岷縣不同茬口對當歸苗栽生長及抗病性的影響
2018-04-18 07:48:35金彥博郭鳳霞陳垣白剛袁洪超梁偉
草業學報 2018年4期
金彥博,郭鳳霞*,陳垣,2*,白剛,袁洪超,梁偉
(1.甘肅省中藥材規范化生產技術創新重點實驗室,甘肅省藥用植物栽培育種工程研究中心,甘肅省干旱生境作物學重點實驗室,甘肅農業大學生命科學技術學院,甘肅農業大學農學院,甘肅 蘭州 730070;2.甘肅天士力中天藥業有限責任公司,甘肅省特色藥用植物資源保護與利用工程實驗室,甘肅省特色藥材規范化可追溯栽培工程技術研究中心,甘肅 定西 748100)
當歸(Angelicasinensis)為傘形科多年生草本藥用植物,以根入藥,具補血活血和調經止痛的功效,是我國常用大宗藥材之一[1]。我國西北地區的甘肅省岷縣、漳縣、渭源和宕昌一帶是我國當歸的主要道地產區[2]。其中岷縣一帶由于特殊的地形地貌和氣候條件為當歸的生長繁育提供了得天獨厚的自然條件,是我國最大的當歸產區,已有1500多年栽培史,占全國年產量的90%以上,栽培的當歸品質最佳,享譽海內外,于2001年6月被中國特產之鄉推薦暨宣傳活動組織委員會命名為“中國當歸之鄉”[3]。
當歸育苗環境條件嚴苛,要求氣候冷涼陰濕[2]。由于當歸忌連作,長期以來,當歸采用生荒地育苗,即在岷縣和漳縣等高寒陰濕區通過砍伐原生態灌木林,開挖草皮,翻耕后進行育苗。隨著中藥材現代化的發展,當歸藥材需求量逐年增加,種苗供不應求,每年開荒面積已達460 hm2之多[4]。溫室效應的加劇導致適宜當歸育苗區域愈加狹窄,可利用荒地資源已延伸至海拔2700 m以上陡險地帶,荒地植被破壞引起水土流失加劇[5],探尋熟地育苗條件成為促進當歸可持續化發展的唯一途徑。茬口是指栽培上茬作物時給予土地影響的總稱。茬口形成過程中伴隨一系列生物因子和非生物因子的變化,進而對后茬作物產量和品質產生直接或間接影響[6]。研究證明,用豆科牧草和禾本科作物輪作既能提高牧草和作物產量,又可改善土壤條件[7]。彭云等[8]研究表明不同前茬明顯影響煙株的生長、產量、產值和評吸質量。適宜的輪作體系是農田高效利用的基礎。盡管目前對當歸連作障礙的報道研究也較深入,但因生態區和土壤質地的不同存在較大差異,且僅局限于對種苗移栽后成藥期的生產調查研究[9-12],對當歸育苗階段的研究也僅停留在盆栽發芽試驗階段[13],由于實驗室試驗不能完全模擬滿足農田綜合環境條件。因此,在當歸道地產區有計劃種植當地適宜作物,培育作物茬口基礎上再行當歸育苗,探索茬口造成的土壤異質性對影響當歸種苗質量和產量的內在因素具有重要意義,可為探尋可替代生荒地育苗的農茬口資源提供科學依據,有效促進該區土壤資源的可持續化利用和農田生態環境的保護。
1 材料與方法
1.1 試驗區概況
研究區設于甘肅省定西市岷縣禾馱鄉,該區位于N 34°25′27″,E 104°28′24″,海拔2783 m,年平均氣溫為5.5 ℃,最熱7月平均氣溫16 ℃,最冷1月平均氣溫-6.9 ℃,年降水量為635 mm,年無霜期123 d,平均相對濕度68%,平均日照時數2214.9 h,高寒陰濕,植被覆蓋良好,屬高原型大陸氣候。適宜栽培喜涼作物如馬鈴薯(Solanumtuberosum)、油菜(Brassicacampestris)、青稞(Hordeumvulgarevar.nudum)、蠶豆(Viciafaba)等和一些藥用植物如當歸、蒙古黃芪(Astragalusmembranaceusvar.mongholicus)等。
1.2 茬口培育試驗
茬口培育試驗于2015年4月在岷縣禾馱鄉石家臺村甘肅農業大學當歸研究基地進行,采用單因素隨機區組設計,3次重復,區組間距0.5 m,小區長2.5 m,寬1.2 m,小區面積2.5 m×1.2 m=3 m2,即在2014年種植當歸基礎上,2015年按試驗地環境條件(坡度)將試驗地劃分為3個區組,同一區組處于同一坡度,其余環境條件也相對一致,在每個區組內劃分4個小區,分別隨機種植小麥(Triticumaestivum)、蒙古黃芪、馬鈴薯和當歸4種作物,以培育小麥茬、黃芪茬、馬鈴薯茬和當歸重茬。小麥采用撒播,播種量225 kg·hm-2。馬鈴薯穴播,每畦2行,行穴距40 cm,株距50 cm。蒙古黃芪和當歸均采用開溝播種,株距10 cm,行距20 cm,作物成熟后將各小區作物及時收獲,維持小區狀態自然越冬,備用于次年不同茬口當歸育苗試驗。
1.3 當歸播種育苗試驗
