Apaf-1通過wnt/β-catenin信號通路調控肝癌的機制研究*
2018-01-29 03:16:32郭善軍楊宏靜
重慶醫學 2018年3期
郭善軍,李 晶,楊宏靜△
(1.達州職業技術學院,四川達州 635001;2.重慶三峽醫藥高等專科學校 404120)
肝癌是最常見的惡性腫瘤之一,其全球發病率位居癌癥第6位,且治療難、預后差、生存率低,因此研究肝癌發病機制對肝癌的治療具有指導作用[1]。近年來,wnt介導經典的wnt/β-catenin信號通路受到人們的廣泛關注,wnt/β-catenin信號通路不僅對脊椎動物的胚胎發育及組織穩態具有重要功能,而且腫瘤的形成與wnt/β-catenin信號通路異常密切相關。該系統的激活可引起β-catenin 在胞漿中的積聚,進而進入胞核與TCF/LEF轉錄因子結合,激活下游基因的表達。在沒有激活的情況下,β-catenin與人糖原合成酶激酶3β(GSK3β)、Axin、抗原呈遞細胞(APC)、蓬亂蛋白1(DVL-1)和蛋白磷酸酶2A(PP-2A)相結合,β-catenin被磷酸化,進而被蛋白水解酶所降解。wnt信號被激活時,GSK3β的活性被DVL-1所抑制,引起β-catenin 的積聚[2]。
凋亡酶激活因子(apoptotic protease activating facter-1,Apaf-1)是一種新近發現的抑癌基因,參與人體線粒體凋亡途徑,作為一種多功能結構域的構架蛋白,通過與wnt/β-catenin信號通路中的其他蛋白分子結合發揮生物學作用[3]。研究證明,Apaf-1在各種惡性腫瘤中都存在啟動子高甲基化水平導致其自身表達降低,最終導致wnt/β-catenin信號通路異常引起惡性腫瘤的發生[4]。目前國內外關于Apaf-1在肝癌細胞中的表達及作用機制報道缺乏,因此對于研究Apaf-1基因肝癌作用機制的研究十分必要。本實驗通過驗證Apaf-1對wnt/β-catenin信號通路的調控、Apaf-1在肝癌細胞中表達和敲降Apaf-1對肝癌細胞的影響來探討Apaf-1在肝癌細胞中的作用機制,為尋找新的肝癌治療靶點及防治藥物提供理論依據。
1 材料與方法
1.1材料 肝癌細胞株HepG2、HHCC、HB611及正常肝細胞株LO2購自上海奧陸生物科技有限公司,pcDNA3.1質粒、HEK293細胞株來自西南大學淡水魚類資源與生殖發育教育部重點實驗室饋贈,雙熒光素酶檢測試劑盒(Dual-Luciferase Reporter Assay System)購自美國Promega 公司(Promega,USA),RNA提取(RNAiso Plus)、反轉錄試劑盒(PrimeScript RT reagent Kit)和定量PCR試劑盒均來自日本TaKaRa公司,pCDH質粒購自維真生物;抗體購自Bioword公司。

表1 實時定量PCR相關引物
1.2方法
1.2.1驗證Apaf-1基因對wnt/β-catenin信號通路調控作用 為了驗證Apaf-1基因對wnt/β-catenin信號通路具有調控作用,本研究將人的Apaf-1、β-catenin基因全長cDNA克隆連接到pcDNA3.1質粒,通過細胞轉染至HEK293細胞株,轉染過程每孔加入0.50 μg TOPFlash報告質粒(判斷wnt/β-catenin信號通路是否激活的標志)、0.05~0.50 μg包含Apaf-1、β-catenin全長的pcDNA3.1質粒、100 ng/孔的pRL-TK(pRL-TK表達的海腎熒光素酶作為一個內參因子可以用來判斷轉染效率,并計算相對熒光素酶活性),以未進行轉染和轉染空質粒為空白對照組和陰性對照組,每組3個復孔;轉染后48 h收細胞:磷酸緩沖鹽溶液(PBS)洗3次,每孔加入100 μL裂解液,室溫裂解10 min。用Dual-Luciferase Reporter Assay System試劑盒在在Luminosakan Ascent化學發光分析儀上測定熒光素酶活性[5]。
