl濃度為0 mmol·L-1(CK)時,燕麥種子Gp隨AsA濃度的增加變化不顯著(P>0.05),但其Gi呈下降趨勢,并在AsA濃度為1.5和2.0 mmol·L-1時顯著(P<0.05)低于0和0.5 mmol·L-1,且其MGT在AsA濃度為1.0~2.0 mmol·L-1時顯著(P<0.05)高于0和0.5 mmol·L-1。這說明在正常條件下,AsA引發導致了燕麥種子活力的下降。這可能與燕麥種子本身活力水平較高(初始發芽率為100%)有關,種子在正常萌發過程中AsA含量及相關抗氧化酶活性也會明顯升高[26]。然而,AsA濃度的增加改變了其引發溶液的水勢,從而造成燕麥種子內滲透調節物質的含量發生了變化,導致其在引發過程遭受滲透損傷[27]。因此,在農牧業生產中對AsA引發的應用要根據種子本身活力水平及萌發條件而定。
植物體內AsA含量高低直接決定了其抗逆性的強弱[28]。因此,外源AsA處理可有效促進植物種子的耐鹽性萌發及幼苗生長,這已在紫花苜蓿(Medicagosativa)[29]、黃芩(Scutellariabaicalensis)[30]及油菜(Brassicacampestris)[31]等植物種子中被驗證。本試驗結果表明,隨AsA濃度增加,燕麥種子Gp在NaCl濃度為100和200 mmol·L-1時呈先升后降的趨勢,這說明AsA引發能夠提高燕麥種子的耐鹽性萌發。NaCl脅迫會導致植物種子內H2O2快速產生,而外源AsA可以增強酶促或非酶促抗氧化途徑的作用來有效清除鹽脅迫產生的H2O2,從而提高植物種子耐鹽性萌發及幼苗生長[32-34]。然而,外源AsA引發提高燕麥種子耐鹽性萌發及其幼苗生長的機理是否與此相同仍需進一步深入探討。本試驗中,燕麥種子Gp均在AsA濃度為1.0 mmol·L-1時顯著大于其他濃度,且不同濃度AsA引發濃度對燕麥種子Gp、Gi和MTG的影響差異極顯著(P<0.01),這說明AsA引發對NaCl脅迫的緩解作用與其濃度有關,以1.0 mmol·L-1的AsA引發對NaCl脅迫的緩解效果最好。江緒文等[30]研究發現適宜濃度(0.50 mmol·L-1)的AsA能提高種子的萌發能力和幼苗對鹽脅迫的適應能力,從而起到緩解鹽脅迫對種子萌發及幼苗生長的抑制作用。范美華等[31]也發現,20 mg·L-1的AsA處理對30%海水脅迫下油菜種子萌發及幼苗生長的緩解效果最好。外源AsA濃度的增加也會造成燕麥種子內滲透勢的增強。同時,外源AsA會導致NaCl脅迫種子內可溶性糖和脯氨酸等滲透調節物質含量增加[30-31]。因此,高濃度AsA會導致NaCl脅迫燕麥種子的萌發及幼苗生長能力的下降。
然而,植物種子耐鹽性萌發及幼苗生長的增強不僅與外源AsA濃度有關,還與其遭受的鹽脅迫強度有關。NaCl脅迫程度越重,植物種子內H2O2積累就越多[33-34]。因此,維持種子內H2O2平衡所需的AsA就越多。試驗中,燕麥種子Gp在NaCl濃度為50 mmol·L-1時變化不顯著,而在NaCl濃度為100和200 mmol·L-1時呈先升后降的趨勢;且不同鹽脅迫及其與AsA引發濃度之間的交互作用對燕麥種子Gp、Gi和MTG的影響差異極顯著(P<0.01)。因此,在農牧業生產實踐中應根據燕麥種子本身活力及鹽脅迫的程度采用最佳濃度的AsA進行引發處理。本試驗中,隨AsA濃度增加,燕麥種子Gi呈下降趨勢,而其MGT呈上升趨勢,說明AsA引發延長了燕麥種子萌發的時間,這可能由于外源AsA在清除NaCl脅迫過程產生的H2O2需要反應時間,而且所需的反應時間與AsA及NaCl濃度關系密切。
4 結論
外源AsA引發可提高燕麥種子在NaCl脅迫下的萌發及幼苗生長能力,且這種作用效果與AsA及NaCl的濃度有關,以1.0 mmol·L-1的AsA引發對100和200 mmol·L-1的NaCl脅迫效果最好。然而,外源AsA引發延長了NaCl脅迫下燕麥種子的MGT。
參考文獻References:
[1] Gao C T, Liu J H, Zhang Y Q,etal. physiological responses of oats seedling to short-term salt stress. Acta Agrestia Sinica, 2017, 25(2): 337-343.