不同茬口當歸育苗試驗于2016年6月5日在上述對應茬口區組小區進行,試驗重復3次,當歸種子來源于甘肅省漳縣菜子川村2015年繁種田采收的種子。播種前每小區施有機肥(甘肅天士力中天羊業有機肥廠生產,主要原料為羊糞)1.5 kg,采用撒播播種,每小區播種量10 g(種子大小和飽滿度基本一致,千粒重為1.6326 g,凈度為97.7%),各小區控制均勻一致,播后用鐵篩在當歸種子表面撒蓋一薄層土壤,覆土厚度為0.4 mm左右,以正好蓋住當歸種子為宜,后覆蓋一層小麥秸稈以保溫保濕,秸稈厚度為2~3 cm。當歸育苗期田間管理控制在一致的水平,整個育苗期人工除草3次。
2016年8月中旬當歸出苗整齊后揭掉麥草,用白色線繩在每小區中央劃定0.5 m×0.5 m小樣方(sample square),統計各重復小區樣方苗數,并在每個樣方中央選定標記5株作為抽樣單位測定株高、株幅、莖粗和真葉數,此后每月中旬統計測定1次,直到育苗結束,當歸育苗期共測定3次(8月10日,9月21日和10月22日)。株高以測定距地面植株垂直高度為標準,株幅以株冠最大距離為標準,均采用卷尺(精度為0.1 cm)測定。莖粗采用數顯游標卡尺測定(精度0.001 cm)。
1.4 當歸苗栽質量性狀指標和抗病性指標測定
2016年10月22日當歸育苗結束,用叉鋤松動當歸種苗根圍土壤,收集整個小區當歸種苗。按照陳垣等[14]的方法在各小區隨機抽取20株統計當歸種苗根腐病等級,計算發病率和病情指數。
發病率(disease ratio)=病根數×100%/調查總根數
病情指數(disease index)=∑(各級根數×病級數)×100%/(調查總數×最高級)
根病統計結束,各茬口重復小區隨機抽取10株無病完整苗,帶土低溫運回實驗室,清理干凈根部泥土后測定根部形態和生長發育指標。主根長采用卷尺 (精度0.1 cm),根直徑采用數顯游標卡尺測定 (以蘆頭下1 cm為準),單株和單根鮮重采用電子秤測定 (精度0.01),最后計算根冠比。
根冠比(root shoot ratio)=單根鮮重/(單株鮮重-單根鮮重)
1.5 苗栽質量綜合評價
當歸苗栽綜合評價方法參照蘇李維等[15]的方法進行,即首先在主成分分析基礎上取初始特征根大于1的各指標主成分值,計算各指標的權重(Wj),然后計算隸屬函數值,最后估算綜合指數(comprehensive index, CI)。
Wj=∑(Cl,j×VPl)/∑∑(Cl,j×VPl)
R(Xij)=(Xij-Xjmin)/(Xjmax-Xjmin)
RR(Xij)=1-(Xij-Xjmin)/(Xjmax-Xjmin)
CIj=∑[R(Xij)×Wj]
式中:Cl,j表示第j個指標的第l主成分值;VPl表示第l主成分方差的百分率;Wj表示第j個指標的權重值;i表示不同試驗茬口;j表示不同指標;R(Xij)表示i茬口j指標的隸屬函數值;RR(Xij)表示i種類j指標的反隸屬函數值;Xij表示i茬口j指標的平均觀測值;Xjmin表示所有試驗茬口j指標的最小值;Xjmax表示所有試驗茬口j指標的最大值。CIj為第i茬口j個指標的累計綜合指數。
1.6 數據分析
采用Excel 2003制圖和t檢驗,采用SPSS 22.0 統計軟件進行方差分析和主成分分析,多重比較采用Duncan法,表中數據表示平均數±標準差,圖中數據點表示平均數±標準誤。
2 結果與分析
2.1 不同茬口條件下當歸苗數的動態變化
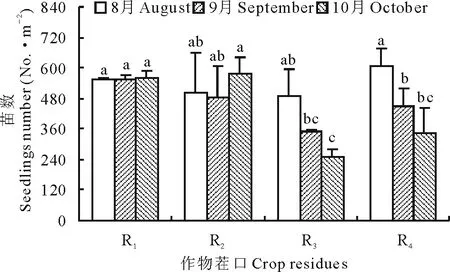
圖1 不同茬口對不同生長季當歸苗數動態變化的影響Fig.1 Effects of different crop residues on seedlings numberdynamics of A. sinensis in various stages R1:小麥茬Wheat residue;R2:黃芪茬Astragalus residue;R3:馬鈴薯茬Potato residue;R4:當歸茬Angelica residue.不同小寫字母表示差異顯著(P<0.05),下同。The different letters mean significance at P<0.05, the same below.
當歸種子6月5日播種后,8-10月每月中旬田間統計顯示(圖1),不同茬口對當歸種子出苗數和成苗數均具有顯著影響,影響程度隨茬口和生長季的不同而異。8月當歸重茬出苗數最多,分別較小麥茬、黃芪茬和馬鈴薯茬提高9.6%、21.0%和23.6%,馬鈴薯茬出苗數最低,但該時期各茬口當歸出苗數差異性均不顯著(F=0.212,P>0.05)。9月中旬各茬口因高溫干旱出現死苗現象,造成茬口間差異性增大,但小麥茬當歸苗數仍維持在8月出苗的水平,僅下降0.2% (P>0.05),黃芪茬次之,苗數較8月下降3.71% (P>0.05),而馬鈴薯和當歸重茬死苗最為嚴重,苗數分別下降28.7%和25.9%,但差異性均未達顯著水平(P>0.05)。9月后各茬口出現出苗和死苗動態交替變化,造成10月各茬口當歸苗數差異性達極顯著水平(F=2.346,P<0.05),小麥茬苗數仍維持穩定,由于種子尚零星出苗,苗數較9月提高1.45%(P>0.05),黃芪茬更為明顯,苗數較9月提高19.0% (P>0.05),馬鈴薯和當歸重茬死苗加重,分別較9月下降28.1%和23.1%,較8月下降48.8%(t=2.257,P<0.05)和43.0%(t=2.232,P<0.05) (圖1)。