1.2.2實時熒光定量PCR檢測肝癌細胞株及正常肝細胞株中Apaf-1的表達 取對數生長期HepG2、HHCC、HB611及LO2細胞株,用含10% 胎牛血清的DMEM 培養,然后提取各個細胞株總RNA,反轉錄cDNA,通過實時定量PCR方法檢測Apaf-1的表達,以β肌動蛋白(β-actin)為內參基因。使用Primer Express 軟件(Applied Biosystems,USA)設計定量PCR引物,并遵循至少一個引物跨不同外顯子從而避免基因組DNA污染,見表1。使用目的基因不同濃度梯度(102~108拷貝)的質粒作為模板檢測引物的擴增效率[6-7]。
1.2.3HepG2細胞株中基因敲降Apaf-1 取對數生長期HepG2細胞株,調整細胞密度6×105/mL,接種至6孔板,每孔4 mL,用10%胎牛血清的DMEM 培養,將構建好的Apaf-1 shRNA pCDH質粒按Lipofectamine2000說明書的方法轉染到HepG2細胞中,對照組轉染陰性空載質粒,每組設置3個復孔。
1.2.4轉染效率 將1.2.3中培養的細胞株在36 h后放入熒光顯微鏡下觀察,轉染陽性細胞可看到熒光,陰性或者正常細胞不可見熒光。
1.2.5實時定量PCR儀檢測相關基因表達量及抑制效率 將1.2.3中培養的細胞株在48 h后收集,按照常規方法提取細胞總RNA,反轉錄成cDNA。以cDNA為模板,β-actin為內參基因進行實時定量PCR反應檢測細胞周期蛋白A(Cyclin A)、細胞周期素依賴性激酶2(CDK2)、減數分裂相關蛋白(p-CDK2)、β-catenin、APC、p-STAT3、p-STAT5、STAT3、STAT5、PRR11基因的表達。
1.2.6Wstern blot檢測相關蛋白的表達量 將1.2.2中培養的細胞株在48 h后收集,按照常規方法提取細胞內總蛋白,用紫外分光光度計測定吸光值分析總蛋白純度,稀釋各組蛋白的濃度至終濃度一致,每組取50 μg蛋白樣品,進行SDS-PAGE電泳,然后將蛋白轉移至PVDF膜上,5%牛血清白蛋白封閉2 h,加一抗,室溫孵育4 h,滴加二抗,室溫孵育2 h,利用化學發光凝膠成像儀進行成像[8]。
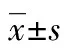
2 結 果
2.1Apaf-1可抑制wnt/β-catenin介導的信號通路 用Super 8×TOPFlash報告因子研究Apaf-1對β-catenin介導的經典wnt信號通路的調控作用。結果發現Apaf-1可以抑制β-catenin激活的LEF熒光素酶活性,并且這種作用具有劑量依賴效應,見圖1。

A:TOPFlash熒光素酶表達結果;B:TOPFlash熒光素酶劑量效應結果;a:P<0.05
圖1 TopFlash-熒光素酶報告實驗
2.2各細胞株中Apaf-1的表達 Apaf-1在正常的肝細胞株LO2的表達量明顯高于肝癌細胞株中的表達量,且在HepG2細胞株中的表達量最低,說明HepG2細胞株對Apaf-1的更敏感,差異均具有統計學意義(P<0.05),見圖2。

a:P<0.05
圖2定量PCR檢測Apaf-1在不同細胞株中的表達
2.3基因敲降轉染效率 在RNA干擾的HepG2細胞株中,與對照組相比,實驗組可以觀察到明顯的熒光信號,且熒光信號較多,而對照組無明顯熒光信號,見圖3。

A、C實驗組;B、D對照組
圖3根據同一視野下標記有熒光信號細胞的多少計算轉染效率(×100)
2.4Real-timePCR檢測相關基因的表達 在RNA干擾Apaf-1的HepG2細胞株中,與正常的HepG2細胞株相比,Apaf-1的表達量降低,說明在HepG2細胞中,Apaf-1敲降成功,同時wnt/β-catenin信號通路的相關基因或者肝癌相關基因的表達顯著升高或者降低,如β-catenin、Cyclin A、CDK2、wnt5a、STAT3、EGFR、APC(P<0.05),見圖4。

siRNA:在HepG2細胞株中敲降Apaf-1基因;a:P<0.05
圖4定量PCR分析在HepG2細胞株中敲降Apaf-1基因后相關基因表達量變化