高彩婷, 劉景輝, 張玉芹, 等. 短期鹽脅迫下燕麥幼苗的生理響應. 草地學報, 2017, 25(2): 337-343.
[2] Claudine C, Tom S, Susan R V,etal. Comparison of two headspace sampling techniques for the analysis of off-flavour volatiles from oat based products. Food Chemistry, 2012, 134(3): 1592-1600.
[3] Shewry P R, Piironen V, Lampi A M,etal. Phytochemical and fiber components in oat varieties in the HEALTHGRAIN diversity screen. Journal of Agricultural and Food Chemistry, 2008, 56(21): 9777-9784.
[4] Guo H Y, Jia J Q, Lü J H,etal. Analyses of genetic diversities and evolutions ofAvenaL. germplasm resources by ISSR. Acta Agrestia Sinica, 2014, 22(2): 344-351.
郭紅媛, 賈舉慶, 呂晉慧, 等. 燕麥屬種質資源遺傳多樣性及遺傳演化關系ISSR分析. 草地學報, 2014, 22(2): 344-351.
[5] Liu F Q, Liu J L, Zhu R F,etal. Physiological responses and tolerance of four oat varieties to salt stress. Acta Prataculturae Sinica, 2015, 24(1): 183-189.
劉鳳歧, 劉杰淋, 朱瑞芬, 等. 4種燕麥對NaCl脅迫的生理響應及耐鹽性評價. 草業學報, 2015, 24(1): 183-189.
[6] Dai W, Qiu G J, Shi Y G,etal. Effects of salt stress on growth and physiological characteristics ofLigustrumjapohicum‘Howardii’. Journal of Shanxi Agricultural University (Natural Science Edition), 2017, 37(3): 183-188.
戴文, 邱國金, 史云光, 等. 鹽脅迫對金森女貞的生長與生理特性的影響. 山西農業大學(自然科學版), 2017, 37(3): 183-188.
[7] Han L P, Ma F J, Liu J T,etal. Analysis of oat-straw salt ion accumulation and the potential for improving saline-alkali soils in coastal Hebei province. Chinese Journal of Eco-Agriculture, 2012, 20(12): 1706-1712.
韓立樸, 馬鳳嬌, 劉金銅, 等. 遠東近濱海地區燕麥秸稈鹽分積累與改良鹽堿地潛力分析. 中國生態農業學報, 2012, 20(12): 1706-1712.
[8] Wu J Y, Liu J H, Zhai L J,etal. Salt tolerance of seed germination and seedling growth of different oat varieties. Chinese Journal of Ecology, 2009, 28(10): 1960-1965.
武俊英, 劉景輝, 翟利劍, 等. 不同品種燕麥種子萌發和幼苗生長的耐鹽性. 生態學雜志, 2009, 28(10): 1960-1965.
[9] Liu J X, Wang J C, Jia H Y. Different between physiological responses ofAvenanudaseedling to salt and alkali stresses. Journal of Soil and Water Conservation, 2015, 29(5): 331-336.
劉建新, 王金成, 賈海燕. 燕麥幼苗對鹽脅迫和堿脅迫的生理響應差異. 水土保持學報, 2015, 29(5): 331-336.