從當歸開始出苗至育苗結束,各茬口坡度區組間的出苗差異性均未達到顯著水平(P>0.05),說明坡度對當歸育苗數的影響較小(圖1)。小麥茬和黃芪茬當歸出苗數隨生育時期的延后均表現為先降低后升高的趨勢,小麥茬當歸出苗數始終維持在穩定的水平,育成苗數較馬鈴薯和當歸重茬分別提高122.8%(t=8.072,P<0.05)和61.9%(P=0.05)。黃芪茬當歸種子遇適宜條件后出苗最為顯著,育成苗數較小麥茬提高2.6%(P>0.05),較馬鈴薯茬和當歸重茬提高128.6%(P<0.05)和66.2%(P<0.05)。而馬鈴薯和當歸重茬隨季節延后死苗嚴重,當歸苗數表現持續降低的趨勢,馬鈴薯茬當歸死苗程度更為顯著。
2.2 不同茬口對當歸苗栽地上部生長動態的影響
圖2顯示,不同茬口對當歸幼苗地上部生長發育動態具有顯著影響,影響程度隨茬口、測定指標和發育階段的不同而異。各茬口當歸幼苗地上部生長發育指標隨生長期延后呈基本一致的變化趨勢,均隨生育進程的推進先升高后降低,其中株幅在各月間差異達極顯著水平(P<0.01),葉片數達顯著水平(P<0.05)。8月中旬當歸種子出苗基本結束,各茬口當歸幼苗株高、株幅、真葉數從高到低均依次為黃芪茬>小麥茬>當歸重茬>馬鈴薯茬。莖粗有所不同,依次為當歸重茬>小麥茬>黃芪茬>馬鈴薯茬。至9月中旬各茬口苗栽進入生長盛期,當歸幼苗株高從高到低依次為小麥茬>當歸重茬>黃芪茬>馬鈴薯茬,株幅依次為當歸重茬>小麥茬>黃芪茬>馬鈴薯茬,真葉數依次為小麥茬>黃芪茬>當歸重茬>馬鈴薯茬,莖粗依次為黃芪茬>小麥茬>當歸重茬>馬鈴薯茬,馬鈴薯茬培育的當歸幼苗株高和株幅均顯著低于其他3個茬口的水平 (P<0.05),真葉數較小麥茬和黃芪茬顯著減少(P<0.05)。至10月,不同茬口條件下當歸幼苗生長表現出與生長旺盛期基本一致的趨勢,馬鈴薯茬生長指標始終處于最低水平,黃芪茬當歸苗最健壯,小麥茬次之,當歸重茬育成苗株幅最大,株型不緊湊。
2.3 不同茬口對當歸苗栽質量性狀的影響
圖3顯示,不同茬口對當歸種苗個體產量構成因素均具有較大影響,影響程度因指標的不同而異,其中對根長和根粗的影響達極顯著水平(P<0.01),對單株鮮重和單根鮮重的影響達顯著水平(P<0.05)。黃芪茬培育的當歸種苗根最長,小麥茬次之,二者較馬鈴薯茬分別極顯著增長66.6%(t=8.534,P<0.01)和46.4%(t=7.998,P<0.01),較當歸重茬分別極顯著增長54.9%(t=6.821,P<0.01)和36.1%(t=5.611,P<0.01),馬鈴薯茬種苗根長較當歸重茬縮短7.0%(P>0.05),根粗較當歸重茬降低28.2%(P>0.05)。小麥茬和黃芪茬培育的當歸種苗根最粗和次之,兩者差異性不顯著(P>0.05),均較馬鈴薯茬分別增粗51.0%(t=4.391,P<0.01)和48.1%(t=3.280,P<0.05),較當歸重茬分別增粗8.4%(t=0.530,P>0.05)和6.4%(t=0.372,P>0.05)。
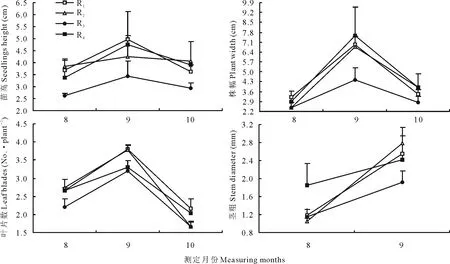
圖2 不同作物茬口對當歸苗栽地上部生長動態的影響Fig.2 Effects of different crop residues on aboveground growth dynamics of A. sinensis seedlings
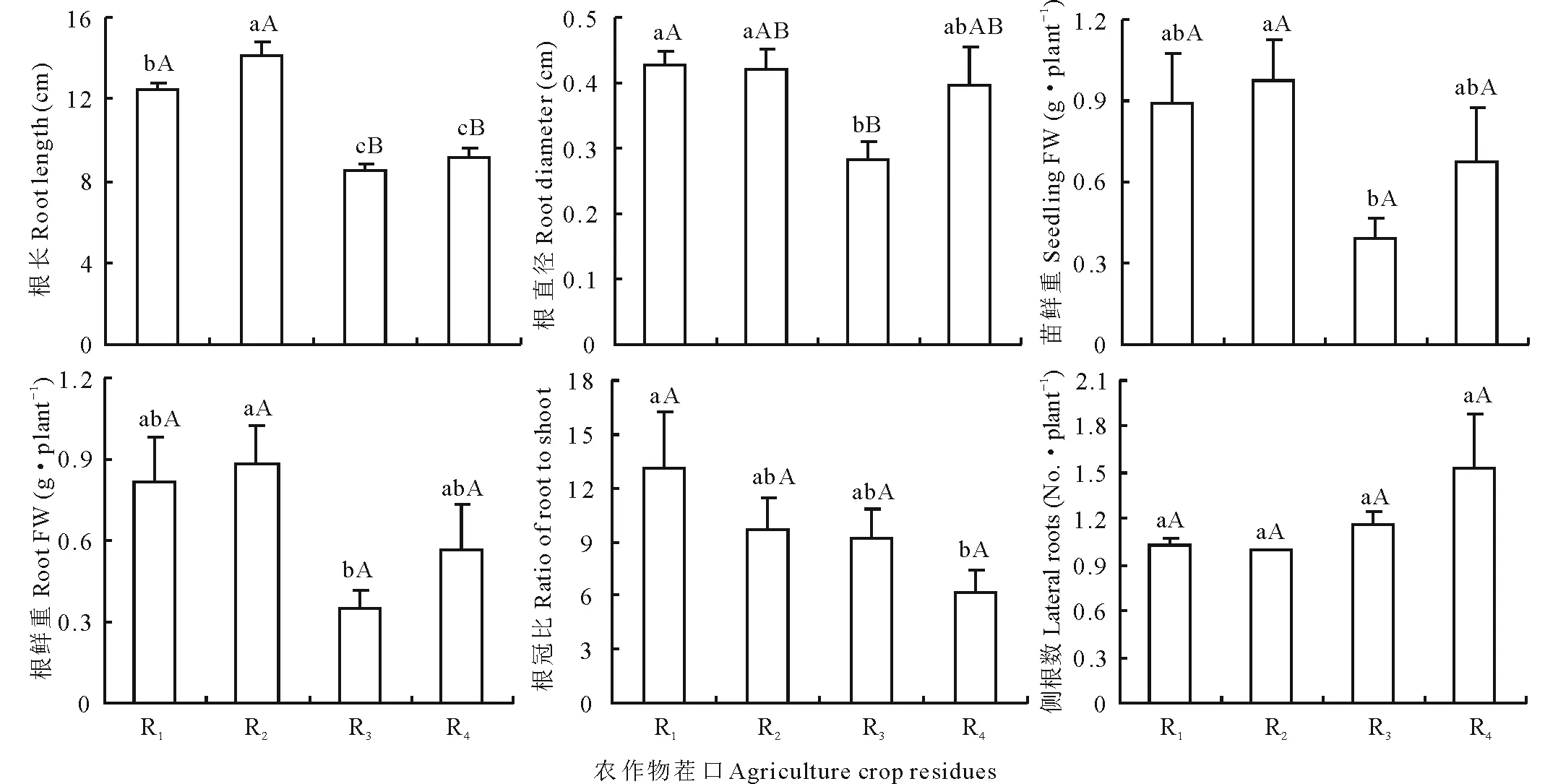
圖3 不同作物茬口對當歸苗栽質量性狀的影響Fig.3 Effects of different crop residues on quality indicators of A. sinensis seedlings 不同大寫字母表示差異極顯著(P<0.01),不同小寫字母表示差異顯著(P<0.05),下同。The different capital letters mean great-significance at P<0.01, the different small letters mean significance at P<0.05, The same below.
