川西北高寒沙地不同年限高山柳林下優(yōu)勢植物碳、氮、磷生態(tài)化學計量特征
2018-04-18 07:51:08佘淑鳳胡玉福舒向陽嚴星李智王琴何佳賈安都
草業(yè)學報 2018年4期
關鍵詞:植物
佘淑鳳,胡玉福,舒向陽,嚴星,李智,王琴,何佳,賈安都
(四川農(nóng)業(yè)大學資源學院,四川 成都611130)
生態(tài)化學計量學是一門研究生態(tài)系統(tǒng)和生態(tài)過程中多重化學元素平衡關系的科學,它主要強調活有機體組成元素(特別是C、N和P)的化學計量特征關系[1]。碳(C)是植物體內干物質最主要的構成元素,也是各種生理生化過程的能量來源[2]。氮(N)、磷(P)分別作為蛋白質和核酸的重要組成元素,同時也是植物生長所需基本營養(yǎng)元素。C∶N和C∶P在一定程度上可以反映植物生長速度,并且能夠表明植物生長速度與植物N和P的利用效率的相關性[3]。植物葉片N∶P的臨界值被認為是判斷土壤對植物養(yǎng)分供應狀況的指標,因此植物N、P化學計量研究成為揭示植物養(yǎng)分限制狀況及其適應策略的手段[4]。研究碳氮磷生態(tài)化學計量特征有助于認識植物應對不同生境條件的養(yǎng)分利用策略,對于解釋植物與生境間的耦合關系具有重要意義[5]。
林下植被是林地養(yǎng)分循環(huán)系統(tǒng)的重要組成,并在維護生態(tài)系統(tǒng)多樣性和穩(wěn)定性方面有突出作用[6]。其對灌木的生長、發(fā)育及地下過程如分解、土壤養(yǎng)分流動和積累等均產(chǎn)生很大影響[7]。同時大量研究表明,灌木在林下植被的生存、生長、繁殖等過程中也起到了重要的促進作用,尤其是在極端環(huán)境條件下[8-11]。林下植被的生長主要受到環(huán)境、灌木類型及發(fā)育狀況等因素的影響[7,12]。灌草的相互作用如競爭、干擾等被認為是構成植物種群結構的主要方式[13]。但隨著灌木年限的增長,林地養(yǎng)分循環(huán)系統(tǒng)的結構及組成也會隨之變化,同時灌木與林下植被的競爭關系日益凸顯,進而影響林下植被的生長及養(yǎng)分分配格局。研究林下植被主要元素的分配及化學計量特征隨年限產(chǎn)生的變化有助于闡明植物體的內穩(wěn)性特征,揭示該區(qū)植物生長狀況和生境養(yǎng)分供應狀況。目前,國內外對于植物生態(tài)化學計量學已有大量研究,但多集中于濕地[14]或人造林生態(tài)系統(tǒng)[15-16],在高寒沙地生態(tài)系統(tǒng)的研究主要集中于土壤特性[17-18]方面,關于林下植被生態(tài)化學計量特征方面的研究鮮少。
本研究選取川西北高寒沙地不同年限高山柳灌木林下藏沙蒿(Artemisiawellbyi)、裂葉獨活(Heracleummillefolium)和鐮莢棘豆(Oxytropisfalcata)3種優(yōu)勢草本植物為對象,以植物生態(tài)化學計量學為研究手段,探究不同年限高山柳灌木林下藏沙蒿、裂葉獨活和鐮莢棘豆葉片和根碳、氮、磷化學計量特征及其變化特征,同時研究3種植物在沙地生態(tài)系統(tǒng)中的主要養(yǎng)分限制狀況,為提高區(qū)域植被覆蓋率和改善沙地修復的養(yǎng)分供應措施提供理論依據(jù)。
1 材料與方法
1.1 試驗地概況
