兩種藥物協同效應對酵母細胞轉錄組的影響
2018-01-17 02:54:56金一然袁辰陽羅靜帆黃新河
生物信息學 2017年4期
沈 成,童 僑,金一然,袁辰陽,羅靜帆,黃新河
(西南交通大學 生命科學與工程學院,成都 610031)
衰老是生物體維持分子、細胞或個體水平生理完整性能力的逐步喪失,表現為細胞或生物體對各種壓力和疾病的抗性逐步下降的生物學過程[1]。衰老幾乎發生于每個物種中,人類也不例外。近年研究顯示,衰老本身是導致衰老相關疾病(包括心血管疾病、II型糖尿病、神經退行性疾病及癌癥等)最主要的風險因子[2],因此,通過延緩衰老本身來預防和治療衰老相關疾病正成為業內共識,而尋找抗衰老活性分子和策略正成為衰老研究領域的重點和熱點。
天然產物多球殼菌素(Myriocin, 分子式C12H39NO6,分子量401.5,一種神經鞘脂合成特異性抑制劑)是前期發現的一種新的抗衰老活性分子[3],雷帕霉素(Rapamycin, 分子式C51H79NO13,分子量914.0,一種TORC1激酶特異抑制劑)是一種廣譜的抗衰老活性分子。前期研究表明,多球殼菌素和雷帕霉素可通過分子間協同效應延緩芽殖酵母細胞衰老并延長細胞壽命[4]。為進一步闡明此協同效應的作用機制,本文考察了該協同效應對細胞轉錄組的影響,進而從生信角度分析了該協同效應調控的細胞過程和信號通路,為闡明該協同效應的作用機制提供依據和思路。
1 材料與方法
1.1 實驗材料與原始數據