各茬口單苗鮮重和單根鮮重變化趨勢基本一致,個體產量從高到低依次為黃芪茬>小麥茬>當歸茬>馬鈴薯茬。小麥茬和黃芪茬單苗鮮重較馬鈴薯茬分別提高126.2%(t=2.518,P<0.05)和147.4%(t=3.562,P<0.05),較當歸重茬提高32.5%(t=1.318,P>0.05)和45.0%(t=1.859,P>0.05)。小麥茬和黃芪茬種苗單根鮮重較馬鈴薯茬分別提高132.3%(t=2.691,P<0.05)和150.0%(t=3.493,P<0.05),較當歸重茬分別提高44.4%和55.4%,黃芪茬當歸種苗個體產量指標更為優異。各茬口根冠比依次為小麥茬>黃芪茬>馬鈴薯茬>當歸茬,側根數正好相反,依次為當歸重茬>馬鈴薯茬>小麥茬和黃芪茬,小麥茬和黃芪茬苗栽側根數一致。茬口對根冠比的影響更大,與當歸重茬相比,小麥茬培育的當歸種苗根冠比提高112.9%(t=2.065,P=0.05),黃芪茬提高58.0%,馬鈴薯茬提高49.8%。
相關分析顯示(表1),根長和根粗均與單根鮮重呈極顯著正相關(P<0.01)。株高與根長和側根數呈顯著正相關(P<0.05),與株幅、根粗、單株鮮重和單根鮮重均呈極顯著正相關(P<0.01)。株幅與根粗、單苗鮮重均呈極顯著正相關(P<0.01),與單根鮮重呈顯著正相關(P<0.05)。
2.4 不同茬口對當歸苗栽根病發病率和病情指數的影響
對不同茬口培育的當歸種苗田間病情統計顯示(圖4),不同茬口培育的當歸種苗發病率和病情指數變化趨勢一致,當歸重茬育成種苗發病率和病情指數最高,在小麥茬、黃芪茬和馬鈴薯茬進行當歸育苗可在一定程度上降低種苗的發病率和病情指數,降低程度隨前茬作物的不同差異性較大。與當歸重茬相比,小麥茬培育的當歸種苗發病率和病情指數極顯著降低54.6%(t=5.367,P<0.01)和64.9%(t=4.724,P<0.01),黃芪茬和馬鈴薯茬發病率分別降低45.6%(P<0.01)和18.2%,病情指數分別顯著降低64.1%(t=6.934,P<0.01)和41.0%(t=4.00,P<0.05),不同茬口育成苗發病率和病情指數從低到高依次為小麥茬<黃芪茬<馬鈴薯茬<當歸重茬。
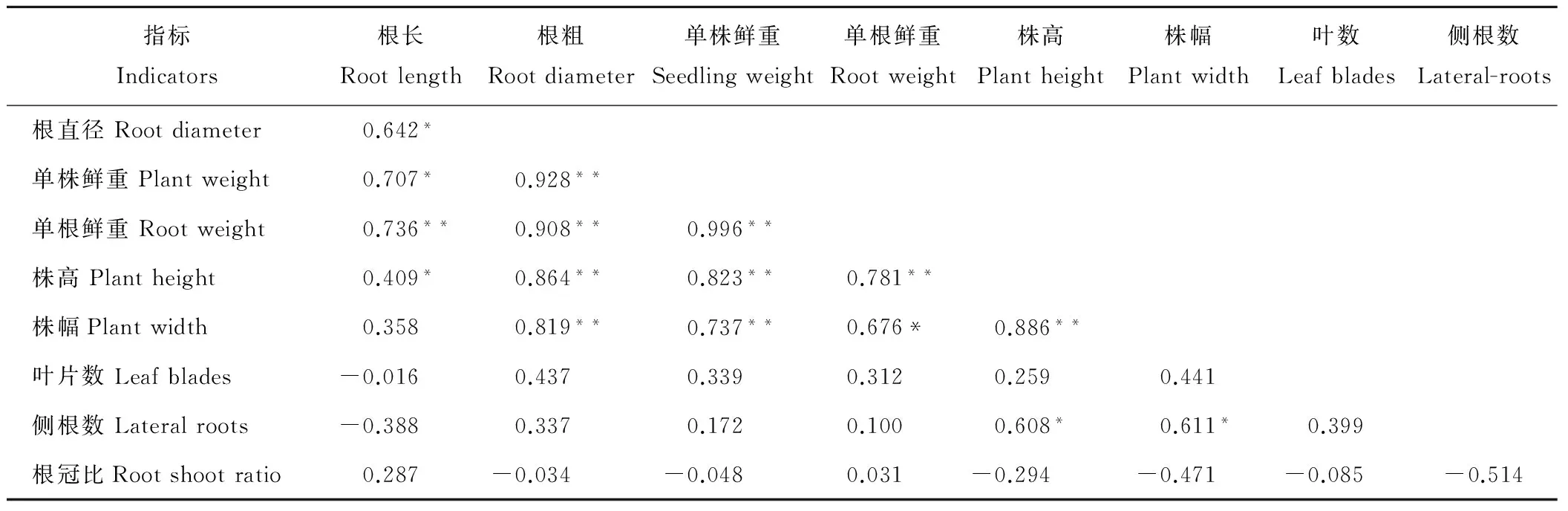
表1 當歸種苗個體質量指標間的相關性分析Table 1 Correlation analysis among individual quality indicators of A. sinensis seedlings
注: **表示相關性達極顯著水平(P<0.01),*表示相關性達顯著水平(P<0.05),以作物茬口每個重復小區中10株個體的平均值作為相關分析數據(n=12,R0.05, df=10=0.576,R0.01, df=10=0.708)。
Note: ** mean correlation was great-significance atP<0.01, * mean correlation was significance atP<0.05, using the mean of 10 single plants in each replicated plots as the correlation analysis data (n=12,R0.05, df=10=0.576,R0.01, df=10=0.708).