研究區(qū)位于四川省阿壩藏族羌族自治州紅原縣,地處青藏高原東部邊緣、四川省西北部,地理坐標為N 31°51′-33°19′、E 101°51′-103°23′,境域分屬長江、黃河兩大水系。地勢為東南向西北傾斜,地貌具有山原向丘狀高原過渡的典型特征,平均海拔在3400 m以上;屬高原大陸性寒溫帶季風氣候,干濕季節(jié)分明,雨熱同期,降水主要集中于5-10月,年降水量達 686.75 mm;空氣較為稀薄,年均氣壓為66.56 KPa;氣候寒冷,年均溫1.1 ℃,極端低溫為-33.5 ℃;日照時間較長,年均日照時長約為2158.7 h,太陽輻射強,年總量為6194 MJ·m-2;冰凍期長,寒凍風化作用強烈,季節(jié)性凍土分布普遍。植被以藏沙蒿、沙生苔草(Carexpraeclara)、鐮莢棘豆、裂葉獨活、線葉嵩草(Kobresiacapillifolia)、賴草(Leymussecalinus)、淡黃香青(Anaphalisflavescens)、黑穗苔草(Carexatrata),木里苔草(Carexmuliensis)、細葉亞菊(Ajaniatenuifolia)等為主,植被組合以亞高山草甸為主,沼澤草甸與沼澤植被較為發(fā)達,植物群落外貌鮮艷,富有季相變化。
1.2 試驗材料與方法
樣品采集于2015年8月,在研究區(qū)內選擇人為活動影響較小的地區(qū)作為樣品采集地點。通過對紅原縣高寒草地的實地勘察,在川西北沙化治理典型示范區(qū)內共設置了3個年限(6、18、24年)的高山柳實驗樣地來代表不同年限的高山柳,選取作為林下植被代表,因高山柳林下草本菊科植物藏沙蒿、豆科植物鐮莢棘豆、傘形科植物裂葉獨活這3種植物在6、18和24年高山柳林下均為優(yōu)勢植物,能夠起到代表和對比作用,將其作為研究對象。各年限分別設置3個10 m×10 m的樣方, 在植物樣方調查基礎下采集完整植物樣品, 樣方基本信息見表1。植物樣品取回后,洗掉植株樣品根部泥土后快速風干;然后分葉、根裝袋,在105 ℃下10 min殺青,最后80 ℃烘干至恒質量不變,稱干質量。采用重鉻酸鉀容量法測定植物樣品的有機碳含量,采用半微量凱氏法測定全氮含量,采用鉬銻抗比色法測定全磷含量[19]。
1.3 數(shù)據(jù)處理與分析
采用Microsoft Excel 2003對數(shù)據(jù)進行處理、整理和繪圖,用SPSS 19.0對數(shù)據(jù)進行差異顯著性檢驗(LSD法)和相關性分析。
2 結果與分析
2.1 不同年限高山柳林下植被C含量變化
葉片C含量表現(xiàn)為藏沙蒿>裂葉獨活>鐮莢棘豆,三者的C含量范圍分別為252.98~322.60 mg·g-1、 66.73~71.34 mg·g-1和203.20~299.29 mg·g-1

表1 樣方基礎信息Table 1 Basic information of samples
(圖1A)。林下植被葉片C含量隨年限增加呈下降趨勢,但不同植物間表現(xiàn)出不同變化趨勢。藏沙蒿葉片C含量在6年達到最大,后降低并保持穩(wěn)定,在18和24年間無顯著差異;鐮莢棘豆葉片C含量在6、18、24年間均無顯著差異;裂葉獨活葉片C含量隨年限增加先增加,在18年時達到最大值后降低,且變化顯著(圖 1A)。高山柳林下植被根中C含量隨年限增加無顯著變化,根部C含量表現(xiàn)為裂葉獨活>藏沙蒿>鐮莢棘豆,其中裂葉獨活為387.95~400.95 mg·g-1,藏沙蒿為150.39~180.68 mg·g-1,鐮莢棘豆為143.87~164.21 mg·g-1(圖 1B)。

圖1 不同年限高山柳林下植被C含量變化 Fig.1 Changes in the understory plants C content in different plantations age of S. cupularis 不同字母表示同一植物不同林齡差異顯著(P<0.05),下同。Different lowercase letters in the same plantation indicate significant differences among the plantation age (P<0.05). The same below.