實驗菌種、培養基及處理均同文獻[4]。菌種為野生型芽殖酵母(Saccharomycescerevisiae)DBY746;培養基為SDC(Synthetic Dextrose Complete);實驗處理為:無藥物處理組(ND,樣本編號DS-1~5)、45 ng/ml Myr處理組(Myr,樣本編號DS-6~10)、450 pg/ml Rap處理組(Rap,樣本編號DS-11~15)、45 ng/ml Myr及450 pg/ml Rap組合處理組(Syn,樣本編號DS-16~20)。隨后利用RNeasy 試劑盒 (Qiagen, Cat#74104)提取總RNA,在各組中篩選出組內RNA質量濃度相近且質量好的3個樣本,進行基因芯片檢測,得到微陣列MAS5信號值作為原始數據[5-8]。
1.2 生物信息學分析
1.2.1 主成分分析
主成分分析(principal component analysis, PCA)使用R軟件(版本3.3)及R Studio(版本1.0.136),通過擴展包 “psych”,對每個樣本的5 900個探針信號值數據做降維處理,提取3個主成分;通過擴展包“rgl”,將3個主成分的因子載荷(loadings)作為三維坐標,進行可視化。
1.2.2 差異表達基因
通過使用R語言平臺(版本3.3,https://www.r-project.org/ 下載)、“Bioconductor”擴展包分析基因芯片數據,得到差異表達基因(differentially expressed genes, DEGs) 列表(DEGs list)。