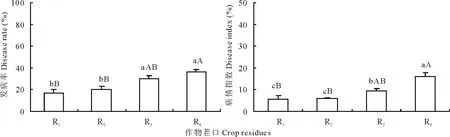
圖4 不同茬口對當歸種苗發病率和病情指數的影響Fig.4 Effects of different crop residues on disease ratio and index of A. sinensis seedlings
2.5 不同茬口當歸苗栽質量性狀的綜合分析
經對各茬口苗栽性狀指標主成分分析顯示(表2),其中前3個主成分的特征根均大于1,累積貢獻率達100%,故提取第1~3主成分值的特征根和貢獻率計算各指標的權重值(表3)。根據各指標隸屬度值與權重,通過加乘法則,對各指標進行合成運算,對各茬口當歸種苗綜合評價指數估算表明,不同茬口培育的當歸種苗綜合指數大小依次為黃芪茬>小麥茬>當歸重茬>馬鈴薯茬(表4)。
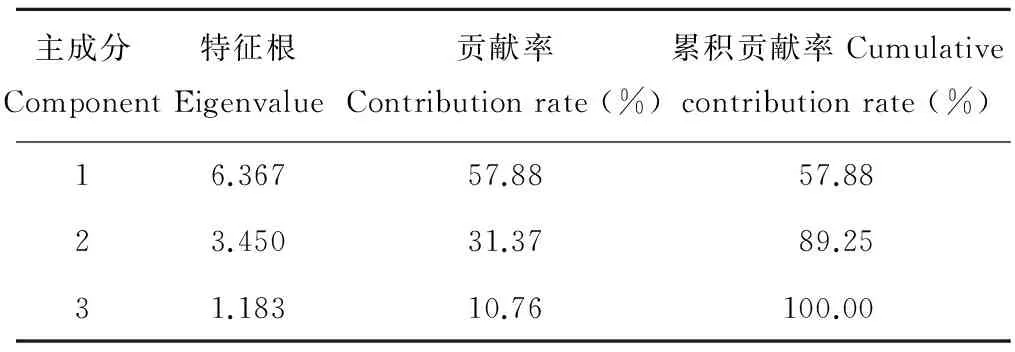
表2 當歸種苗質量性狀指標主成分分析Table 2 Principal components analysis of characteristic indicators of A. sinensis seedlings
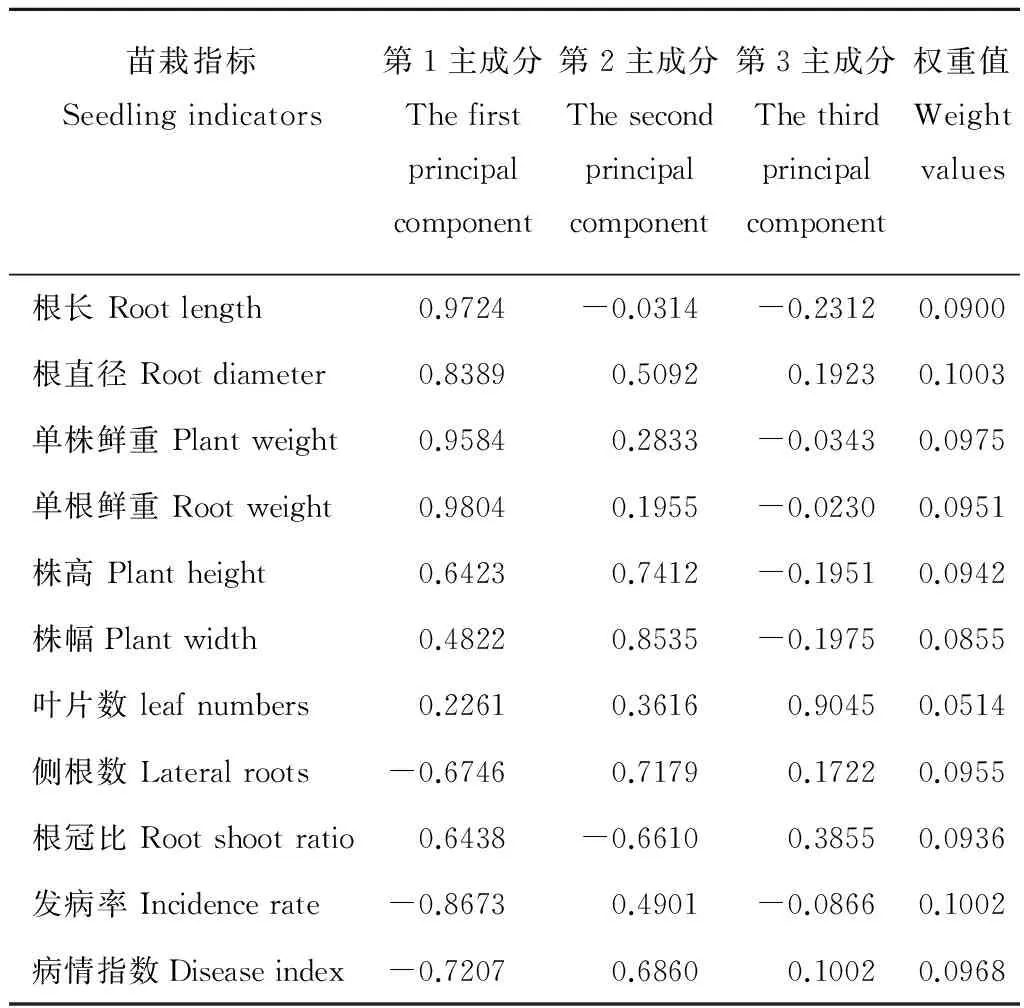
表3 當歸種苗性狀指標的負荷量和權重Table 3 Capacity and weight of characteristic indicators of A. sinensis seedlings
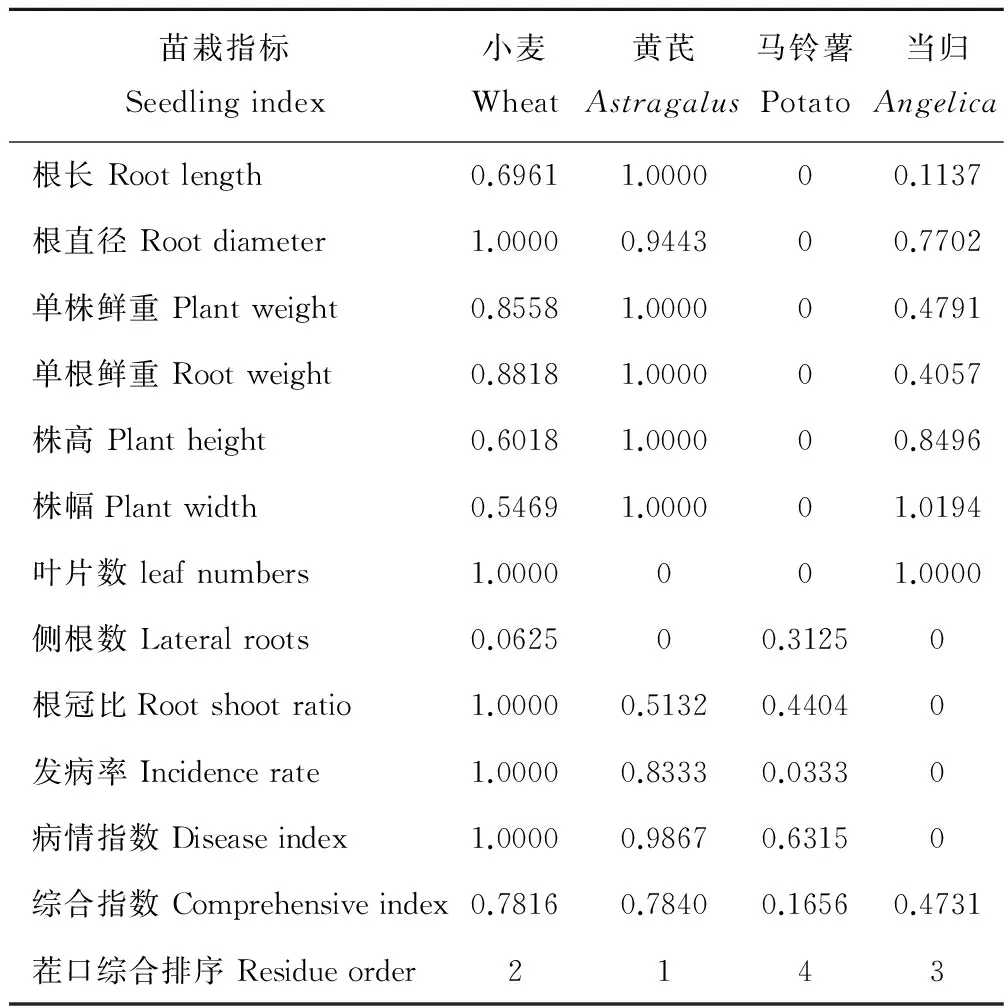
表4 當歸種苗性狀指標的隸屬度值及綜合評價指數Table 4 Subordination value of seedling characteristic indicators and comprehensive evaluation index of A. sinensis seedlings
3 討論
3.1 不同作物茬口對當歸成苗數具有顯著影響
連作障礙是限制作物生產最主要的因素之一,尤其在藥用植物上表現更為突出[16]。連作易引起土壤養分匱乏、土壤理化性質惡化和土壤微生物多樣性的變化等[17-18]。輪作倒茬是一種緩解土壤環境惡化的有效措施。一般來說,倒茬可顯著提高土壤酶活性和速效養分含量并豐富微生物多樣性,進而提高土壤肥力[19-20]。茬口形成的因素主要有作物吸收養分和水分、根系、落葉、根際微生物、伴隨作物發生的雜草病蟲害、耕作栽培技術對土壤的影響和作物向土壤分泌物質等,茬口特性的好壞最終體現在后茬作物的生長發育和產量上[6]。徐雪風等[21]研究表明,輪作油葵對馬鈴薯生長發育及抗性具有顯著影響。本研究在岷縣熟地培育小麥、黃芪、馬鈴薯和當歸茬口基礎上進行當歸育苗,田間定期統計發現,梯田坡度對當歸育苗的影響較小,而茬口對當歸出苗和成苗的影響隨生長季延后差異性增大。當歸重茬出苗勢最高,馬鈴薯茬最低,小麥茬和黃芪茬居中,但各茬口間差異性不顯著,隨著生長季延后田間出現死苗和出苗交替現象,當歸重茬和馬鈴薯茬死苗率達40%以上,而小麥茬和黃芪茬死苗率均低于5%,黃芪茬遇適宜條件還有出苗勢頭,造成各茬口育成當歸苗數差異性增大,成苗數依次為黃芪茬>小麥茬>當歸茬>馬鈴薯茬,小麥茬和黃芪茬當歸成苗數多,保苗率高,較馬鈴薯和當歸重茬分別提高120%和60%以上,為提高種苗產量奠定了數量基礎,可能是由于小麥茬和黃芪茬土壤有機質含量高,土壤比熱容大,保墑性能好,對土壤溫度具有一定的調控能力的緣故,因為2016年當歸播種出苗后正直生長期,遇持續高溫干旱天氣,小麥茬和黃芪茬造就了良好的土壤環境條件,這也印證了溫健[22]和馬偉明[23]采用盆栽育苗研究的結論,即不同茬口土壤育成當歸苗根際土壤肥力存在顯著差異,黃芪茬口堿性較弱,速效鉀含量高,微生物多樣性豐富,當歸苗抗逆性強,有利于當歸苗的生長發育。
3.2 不同茬口對當歸育成苗栽產量性狀具有顯著影響