2.2 不同年限高山柳林下植被N含量變化
葉片N含量表現(xiàn)為裂葉獨活>藏沙蒿>鐮莢棘豆,三者的 N含量范圍分別為37.40~52.77 mg·g-1、22.77~47.96 mg·g-1和 13.29~34.70 mg·g-1(圖2A)。年限對林下植被葉片N含量影響較顯著,且3種植物呈不同的變化趨勢。隨年限增加,藏沙蒿葉片N含量先下降,在18年達到最低點后略上升;鐮莢棘豆隨高山柳年限的增長而顯著增加(P<0.05),在24年達到最大值;裂葉獨活在6、18、24年均為三者最高,且隨年限增長先下降,在18年達到最低點后再上升(圖 2A)。高山柳林下植被中,鐮莢棘豆根部N含量顯著高于藏沙蒿和鐮莢棘豆,三者分別為20.69~70.04 mg·g-1、20.33~37.47 mg·g-1和1.19~43.46 mg·g-1(圖 2B)。
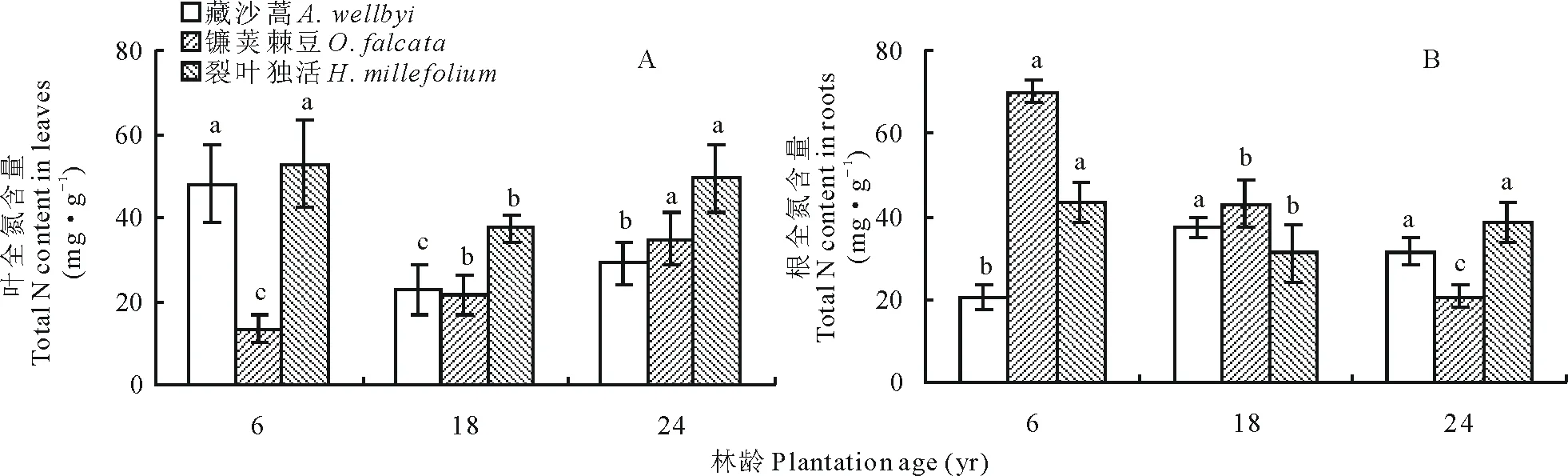
圖2 不同年限高山柳林下植被N含量變化Fig.2 Changes in the understory plants N content in different plantations age of S. cupularis
2.3 不同年限高山柳林下植被P含量變化
葉片P含量隨年限增長變化顯著,裂葉獨活、藏沙蒿和鐮莢棘豆分別為1.90~2.52 mg·g-1、1.08~3.05 mg·g-1和2.01~2.29 mg·g-1(圖 3A)。隨著高山柳種植年限增長,其林下植被的葉和根P含量變化均呈相反趨勢。藏沙蒿葉全P含量隨年限增長先降低再升高,根中P含量隨年限增長先升高再降低;鐮莢棘豆葉全P含量隨年限增長呈增長趨勢,其根則逐漸降低;裂葉獨活葉全P含量隨年限增長呈逐漸降低趨勢,其根隨年限增長而逐漸升高。其中藏沙蒿葉全P含量隨年限增長變化較大,而鐮莢棘豆和裂葉獨活無顯著變化(P>0.05)(圖 3B)。
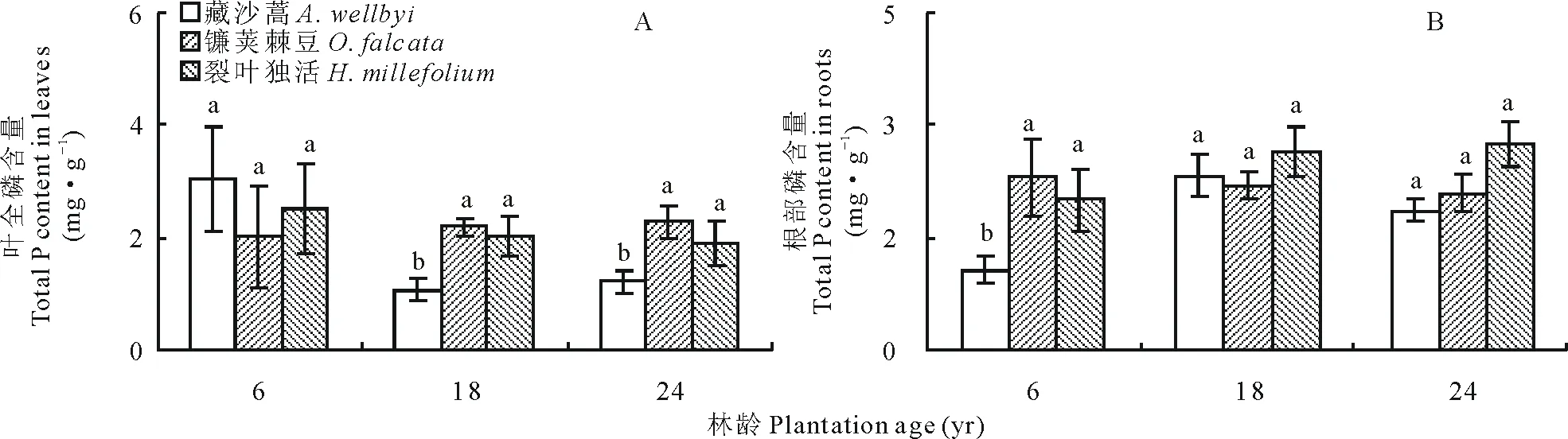