采用在線程序DAVID (database for annotation, visualization and integrated discovery, 版本6.8, https://david-d.nciFold Changerf.gov/)補充探針注釋[9],YEASTRACT(yeast search for transcriptional regulators and consensus tracking, http://www.yeastract.com/ )補充ORF/Gene Symbol。同時通過擴展包“Vennerable”繪建韋恩圖(Venn diagrams),對差異表達基因進行可視化[10]。
1.2.3 基因本體聚類(GO terms clustering)
采用在線程序DAVID (database for annotation, visualization and integrated discovery, https://david-d.nciFold Changerf.gov/) Functional Annotation Clustering工具[9],對DEGs list (FDR<0.05 & Fold Change>10%)進行基因本體聚類[11],選擇Benjamin=0.05為閾值篩選clusters,選擇Benjamin<0.05 得到Go Terms。
1.2.4 信號通路分析(pathways analysis)
采用在線程序DAVID (database for annotation, visualization and integrated discovery, https://david-d.nciFold Changerf.gov/) Functional Annotation Clustering工具[9],對DEGs list (FDR<0.05 & Fold Change>10%)進行信號通路[12]聚類,選擇Benjamin=0.05為閾值篩選聚類簇,選擇Benjamin<0.05 的信號通路。
2 結果分析
2.1 主成分分析
主成分分析結果見表1。在提取的3個主成分中,協同效應組(Syn)的主成分1因子載荷與其他3組體現出明顯的差異。對表1中數據進行可視化如圖1所示,結果顯示,Syn與單種藥物處理組(Myr, Rap)及單一藥物處理組(Myr, Rap)或無藥物處理組(ND)呈顯著差異,同時,Myr、Rap及ND等3組在主成分上有明顯重疊,這表明低劑量藥物處理芽殖酵母時,與對照組無顯著差異,而兩種藥物組合使用時,產生了顯著差異。
2.2 差異表達基因分析
分析差異表達基因數目可知,相比于無藥物處理組,協同效應用藥導致2 546個基因的表達水平發生顯著變化,而其中超過89%的差異表達基因在Myr組、Rap組未出現。Myr組、Rap組及Syn組處理導致的差異表達基因列表見表2.
通過繪制韋恩圖(如圖2所示)可直觀看出,在該篩選條件下,絕大多數差異表達基因只存在于協同效應組中,協同用藥與單一藥物處理有顯著差異。
2.3 基因本體聚類分析
生物學過程的聚類結果(如圖3所示)顯示,差異表達的基因主要與氧化還原過程(GO:0055114 oxidation-reduction process)、細胞的氧化應激反應(GO:0034599 cellular response to oxidative