連作易引起土壤有害微生物的富集,導致病蟲草害的爆發,降低作物生產力。輪作油葵可顯著改善馬鈴薯連作田土壤環境,促進馬鈴薯地上部生長發育,提高單株產量[21]。Zhao等[24]研究表明,輪作花生可顯著促進燕麥植株生長,提高產量。Graham[25]認為較高的根冠比是作物獲得高產的重要特征。對于藥用植物而言,獲得高產的關鍵取決于根系的發育狀況[26]。Xu等[27]研究指出,適當的干旱土壤環境可以顯著提高水稻的根冠比,增強抗旱性。當歸是具有明顯自毒作用的藥用植物[9]。傳統上當歸采用生荒地育苗,熟地栽培的方式。當歸苗栽采挖后以根越冬,根部性狀決定了種苗的產量和質量。馬偉明等[13]采集岷縣4種茬口土壤進行當歸盆栽育苗發現,黃芪茬口土壤當歸根系發達,生長量與生荒地相當,根系活力更強,具有潛在的抗逆基礎,而馬鈴薯茬和當歸重茬口土壤當歸苗長勢弱,根系活力低。張新慧等[11]研究表明,馬鈴薯-小麥-當歸輪作栽培促進了當歸根系生長,產量和品質均較當歸連作顯著提高。本研究中,小麥茬和黃芪茬均顯著提高了當歸種苗根長,在促進根系生物量積累的同時,改善了地上部生物量積累,表現為葉片數的增加和根冠比的提高,培育的種苗根系較發達,為抗旱抗熱奠定了物質和能量基礎。相關分析表明,茬口對當歸種苗產量構成因素的影響主要體現在株高、株幅、葉片數、主根長、單根鮮重和根冠比等指標。馬鈴薯茬和當歸重茬種苗根系生長受到抑制,易形成老小苗,側根多,地上部生長勢弱,可能是由于馬鈴薯茬和當歸重茬土壤中有機養分含量較差,根系分泌的化感物質成分復雜[12],而單子葉植物小麥具有獨特的凝集素和酶系統,可以促進其根際土壤中有機質的演化和化感物質的降解程度[28],豆科植物黃芪可以改善根際微生物多樣性[22]的緣故。
3.3 不同茬口對當歸育成苗栽抗病性具有顯著影響
植物經常會受到土壤、溫度、濕度等生存環境因子的影響,這些環境因子的變化有時非常劇烈,超過了植物正常生長所能忍受的范圍,從而導致植物受到傷害,抗逆性強的植物受害程度較輕[29]。當歸根腐病是限制當歸生產的主要因素[30]。發病率和病情指數是衡量病情和程度的主要指標,合理輪作能有效抑制病原微生物的繁殖和傳播[31]。黃亞萍[30]經對當歸根腐病發病部位病原物分離鑒定表明,當歸根腐病主要致病菌有茄病鐮刀菌(Fusariumsolani)、尖孢鐮刀菌(F.oxysporum)和芬芳鐮刀菌(F.redolens),主要通過傷口侵染,持續光照產孢量最大,持續黑暗利于菌絲生長,生長最適溫度25~30 ℃。當歸是典型喜涼藥用植物,適宜高寒陰濕的環境條件,傳統一般選用高海拔區生荒地育苗,熟地育苗受到高溫和干旱的威脅,2016年7-8月高溫干旱嚴重,本研究在熟地采用4種作物茬口育苗,苗栽采挖期田間觀察到,當歸種苗根腐病表現為根部出現大小不等的網狀斑和小黑孔,呈紅褐色至黑褐色凹陷斑,嚴重時腐爛,一般發病較輕的幼苗地上部無明顯癥狀,但發病重的幼苗生長勢弱,造成根莖腐爛,這與黃亞萍[30]研究的結果相吻合。本研究還發現,輪作換茬可有效抑制后茬育成當歸種苗的發病率和病情指數,其中小麥茬和黃芪茬當歸種苗受病原微生物感染程度低,種苗質量優異,當歸重茬培育的種苗發病率和病情指數均處于最高水平,說明當歸重茬種苗抗病能力較差,同時突顯出當歸連作障礙問題的嚴重性,這與劉星等[16]對馬鈴薯連作的研究結果相似,可能是由于當歸重茬蟲害發生嚴重,病原菌富集,造成根莖部傷口,病原菌侵染所致的緣故。
4 結論
植物在受到逆境脅迫而引起體內生理生化指標變化的因素較為復雜[15],運用單一指標很難反映當歸種苗抗性的好壞。為了全面準確地評判各茬口育成當歸種苗的優劣,對多個生長發育指標和抗病指標運用主成分分析結合隸屬度綜合分析顯示,不同茬口當歸種苗優劣次序為黃芪茬>小麥茬>當歸重茬>馬鈴薯茬。采用熟地當歸育苗,小麥茬和黃芪茬當歸成苗率高,生長勢強,苗栽健壯優異,產量高,抗病性強。馬鈴薯和當歸重茬當歸苗栽生長受季節影響較大,抗性較差,死苗率高,種苗易形成側根數較多的老小苗,更易受病菌感染,馬鈴薯茬死苗率最高,當歸茬根腐病發生更為嚴重,說明正茬黃芪和小麥茬口是適宜當歸熟地育苗的床土資源。下一步需對不同茬口育成種苗進行移栽,對藥材產量和質量進一步鑒定比較研究。
致謝:
岷縣禾馱鄉郎銀忠提供試驗地和幫助,隴西縣郭志軍,郭太平、牟小紅、趙巧云、馬建雄參與試驗栽培管理。甘肅農業大學博士生安志剛、姜小鳳,碩士生周傳猛、郭一青、王小琴,蘭州大學碩士生郭愛峰,蘭州財經大學長青學院本科生郭愛偉參與作物茬口培育及育苗試驗,本科生許亞強、嚴涇國、張海龍和張軍剛參與種苗測定,在此一并致謝。
參考文獻References:
[1] State Pharmacopoeia Commission. People’s Republic of China Pharmacopoeia (one). Beijing: China Medical Science Press, 2015: 36-38.