圖3 不同年限高山柳林下植被P含量變化Fig.3 Changes in the understory plants P content in different plantations age of S. cupularis
2.4 不同年限高山柳林下植被C∶N
葉片C∶N表現(xiàn)為藏沙蒿>裂葉獨活>鐮莢棘豆,三者分別為6.73~11.17、4.12~8.00和1.92~5.22(圖 4A)。年限對3種草本植物葉片C∶N均有顯著影響(P<0.05),藏沙蒿在6年最小,先隨年限增長而增加,在18年達到最大值后下降;裂葉獨活隨年限增長先增加,在18年達到最大值后下降至最小值;鐮莢棘豆則隨年限增長一直降低,24年時達到最低(圖 4)。高山柳林下植被根部C∶N隨年限增長變化顯著,藏沙蒿隨年限增長先由最大值8.89下降至最低值4.01后增加;鐮莢棘豆隨年限增長而增加,在24年達到最大值7.94;裂葉獨活隨年限增長先增加至最大值12.86后降低(圖 4B)。
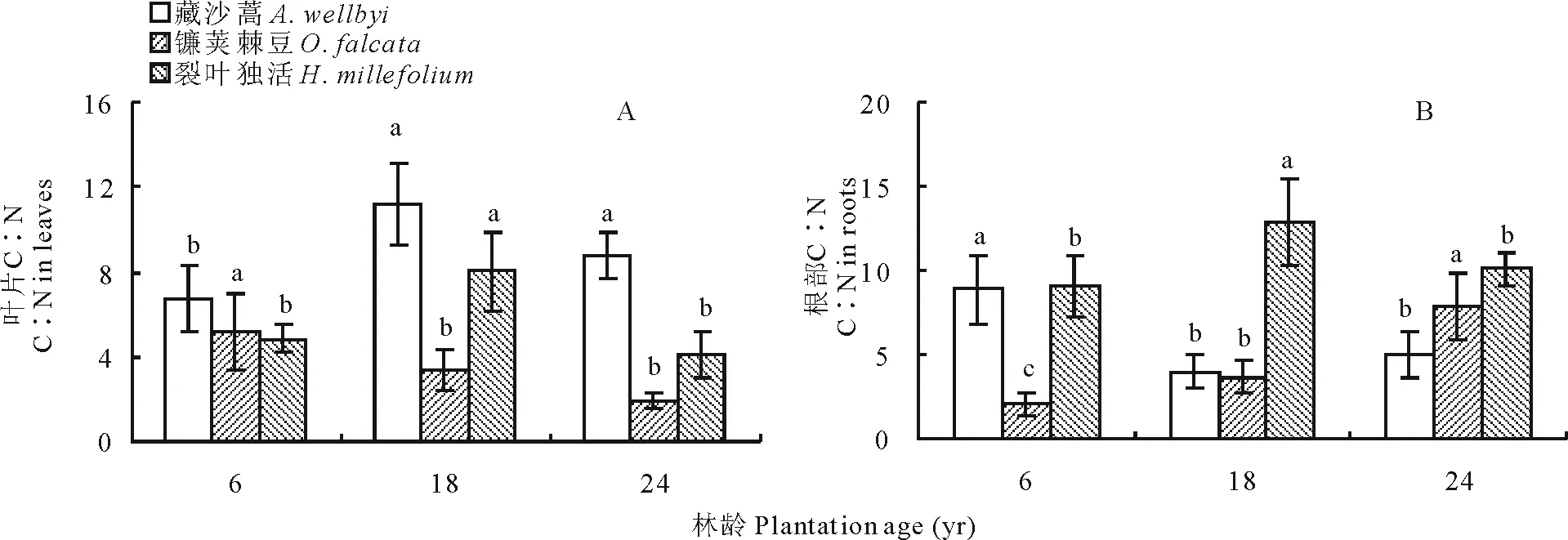
圖4 不同年限高山柳林下植被含量C∶N變化Fig.4 Changes in the understory plants C∶N in different plantations age of S. cupularis
2.5 不同年限高山柳林下植被C∶P
高山柳林下植被葉片C∶P表現(xiàn)為藏沙蒿>裂葉獨活>鐮莢棘豆,藏沙蒿和裂葉獨活隨年限增長變化顯著(P<0.05),藏沙蒿隨年限增長由6年時最低值105.65增加至18年時最大值235.66而后再降低;裂葉獨活同藏沙蒿一樣由最低值101.67先增加至最高值147.58而后降低;鐮莢棘豆隨年限增長差異不顯著,有下降趨勢(圖5A)。根部C∶P隨年限增長變化顯著,藏沙蒿隨年限增長先下降,在18年降至最小值64.50后增加;裂葉獨活隨年限增長一直下降,在6年為最大值196.80,24年時為最小值141.68;而鐮莢棘豆隨年限增長一直增加,在24年達到最大值78.32(圖 5B)。
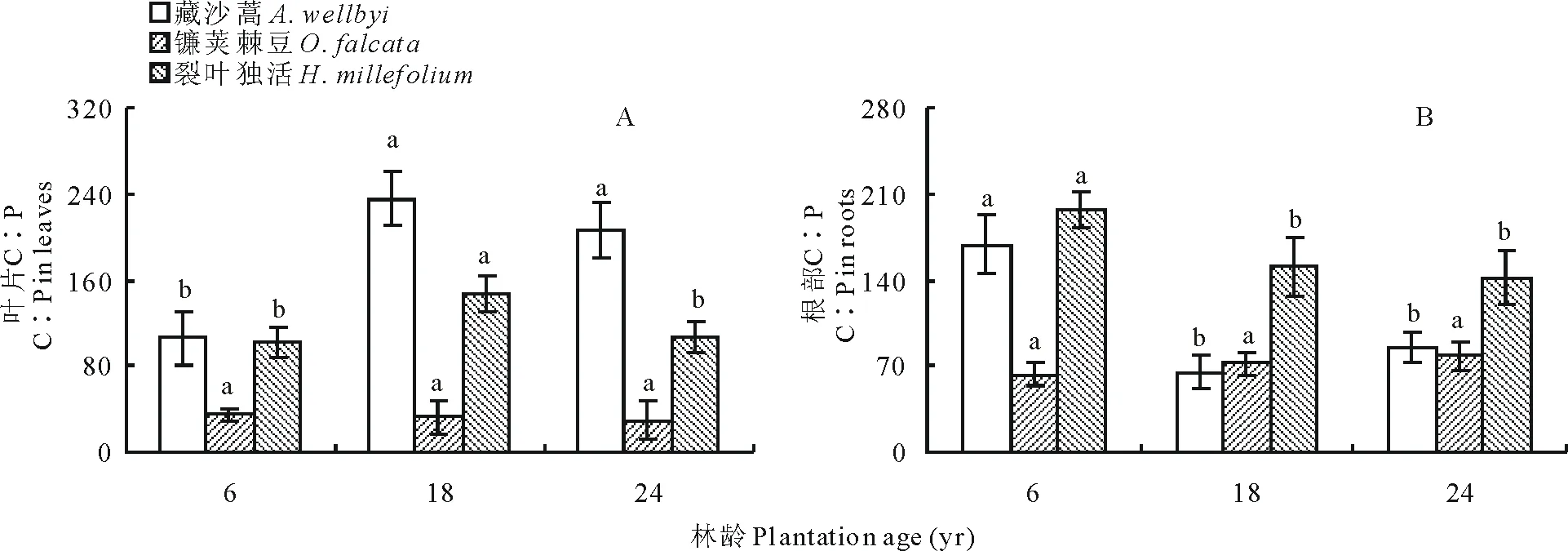