[10] Xia F, Chen L, Yan H,etal. Antioxidant and ultrastructural responses to priming with PEG in aged, ultra-dry oat seed. Seed Science and Technology, 2016, 44(3): 1-13.
[11] Bewley J D, Bradford K J, Hilhorst H W M,etal. Seeds: Physiology of development, germination and dormancy. 3rd. New York: Springer, 2013.
[12] Yu L, Liu Y H, Yuan W C,etal. Recent advances in the study of accumulation of ascorbic acid and its molecular mechanism in plants. Chinese Bulletin of Botany, 2016, 51(3): 396-410.
俞樂, 劉擁海, 袁偉超, 等. 植物抗壞血酸積累及其分子機制的研究進展. 植物學報, 2016, 51(3): 396-410.
[13] Lee Y P, Baek K H, Lee H S,etal. Tobacco seeds simultaneously over-expressing Cu/Zn-superoxide dismutase and ascorbate peroxidase display enhanced seed longevity and germination rates under stress conditions. Journal of Experiment Botany, 2010, 61(9): 2499-2506.
[14] Saeidi-Sar S, Abbaspour H, Afshari H,etal. Effects of ascorbic acid and gibberellin A3on alleviation of salt stress in common bean (PhaseolusvulgarisL.) seedlings. Acta Physiologiae Plantarum, 2013, 35(3): 667-677.
[15] Ejaz B, Sajid Z A, Aftab F. Effect of exogenous application of ascorbic acid on antioxidant enzyme activities, proline contents, and growth parameters ofSaccharumspp. hybrid cv. HSF-240 under salt stress. Turkish Journal of Biology, 2012, 36(6): 630-640.
[16] Beltagi M S. Exogenous ascorbic acid (vitamin C) induced anabolic changes for salt tolerance in chick pea (CicerarietinumL.) plants. African Journal of Plant Science, 2008, 2(10): 118-123.
[17] Chang Y X, Xu K D, Zhou L,etal. Ascorbic acid mitigating the inhibition of salt stress to wheat seedling growth. Journal of Triticeae Crops, 2013, 33(1): 151-155.
常云霞, 徐克東, 周琳, 等. 抗壞血酸對鹽脅迫下小麥幼苗生長抑制的緩解效應. 麥類作物學報, 2013, 33(1): 151-155.
[18] Khan A, Iqbal I, Shah A,etal. Alleviation of adverse effects of salt stress in brassica (Brassicacampestris) by pre-sowing seed treatment with ascorbic acid. American-Eurasian Journal of Agricultural and Environmental Science, 2010, 7(5): 557-560.
[19] ISTA. International rules for seed testing. Bassersdorf: Zurich, 2015.
[20] Abdul-Baki A A, Anderson J D. Vigour determination in soybean seed multiple criteria. Crop Science, 1973, 13(6): 630-633.
[21] Ellis R H, Roberts E H. The influence of genotype, temperature and moisture on seed longevity in chickpea, cowpea and soybean. Annals of Botany, 1982, 50(1): 69-82.
[22] Lu P N, Liu J H, Li Q,etal. Comparison of quality and yield of different oat varieties in saline-alkali land. Journal of Triticeae Crops, 2016, 36(11): 1510-1516.
盧培娜, 劉景輝, 李倩, 等. 鹽堿地不同燕麥品種的品質及產量比較. 麥類作物學報, 2016, 36(11): 1510-1516.
[23] Gao Z W, Lin J X, Shao S,etal. Effect of complex salt-alkali stresses on seed germination of oat. Pratacultural Science, 2014, 31(3): 451-456.
高戰武, 藺吉祥, 邵帥, 等. 復合鹽堿脅迫對燕麥種子發芽的影響. 草業科學, 2014, 31(3): 451-456.
[24] Zhang Q L, Liu Q, Gao H,etal. Effect of endogenous ascorbic acid on seed germination and seedling growth of rice. Journal of Tropical and Subtropical Botany, 2016, 24(3): 273-279.