stress)、三羧酸循環(GO:0006099 tricarboxylic acid cycle)、需氧呼吸(GO:0009060 aerobic respiration)、線粒體電子傳遞(泛醌至細胞色素c)(GO:0006122 mitochondrial electron transport, ubiquinol to cytochrome c)、ATP生物合成途徑(GO:0006754 ATP biosynthetic process)、線粒體電子傳遞(細胞色素c至氧)(GO:0006123 mitochondrial electron transport, cytochrome c to oxygen)、ATP合成偶聯質子轉運(GO:0015986 ATP synthesis coupled proton transport)等過程密切相關。這些生物學過程幾乎均發生在線粒體區域中,推測線粒體相關生物學過程是協同效應的一個重要靶點。
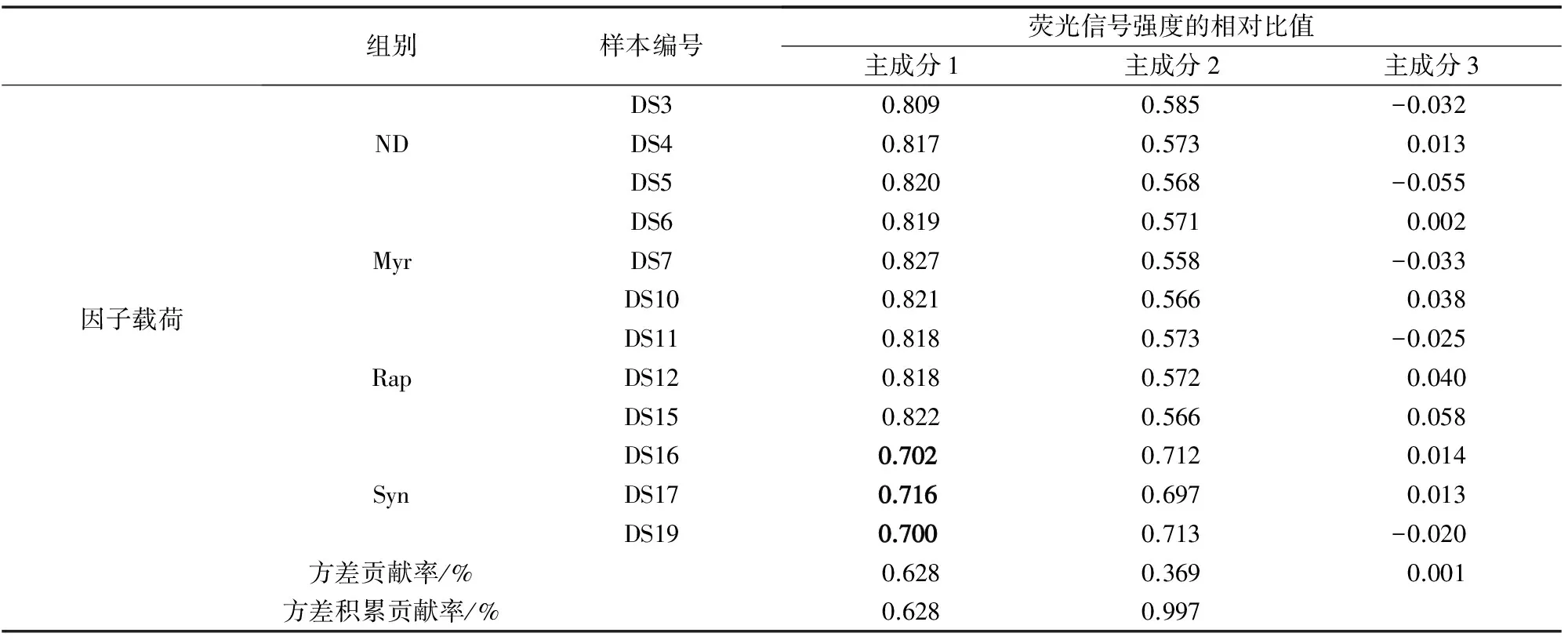
表1 主成分分析數據Table 1 PCA data
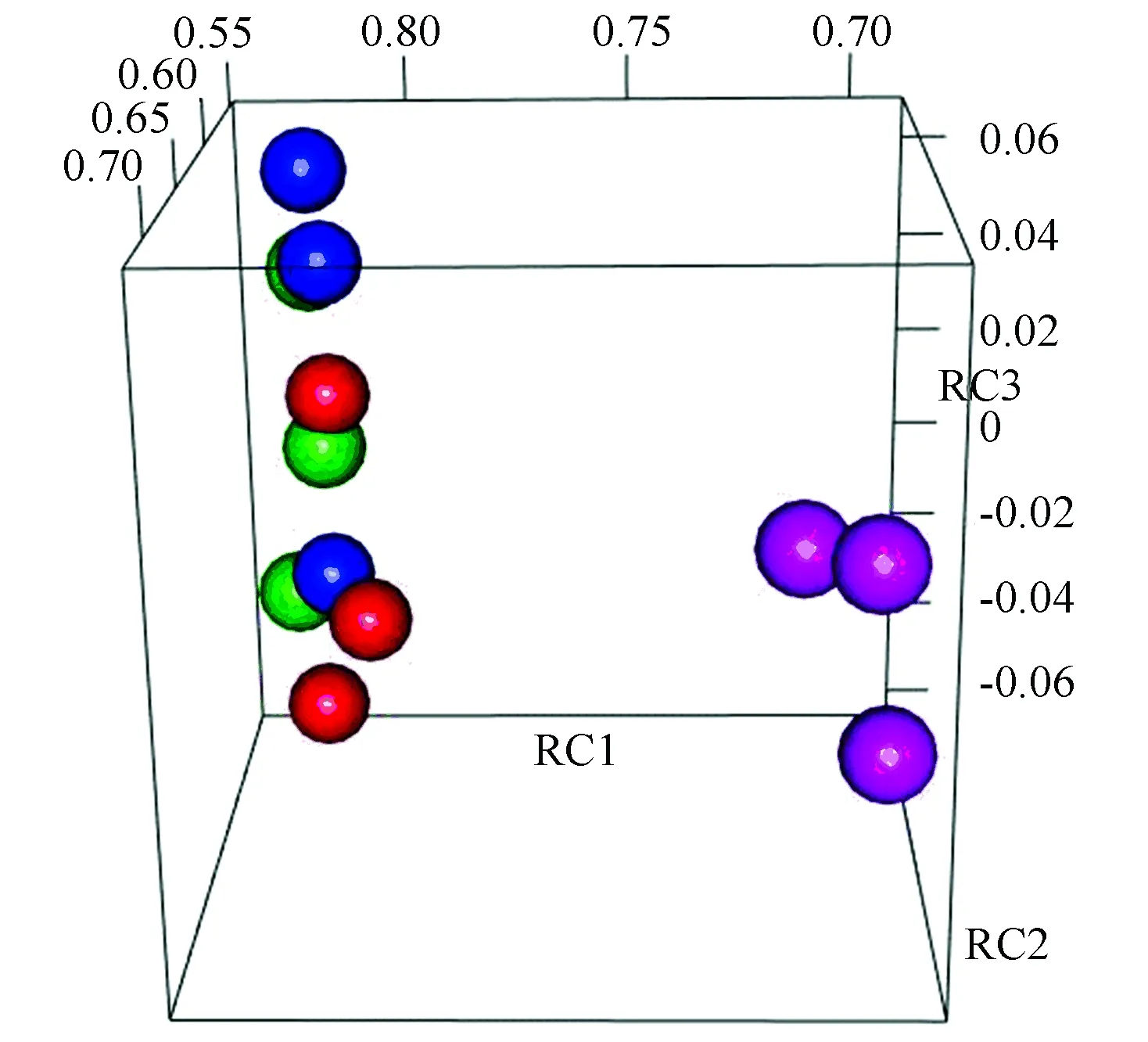
圖1 主成分分析Fig.1 Principle component analysis
注:紅色、藍色、綠色和粉色分別為無藥物處理組、Rap處理組、Myr處理組和協同效應組; RC1、RC2、RC3分別為某樣本的主成分1、2、3的因子載荷。
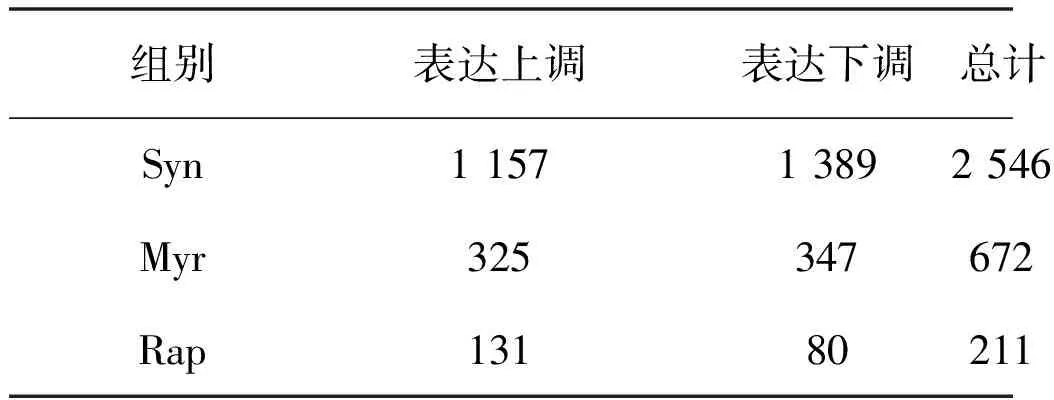