圖5 不同年限高山柳林下植被含量C∶P變化Fig.5 Changes in the understory plants C∶P in different plantations age of S. cupularis
2.6 不同年限高山柳林下植被N∶P
葉片N∶P表現(xiàn)為裂葉獨活>藏沙蒿>鐮莢棘豆,三者分別為18.44~25.89、15.70~23.69和6.60~15.17(圖 6A)。年限對3種草本植物均有顯著影響(P<0.05),藏沙蒿和鐮莢棘豆隨年限增長均顯著增加,由6年最低值增加至24年最高值;裂葉獨活則隨年限增長先下降而后增加至最高值(圖 6A)。根部N∶P隨年限增長變化顯著(P<0.05),鐮莢棘豆隨年限增長顯著下降,由6年最高值30.44降至24年最低值9.87;藏沙蒿隨年限增長則呈先下降后上升的趨勢;裂葉獨活隨年限增長也由最高值21.58先降至最低值11.76而后增加(圖 6B)。
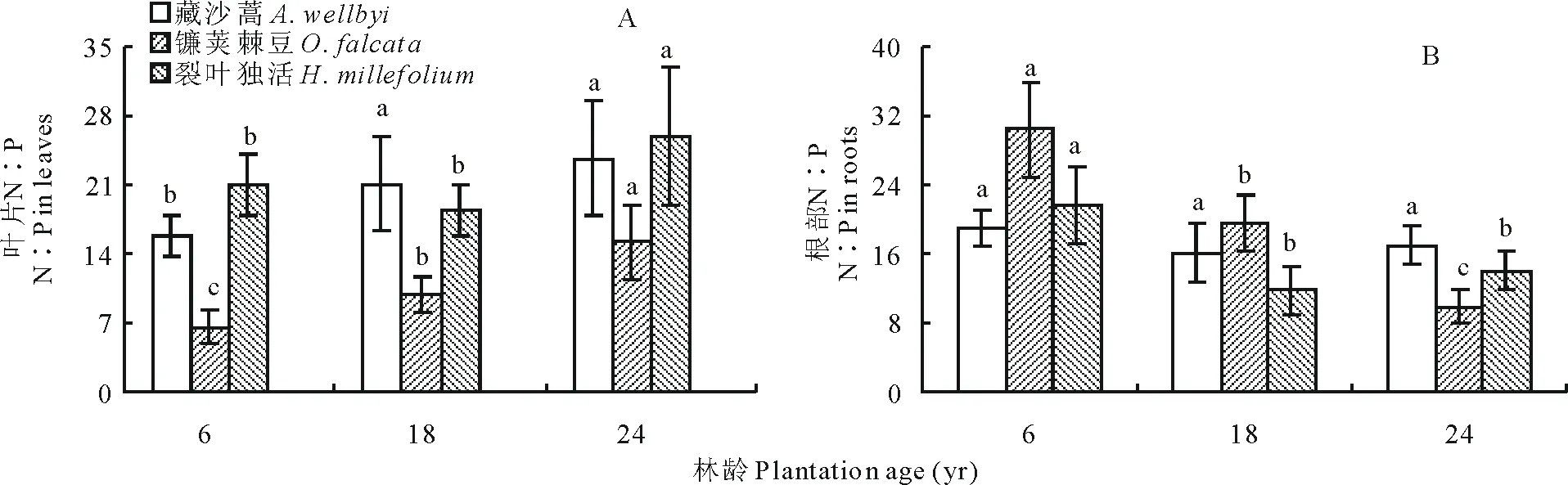
圖6 不同年限高山柳林下植被含量N∶P變化Fig.6 Changes in the understory plants N∶P in different plantations age of S. cupularis
3 討論
植物葉片C∶N、C∶P可以表征植物吸收營養(yǎng)元素同化C的能力,在一定程度上可以反映出單位養(yǎng)分供應量所能達到的生產(chǎn)力、植物對營養(yǎng)的利用效率以及植物的生長速度[20]。Sterner等[21]提出的生長率假說認為,生物體的快速生長需要大量的核糖體RNA合成蛋白質,由于核糖體RNA中含有大量P,從而使得生長率高的生物具有較低的C∶N和C∶P。本研究結果表明,在18和24年高山柳林下植被C∶P明顯高于6年,其原因可能是造林過程中灌木和多年生草本植物對N、P的攝取導致土壤中全N和全P含量減少,進而限制了植物吸收攝取。這一研究結果與崔寧潔等[22]對不同林齡馬尾松生態(tài)化學計量特征研究結果相一致,其研究結果表明,該地區(qū)馬尾松生長早期土壤相對貧瘠,可供植物利用的N、P養(yǎng)分較少,從而限制了馬尾松的生長發(fā)育。較高的C∶N和C∶P也表明植物對于N、P元素利用效率較高,這也證實了植物往往以較高的元素利用效率應對貧瘠的養(yǎng)分狀態(tài)[23]。