國家藥典委員會. 中華人民共和國藥典(一部). 北京: 中國醫藥科技出版社, 2015: 36-38.
[2] Zhao R M, Chen Y, Guo F X,etal. Comparative study on distributed feature of wildAngelicasinensisresources and the difference in growth characteristics with its cultivars native to Min County of Gansu. Acta Prataculturae Sinica, 2014, 23(2): 29-37.
趙銳明, 陳垣, 郭鳳霞, 等. 甘肅岷縣野生當歸資源分布特點及其與栽培當歸生長特性的比較研究. 草業學報, 2014, 23(2): 29-37.
[3] Zhang G X, Li J H.Angelica. Beijing: China Agricultural Publishing Press, 1989.
張廣學, 李靜華. 當歸. 北京: 農業出版社, 1989.
[4] Meng Y, Guo Z X. The investigation and thoughts onAngelicasinensisseedling in Min county. Gansu Agricultural Science and Technology, 2008, (15): 38-39.
孟玉, 郭增祥. 岷縣當歸育苗的調查與思考. 農業科技與信息, 2008, (15): 38-39.
[5] Zhou H J, Wang X, Yuan Y,etal. Rapid-assessing methods of loss in extremely heavy rainfall disaster chain in semiarid region—A case study on a flash flood debris flow in Minxian County, Gansu Province. Arid Zone Research, 2014, 31(3): 440-445.
周洪建, 王曦, 袁藝, 等. 半干旱區極端強降雨災害鏈損失快速評估方法——以甘肅岷縣“5·10”特大山洪泥石流災害為例. 干旱區研究, 2014, 31(3): 440-445.
[6] Wang L X, Li J. Farming science. Beijing: Science Press, 2003: 278-279.
王立祥, 李軍. 農作學. 北京: 科學出版社, 2003: 278-279.
[7] Tian F P, Shi S L, Hong F Z,etal. Research on history and current situation of forage and crop rotation in China. Pratacultural Science, 2012, 29(2): 320-326.
田福平, 師尚禮, 洪紱曾, 等. 我國草田輪作的研究歷史及現狀. 草業科學, 2012, 29(2): 320-326.
[8] Peng Y, Zhao Z X, Li Z H,etal. Effects of different preceding crops on growth, yield and quality of flue-cured tobacco plant. Acta Agronomica Sinica, 2010, 36(2): 335-340.
彭云, 趙正雄, 李忠環, 等. 不同前茬對烤煙生長、產量和質量的影響. 作物學報, 2010, 36(2): 335-340.
[9] Wang T T, Wang Q, Wang H Z,etal. Effects of intercropping patterns on growth characters and yield ofAngelicasinensisunder continuous mono-cropping planting. Acta Prataculturae Sinica, 2013, 22(2): 54-61.
王田濤, 王琦, 王惠珍, 等. 連作條件下間作模式對當歸生長特性和產量的影響. 草業學報, 2013, 22(2): 54-61.