2.5Wstern blot檢測相關蛋白表達 在RNA干擾Apaf-1的的HepG2細胞株中,與正常的HepG2細胞株相比,β-catenin、Cyclin A、CDK2的表達量顯著升高,Apaf-1、APC的表達量降低(P<0.05),見圖5。

siRNA:在HepG2細胞株中敲降Apaf-1基因
圖5 Western blot檢測HepG2和siRNA-Apaf-1 HepG2中相關蛋白的表達
3 討 論
近年來,我國肝癌的發病率呈上升趨勢,且術后復發率高,傳統的輔助治療、放化療等治療研究緩慢,都嚴重影響了肝癌患者的生存率。尋找更有效的診斷和治療方法的首要步驟就是要了解其病理學分子機制。近年隨著醫學分子生物學、細胞生物學快速發展,腫瘤的治療開始向細胞及基因水平發展[9]。越來越多研究證明,肝癌的發生與體內癌基因的過表達及抑癌基因的表達缺失相關[10]。因此,研究這些癌基因或抑癌基因的功能對于肝癌的治療具有重要指導作用。
Apaf-1作為一種新發現的多功能結構域蛋白,它通過調控wnt/β-catenin信號通路的異常激活,促進靶基因的過度表達,影響細胞的增殖與分化,促進腫瘤的發生,因此,Apaf-1屬于wnt/β-catenin信號通路的負調控因子[11-12]。本研究通過TopFlash-熒光素酶報告實驗檢測發現,當加入50 ng Apaf-1質粒時,熒光信號減弱,并且隨著Apaf-1濃度的增加,檢測到的熒光信號逐漸減弱。這些結果揭示了Apaf-1能夠抑制wnt/β-catenin信號通路活性,且具有劑量依賴效應。
在Wnt介導的四種通路中經典的Rspo1/Wnt/β-catenin信號通路最受關注,因為經典信號通路在哺乳動物的發育過程中發揮著舉足輕重的作用,例如器官再生、性別分化和腫瘤形成[7-8]。在正常的肝細胞中,β-catenin作為一種細胞骨架蛋白在胞膜處與鈣黏蛋白(E-cadherin)形成復合體對維持同型細胞的黏附、防止細胞移動發揮作用,而在肝癌細胞中,β-catenin與由APC和Apaf-1組成的復合物減少,從而影響wnt/β-catenin信號通路異常,導致肝癌細胞的構像發生改變[10]。本研究利用PCR檢測肝癌細胞株Apaf-1的表達明顯低于正常肝細胞株,驗證了Apaf-1確實可能通過wnt/β-catenin信號通路抑制肝癌細胞。
長期以來由于技術上的瓶頸,基因的研究很長一段時間大部分都停留在克隆和表達層面,功能研究比較缺乏,極大限制了對許多重要基因的功能認知。RNAi修飾技術的出現是對基因功能研究技術的突破,具有特異性強、操作簡便、高效的特點[4]。構建Apaf-1表達載體轉染至肝癌細胞株HepG2中,根據熒光信號確認成功在HepG2中敲降了Apaf-1基因。通過PCR和Wstern blot檢測發現,敲降了Apaf-1基因后,HepG2中β-catenin積聚,引起與相關聯的wnt/β-catenin信號通路的相關基因和因子的表達量改變,例如與細胞有絲分裂相關的CDK2和Cyclin A上升,wnt/β-catenin信號通路上游與β-catenin結合的靶蛋白APC的表達量下降等,從而引起wnt/β-catenin信號通路異常激活,促進了腫瘤的發生,這些結果都進一步揭示了Apaf-1可能參與調控肝癌的發生[13]。
綜上所述,Apaf-1能夠通過wnt/β-catenin信號通路調控肝癌細胞的生成,為進一步在動物體內研究肝癌的發生提供了一定的理論基礎和依據,同時為肝癌的治療提供了新的思路[14]。
[1]ISAYAMA H,NAKAI Y,RERKNIMITR R,et al.The Asian consensus statements on endoscopic management of walled-off necrosis part 1:epidemiology,diagnosis and treatment[J].J Gastroenterol Hepatol,2016,4(4):13394-13402.