Aerts等[24]研究表明,植物對 N、P 養(yǎng)分缺乏的適應可體現(xiàn)在植物N∶P化學計量比的變化上,因此,N∶P化學計量比可作為當前限制性養(yǎng)分判斷的指標之一。Drenovsky等[25]研究表明,植物N∶P<14反映植物生長受N限制,N∶P >16反映植物受P限制,14
4 結論
在川西北高山沙地修復過程中,隨著灌木高山柳年限的增加,其林下植被藏沙蒿、鐮莢棘豆和裂葉獨活葉片和根C、N、P含量及其比值發(fā)生了顯著變化。不同高山柳林下植被N含量差異顯著,隨著年限增長葉片N含量增加而根部N含量逐漸減少;林下植被P含量呈不同的變化趨勢。隨著高山柳年限增長,灌木和草本植物對土壤養(yǎng)分的不斷攝取,林下植被葉片C∶N和C∶P較高,植被生長受到限制,但同時也反映出在養(yǎng)分供給缺乏的條件下植物對養(yǎng)分的利用效率較高;林下植被N∶P隨年限增加而增加,其中不同年限林下藏沙蒿和裂葉獨活葉片N∶P均大于16,鐮莢棘豆由6.60增長到15.17,由受N限制轉為受N和P的共同限制,而林下植被根部N∶P隨年限增加而降低,但仍表現(xiàn)出主要受P限制,因而該地區(qū)林下植被的生長主要受到P的限制,為提高區(qū)域植被覆蓋率,改善沙漠化情況應合理使用P肥以改善養(yǎng)分供應狀況,且對于年限較大的高山柳林下植物補充P尤為重要。
參考文獻References:
[1] Chrzanowski T H, Kyle M, Elser J J,etal. Element ratios and growth dynamics of bacteria in an oligotrophic Canadian shield lake. Aquatic Microbial Ecology, 1996, 11(2): 119-125.