[10] Wang H Z, Zhang X H, Li Y D,etal. Comparison of photosynthetic characteristic and the essential oils in crop rotation and continuous cropping ofAngelicasinensis. Acta Prataculturae Sinica, 2011, 20(1): 69-74.
王惠珍, 張新慧, 李應東, 等. 輪作與連作當歸光合特性和揮發油的比較. 草業學報, 2011, 20(1): 69-74.
[11] Zhang X H, Zhang E H, Wang Y J,etal. Effects of different stubble on yield and quality ofAngelicasinensis. Journal of Chinese Medicinal Materials, 2010, 33(11): 1678-1680.
張新慧, 張恩和, 王永捷, 等. 不同前茬對當歸藥材產量及品質的影響. 中藥材, 2010, 33(11): 1678-1680.
[12] Wang T T, Wang Q, Wang H Z,etal. Autotoxicity ofAngelicasinensisand allelopathy on tested plants. Acta Agrestia Sinica, 2012, 20(6): 1132-1138.
王田濤, 王琦, 王惠珍, 等. 當歸自毒作用和其對不同作物的化感效應. 草地學報, 2012, 20(6): 1132-1138.
[13] Ma W M, Guo F X, Chen Y,etal. Study on the seedling ofAngelicasinensison soil from different stubbles. China Journal of Chinese Materia Medica, 2009, 34(5): 552-553.
馬偉明, 郭鳳霞, 陳垣, 等. 在不同茬口土地上的當歸育苗研究. 中國中藥雜志, 2009, 34(5): 552-553.
[14] Chen Y, Zhu L, Guo F X,etal. Isolation and identification of the pathogens causingAstragalusmembranaceusvar.mongholicusroot rot in Weiyuan of Gansu Province. Acta Phytopathologica Sinica, 2011, 41(4): 428-431.
陳垣, 朱蕾, 郭鳳霞, 等. 甘肅渭源蒙古黃芪根腐病病原菌的分離與鑒定. 植物病理學報, 2011, 41(4): 428-431.
[15] Su L W, Li S, Ma S Y,etal. A comprehensive assessment method for cold resistance of grape vines. Acta Prataculturae Sinica, 2015, 24(3): 70-79.
蘇李維, 李勝, 馬紹英, 等. 葡萄抗寒性綜合評價方法的建立. 草業學報, 2015, 24(3): 70-79.
[16] Liu X, Zhang S L, Liu G F,etal. Effects of continuous cropping on dry matter accumulation and distribution of potato plants in the yellow river irrigation areas of middle Gansu province. Acta Agronomica Sinica, 2014, 40(7): 1274-1285.
劉星, 張書樂, 劉國鋒, 等. 連作對甘肅中部沿黃灌區馬鈴薯干物質積累和分配的影響. 作物學報, 2014, 40(7): 1274-1285.
[17] Sun X T, Li L, Long G Q,etal. The progress and prospect on consecutive monoculture problems ofPanaxnotoginsen. Chinese Journal of Ecology, 2015, 34(3): 885-893.
孫雪婷, 李磊, 龍光強, 等. 三七連作障礙研究進展. 生態學雜志, 2015, 34(3): 885-893.
[18] Zhang W, Long X, Huo X,etal. 16S rRNA-based PCR-DGGE analysis of actinomycete communities in fields with continuous cotton cropping in Xinjiang, China. Microbial Ecology, 2013, 66(2): 385-393.
[19] Marcinkeviciene A, Boguzas V, Balnyte S,etal. Influence of crop rotation, intermediate crops, and organic fertilizers on the soil enzymatic activity and humus content in organic farming systems. Eurasian Soil Science, 2013, 46(2): 198-203.
[20] Maiti D, Singh R K, Variar M. Rice-based crop rotation for enhancing native arbuscular mycorrhizal (AM) activity to improve phosphorus nutrition of upland rice (OryzasativaL.). Biology & Fertility of Soils, 2012, 48(1): 67-73.
[21] Xu X F, Li C Z, Zhang J L. Effects of oil-sunflower rotation on growth and resistance physiology indexes of potato. Soils, 2017, 49(1): 83-89.
徐雪風, 李朝周, 張俊蓮. 輪作油葵對馬鈴薯生長發育及抗性生理指標的影響. 土壤, 2017, 49(1): 83-89.
[22] Wen J. Studies on phylogenetic diversity of unculturable bacterias fromAngelicasinensisrhizosphere under different residues. Lanzhou: Gansu Agricultural University, 2008.
溫健. 不同茬口當歸根際微生物多樣性研究. 蘭州: 甘肅農業大學, 2008.
[23] Ma W M. Effect of different residues on seedlings vulturing ofAngelica. Lanzhou: Gansu Agricultural University, 2008.
馬偉明. 不同茬口對當歸育苗的影響. 蘭州: 甘肅農業大學, 2008.
[24] Zhao D, Wright D L, Marois J J,etal. Improved growth and nutrient status of an oat cover crop in sod-based versus conventional peanut-cotton rotations. Agronomy for Sustainable Development, 2010, 30(2): 497-504.
[25] Graham R D. Breeding for nutritional characteristics in cereals. Novartis Foundation Symposium, 2001, 236: 205.
[26] Zhang T T, Chen Y, Guo F X,etal. Effect of different sowing quantities on seedling quality ofRheumpalmatum. Acta Prataculturae Sinica, 2013, 22(4): 99-105.
張天銅, 陳垣, 郭鳳霞, 等. 不同播種量對甘肅禮縣掌葉大黃育苗質量的影響. 草業學報, 2013, 22(4): 99-105.
[27] Xu W, Cui K, Xu A,etal. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiologiae Plantarum, 2015, 37(2): 1-11.
[28] Bezrukova M V, Aval’Baev A M, Kil’Dibekova A R,etal. Interaction of wheat lectin with 24-Epibrassinolide in the regulation of cell division in wheat roots. Doklady Biological Sciences, 2002, 387: 533-535.
[29] Guo F X, Zhang M X, Chen Y,etal. Relation of several antioxidant enzymes to rapid freezing resistance in suspension cultured cells from alpineChorisporabungeana. Cryobiology, 2006, 52(2): 241-250.
[30] Huang Y P. Studies on the pathogens causingAngelicasinensisroot. Lanzhou: Gansu Agricultural University, 2009.
黃亞萍. 當歸根腐病病原物研究. 蘭州: 甘肅農業大學, 2009.
[31] Sun Y C, Chen J T, Guo L P,etal. Control of medicinal plant soil-borne diseases by crop rotation: A Review. Modern Chinese Medicine, 2012, 14(10): 37-41.
孫躍春, 陳景堂, 郭蘭萍, 等. 輪作用于藥用植物土傳病害防治的研究進展.中國現代中藥, 2012, 14(10): 37-41.