[3]CHUNG Y J,KIM H J,PARK S H,et al.Transcriptome analysis reveals that Müllerian inhibiting substance regulates signaling pathways that contribute to endometrial carcinogenesis[J].Int J Oncol,2015,46(5):2039-2046.
[4]LI M,WANG X,JIA T,et al.Tankyrase inhibitors attenuate WNT/β-catenin signaling and inhibit growth of hepatocellular carcinoma cells[J].Oncotarget,2015,6(28):25390-25401.
[5]WU L,YANG P,LUO F,et al.R-spondin1 signaling pathway is required for both the ovarian and testicular development in a teleosts,Nile tilapia(Oreochromis niloticus)[J].Gen Comp Endocrinol,2016,4(1):177-185.
[6]WU L,WU F,LANG X,et al.Synergistic role of β-catenin1 and 2 in ovarian differentiation and maintenance of female pathway in Nile tilapia[J].Mol Cell Endocrinol,2016,15(5):33-44.
[7]GALLINA D,PALAZZO I,STEFFENSON L,et al.Wnt/β-catenin-signaling and the formation of Müller glia-derived progenitors in the chick retina[J].Dev Neurobiol,2016,76(9):983-1002.
[8]XIAO W,JIANG W,SHEN J,et al.Retinoic acid Ameliorates pancreatic fibrosis and inhibits the activation of pancreatic stellate cells in mice with experimental chronic pancreatitis via suppressing the Wnt/β-catenin signaling pathway[J].PLoS One,2015,10(11):e0141462.
[9]OKABE H,KINOSHITA H,IMAI K,et al.Diverse basis of β-catenin activation in human hepatocellular carcinoma:implications in biology and prognosis[J].PLoS One,2016,11(4):e0152695.
[10]YIP L,SOSA J A.Molecular-directed treatment of differentiated thyroid cancer:advances in diagnosis and treatment[J].JAMA Surg,2016,151(7):663-670.
[11]HIRPARA J L,LOH T,NG S B,et al.Aberrant localization of apoptosis protease activating factor-1 in lipid raft sub-domains of diffuse large B cell lymphomas[J].Oncotarget,2016,7(51):83964-83975.
[12]GAO J,LU W F,DAI Z J,et al.Induction of apoptosis by total flavonoids from Scutellaria barbata,D.Don in human hepatocarcinoma MHCC97-H cells via the mitochondrial pathway[J].Tumour Biol,2014,33(3):2549-2559.
[13]PHAM H H,SEONG Y A,OH C W,et al.The herbal medicine cyperus amuricus inhibits proliferation of human hepatocellular carcinoma Hep3B cells by inducing apoptosis and arrest at the G0/G1cell cycle phase[J].Int J Oncol,2016,49(5):2046-2054.
[14]PARK H J,JEON Y K,YOU D H,et al.Daidzein causes cytochrome c-mediated apoptosis via the Bcl-2 family in human hepatic cancer cells[J].Food Chem Toxicol,2013,60(10):542-549.
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
電子制作(2018年11期)2018-08-04 03:25:42
海峽科技與產業(2016年3期)2016-05-17 04:32:12
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