[2] Qi D H, Wen Z M, Wang H X,etal. Stoichiometry traits of carbon, nitrogen, and phosphorus in plants of different functional groups and their responses to micro-topographical variations in the hilly and gully region of the Loess Plateau, China. Acta Ecologica Sinica, 2016, 36(20): 6420-6430.
戚德輝, 溫仲明, 王紅霞, 等. 黃土丘陵區(qū)不同功能群植物碳氮磷生態(tài)化學計量特征及其對微地形的響應. 生態(tài)學報, 2016, 36(20): 6420-6430.
[3] Niklas K J, Cobb E D. N, P, and C stoichoimetry ofEranthishyemalis(Ranunculaceae) and the allometry of plant growth. American Journal of Botany, 2005, 92(8): 1256-1263.
[4] Li C J, Lei J Q, Xu X W,etal. The stoichiometric characteristics of C, N, P for artificial plants and soil in the hinterland of Taklimakan Desert. Acta Ecologica Sinica, 2013, 33(18): 5760-5767.
李從娟, 雷加強, 徐新文, 等. 塔克拉瑪干沙漠腹地人工植被及土壤CNP的化學計量特征. 生態(tài)學報, 2013, 33(18): 5760-5767.
[5] Hu W F, Zhang W L, Zhang L H,etal. Stoichiometric characteristics of nitrogen and phosphorus in major wetland vegetation of China. Chinese Journal of Plant Ecology, 2014, 38(10): 1041-1052.
胡偉芳, 章文龍, 張林海, 等. 中國主要濕地植被氮和磷生態(tài)化學計量學特征. 植物生態(tài)學報, 2014, 38(10): 1041-1052.
[6] Vander Schaaf C L. Estimating understory vegetation response to multi-nutrient fertilization in Douglas-fir and ponderosa pine stands. Journal of Forest Research, 2008, 13(1): 43-51.
[7] Nilsson M C, Wardle D A. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Frontiers in Ecology and the Environment, 2005, 3(8): 421-428.
[8] He Q, Cui B, Bertness M D,etal. Testing the importance of plant strategies on facilitation using congeners in a coastal community. Ecology, 2012, 93(9): 2023-2029.
[9] Cavieres L A, Badano E I, Sierra-Almeida A,etal. Positive interactions between alpine plant species and the nurse cushion plantLaretiaacaulisdo not increase with elevation in the Andes of central Chile. New Phytologist, 2006, 169(1): 59-69.
[10] Armas C, Pugnaire F I. Ontogenetic shifts in interactions of two dominant shrub species in a semi-arid coastal sand dune system. Journal of Vegetation Science, 2009, 20(3): 535-546.
[11] Zhou G X, Wang G J, Li L,etal. C, N, P stoichiometry characteristic of Chinese fir root, branch and leaf. Hunan Forestry Science and Technology, 2015, 42(1): 15-18.
周國新, 王光軍, 李櫟, 等. 杉木根, 枝和葉的 C, N, P 生態(tài)化學計量特征. 湖南林業(yè)科技, 2015, 42(1): 15-18.
[12] Barbier S, Gosselin F, Balandier P. Influence of tree species on understory vegetation diversity and mechanisms involved-a critical review for temperate and boreal forests. Forest Ecology and Management, 2008, 254(1): 1-15.