張啟雷, 劉強, 高輝, 等. 內源抗壞血酸對水稻種子萌發及幼苗生長的影響. 熱帶亞熱帶植物學報, 2016, 24(3): 273-279.
[25] Shin R, Schachtman D P. Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(23): 8827-8832.
[26] Zhu Z J. Changes of activities of ascorbate-dependent H2O2-scavenging enzymes during imbibition and germination of oil rape seeds. Journal of Zhejiang Agricultural University, 1997, 23(5): 505-509.
朱祝軍. 油菜種子發芽過程中依賴于抗壞血酸的H2O2清除酶活性的變化. 浙江農業大學學報, 1997, 23(5): 505-509.
[27] Chang Y X, Xu K D, Liu B,etal. Alleviative effect of exogenous ascorbic acid on cadmium toxicity in wheat seedlings. Journal of Triticeae Crops, 2017, 37(2): 246-252.
常云霞, 徐克東, 劉彬, 等. 外源抗壞血酸對鎘毒害小麥幼苗的緩解作用. 麥類作物學報, 2017, 37(2): 246-252.
[28] Gallie D R. The role of L-ascorbic acid recycling in responding to environmental stress and in promoting plant growth. Journal of Experiment Botany, 2013, 64(2): 433-443.
[29] Liu Y J, Fan J, Han X K,etal. Effect of exogenous vitamins on seed germination and recovery ofMedicagosativaL. under high NaCl stress. Chinese Agricultaural Science Bulletin, 2015, 31(26): 12-27.
劉艷軍, 范晶, 韓學珅, 等. 外源維生素對高NaCl脅迫下紫花苜蓿種子萌發及恢復性的影響. 中國農學通報, 2015, 31(26): 12-27.
[30] Jiang X W, Li H Q, Wang J H. Physiological response ofScutellariabaicalensisseed germination and seedling to exogenous ascorbic acid under salt stress. Plant Physiology Journal, 2015, 51(2): 166-170.
江緒文, 李賀勤, 王建華. 鹽脅迫下黃芩種子萌發及幼苗對外源抗壞血酸的生理響應. 植物生理學報, 2015, 51(2): 166-170.
[31] Fan M H, Zhang Y X, Shi Y,etal. Effects of exogenous ascorbic acid on seed germination and growth ofBrassicanapusunder seawater stress. Chinese Journal of Oil Crop Science, 2009, 31(1): 34-38.
范美華, 張義鑫, 石戈, 等. 外源抗壞血酸對油菜種子在海水脅迫下萌發生長的影響. 中國油料作物學報, 2009, 31(1): 34-38.
[32] Shi Y C, Yang Y Y, Xue R L,etal. Research advance of biological function of ascorbic acid in plants. Plant Physiology Journal, 2015, 51(1): 1-8.
石永春, 楊永銀, 薛瑞麗, 等. 植物中抗壞血酸的生物學功能研究進展. 植物生理學報, 2015, 51(1): 1-8.
[33] Wang C, Zhu Y L, Yang L F,etal. Effects of NaCl stress on ascorbate-glutathione cycle in vegetable soybean seeds. Plant Nutrition and Fertilizer Science, 2010, 16(5): 1209-1216.
王聰, 朱月林, 楊立飛, 等. NaCl脅迫對菜用大豆種子抗壞血酸-谷胱甘肽循環的影響. 植物營養與肥料學報, 2010, 16(5): 1209-1216.
[34] Liu Z P, Li B B, Xue H N,etal. Effect of NaCl stress on antioxidant system and ascorbate-glutathione cycle in barley seeds. Journal of Triticeae Crops, 2016, 36(6): 736-741.
劉志萍, 李琲琲, 薛海楠, 等. NaCl脅迫對大麥籽粒抗壞血酸-谷胱甘肽循環的影響. 麥類作物學報, 2016, 36(6): 736-741.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