[13] Zhang G, Zhao L, Yang Q,etal. Effect of desert shrubs on fine-scale spatial patterns of understory vegetation in a dry-land. Plant Ecology, 2016, 217(9): 1141-1155.
[14] Wang W Q, Xu L L, Zeng C S,etal. Carbon, nitrogen and phosphorus ecological stoichiometric ratios among live plant-litter-soil systems in estuarine wetland. Acta Ecologica Sinica, 2011, 31(23): 7119-7124.
王維奇, 徐玲琳, 曾從盛, 等. 河口濕地植物活體-枯落物-土壤的碳氮磷生態(tài)化學計量特征. 生態(tài)學報, 2011, 31(23): 7119-7124.
[15] Wang S Q, Yu G R. Ecological stoichiometry characteristics of ecosystem carbon, nitrogen and phosphorus elements. Acta Ecologica Sinica, 2008, 28(8): 3937-3947.
王紹強, 于貴瑞. 生態(tài)系統(tǒng)碳氮磷元素的生態(tài)化學計量學特征. 生態(tài)學報, 2008, 28(8): 3937-3947.
[16] Fan H, Wu J, Liu W,etal. Linkages of plant and soil C∶N∶P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil, 2015, 392(1/2): 127-138.
[17] Hu Y F, Peng J J, Deng L J,etal. Influences of fencing and planting branchy tamarisk on soil particles composition and mineral nutrients in desertization land in northwestern Sichuan Province. Chinese Journal of Soil Science, 2015, (1): 54-61.
胡玉福, 彭佳佳, 鄧良基, 等. 圍欄種植紅柳對川西北高寒沙地土壤顆粒組成和礦質養(yǎng)分的影響. 土壤通報, 2015, (1): 54-61.
[18] Feng H L, Hu Y F, Shu X Y,etal. Influences of adding needle mushroom edible fungi residues on vegetation and soil enzyme activity of sandy soil in northwest Sichuan. Journal of Soil, 2016, (5): 161-165.
馮慧翎, 胡玉福, 舒向陽, 等. 金針菇菌渣對川西北高寒沙地植被及土壤酶活性的影響. 水土保持學報, 2016, (5): 161-165.
[19] He J S, Han X G. Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems. Chinese Journal of Plant Ecology, 2010, 34(1): 2-6.
賀金生, 韓興國. 生態(tài)化學計量學: 探索從個體到生態(tài)系統(tǒng)的統(tǒng)一化理論. 植物生態(tài)學報, 2010, 34(1): 2-6.
[20] Liu W D, Su J R, Li S F,etal. Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Acta Ecologica Sinica, 2010, (23): 6581-6590.
劉萬德, 蘇建榮, 李帥鋒, 等. 云南普洱季風常綠闊葉林演替系列植物和土壤 C, N, P 化學計量特征. 生態(tài)學報, 2010, (23): 6581-6590.
[21] Sterner R W, Elser J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton: Princeton University Press, 2002.
[22] Cui N J, Liu X B, Zhang D J,etal. The distribution pattern of carbon, nitrogen and phosphorus and the stoichiometry characteristics ofPinusmassonianaplantation in different age. Ecology and Environmental Science, 2014, 23(2): 188-195.
崔寧潔, 劉小兵, 張丹桔, 等. 不同林齡馬尾松(Pinusmassoniana)人工林碳氮磷分配格局及化學計量特征. 生態(tài)環(huán)境學報, 2014, 23(2): 188-195.
[23] Bowman W D. Accumulation and use of nitrogen and phosphorus following fertilization in two alpine tundra communities. Oikos, 1994, 70(2): 261-270.
[24] Aerts R, Chapin F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30(8): 1-67.
[25] Drenovsky R E, Richards J H. Critical N∶P values: predicting deficiencies in desert shrublands. Plant and Soil, 2004, 259(1): 59-69.
[26] Dong W Y, Nie L S, Li J Y,etal. Effects of nitrogen forms on the absorption and distribution of nitrogen inPopulustomentosaseedlings using the technique of15N tracing. Journal of Beijing Forestry University, 2009, (4): 97-101.
董雯怡, 聶立水, 李吉躍, 等. 應用15N示蹤研究毛白楊苗木對不同形態(tài)氮素的吸收及分配. 北京林業(yè)大學學報, 2009, (4): 97-101.
[27] Zhang C H, Wang Z M, Ju W M,etal. Spatial and temporal variability of soil C/N ratio in Songnen Plain Maize Belt. Environmental Science, 2011, 32(5): 1407-1414.
張春華, 王宗明, 居為民, 等. 松嫩平原玉米帶土壤碳氮比的時空變異特征. 環(huán)境科學, 2011, 32(5): 1407-1414.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