表2 差異表達基因數目Table 2 The number of differentially expressed genes (DEGs)
注:篩選條件為FDR< 0.05,Fold Change > 10%。
細胞組分的聚類分析結果(如圖3所示)顯示,差異表達的基因主要與線粒體(GO:0005739 mitochondrion)、線粒體基質(GO:0005759 mitochondrial matrix)、線粒體內膜(GO:0005743 mitochondrial inner membrane)、呼吸鏈(GO:0070469 respiratory chain)、線粒體膜間隙(GO:0005758 mitochondrial intermembrane space)、線粒體呼吸鏈復合物IV(GO:0005751 mitochondrial respiratory chain complex IV)、線粒體呼吸鏈復合物III(GO:0005750 mitochondrial respiratory chain complex III)、線粒體擬核(GO:0042645 mitochondrial nucleoid)等密切相關。這一結果也顯示,線粒體相關細胞組分是協同效應的重要靶點。
2.4 通路分析
通路分析結果(如圖4所示)顯示,相比于無藥物處理組,協同效應用藥組內147條通路發生改變,其中超過60%的通路在Myr組、Rap組沒有顯著變化。按照Benjamin由小至大排列,在前15條通路中,超過1/3的通路(包括氧化磷酸化(sce00190:Oxidative phosphorylation)、碳代謝(sce01200:Carbon metabolism)、三羧酸循環(sce00020:Citrate cycle)、丙酮酸代謝(sce00620:Pyruvate metabolism)、糖酵解(sce00010:Glycolysis / Gluconeogenesis)等)直接與線粒體功能相關,具體可見附加文件GO term & Pathways.xlsx。
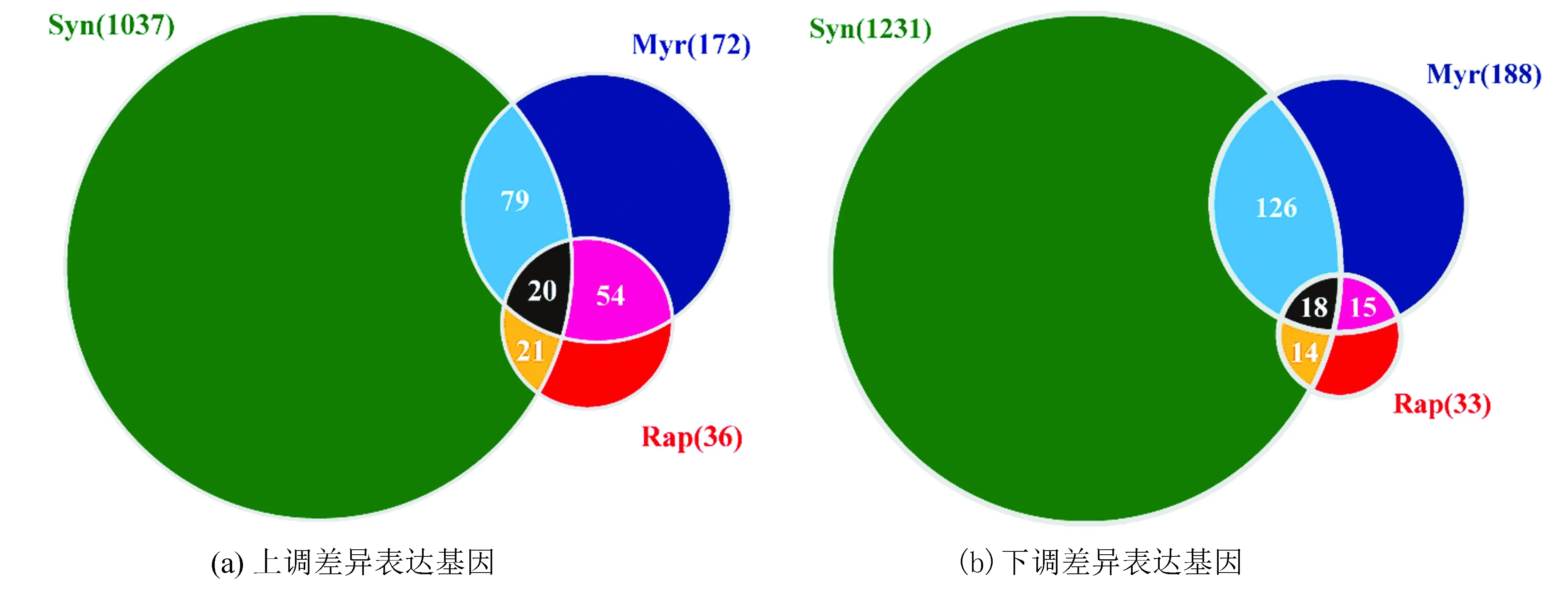
圖2 差異表達基因韋恩圖 Fig.2 Venn diagram of DEGs
注:篩選條件為FDR值< 0.05且Fold change > 10%,圓形區域表示在上述篩選條件下差異表達的基因數量,顏色代表不同的實驗處理組:綠色為Syn組;藍色為Myr組;紅色為Rap組。
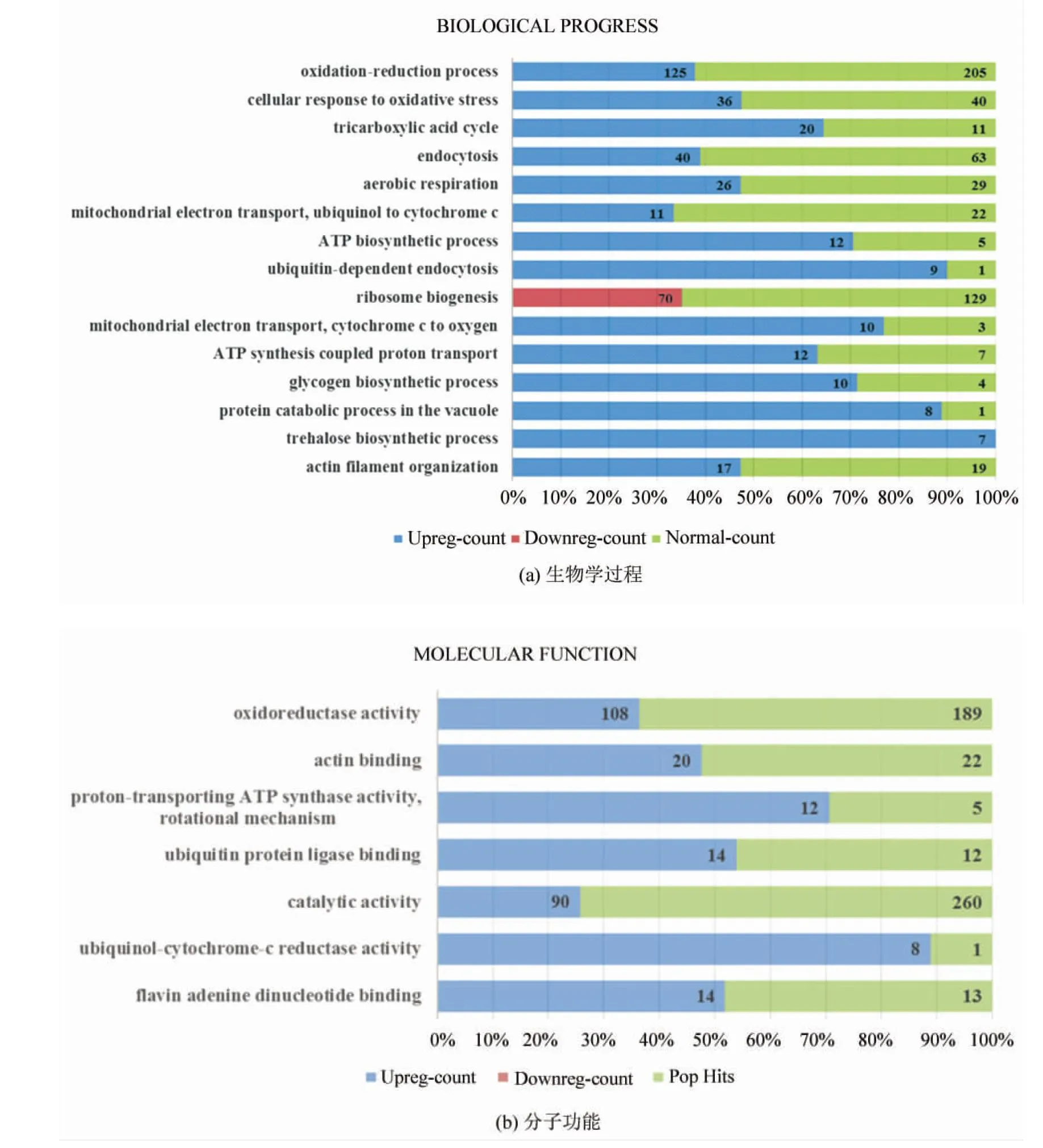
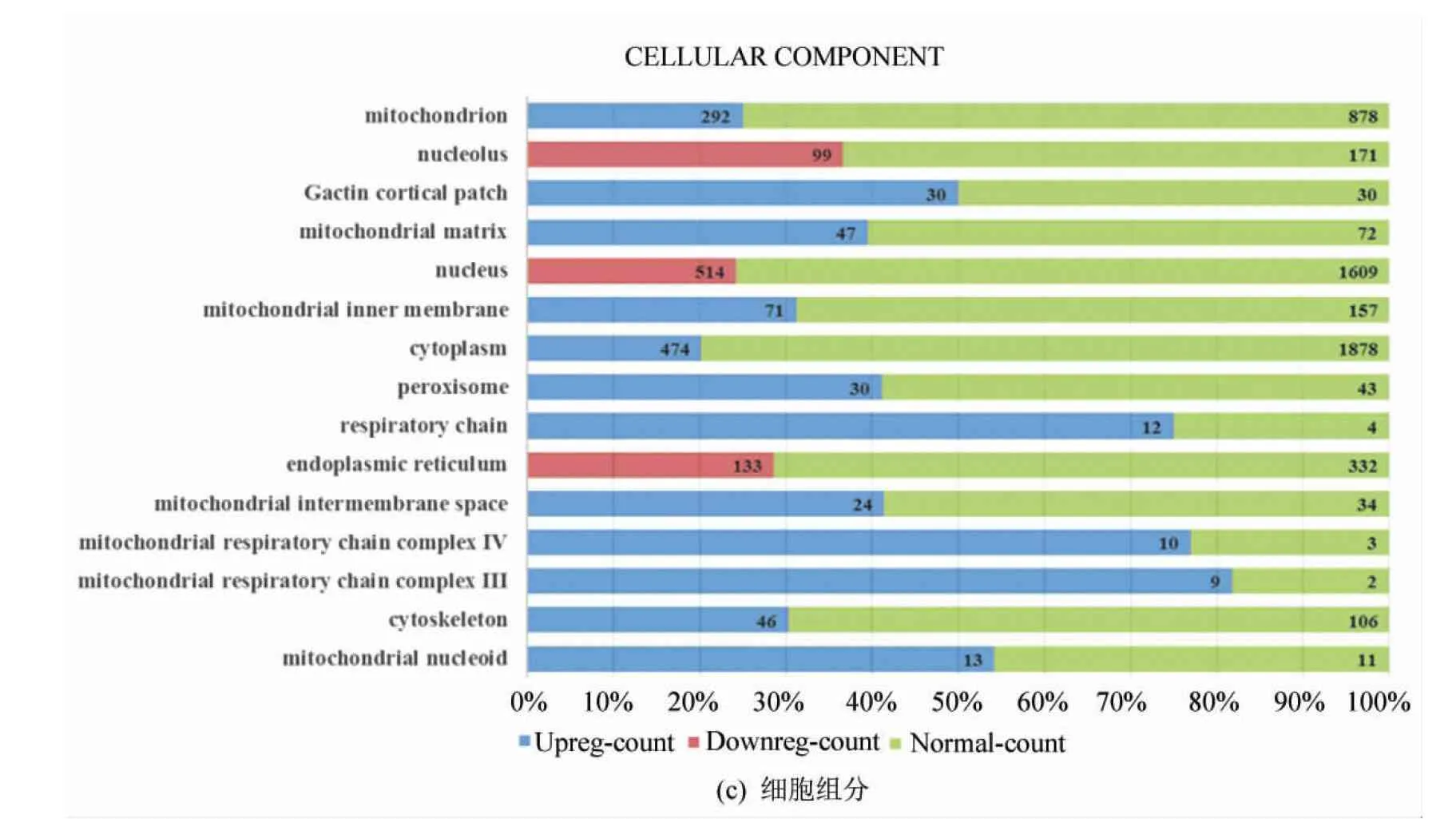
圖3 GO聚類分析
Fig.3 GO terms clustering analysis
注:圖3(a)中左側為不同的基因本體項目(GO terms),右側為某項目中基因表達水平(藍色表示上調、紅色表示下調、綠色表示正常)的計數,圖3(b)、(c)類同。在滿足篩選條件的項目中,對p取負常用對數值,按大小排序,圖3(a)生物學過程取值最大的15個項目,圖3(b)分子功能取全部8項,圖3(c)細胞組分取值最大的15個項目。
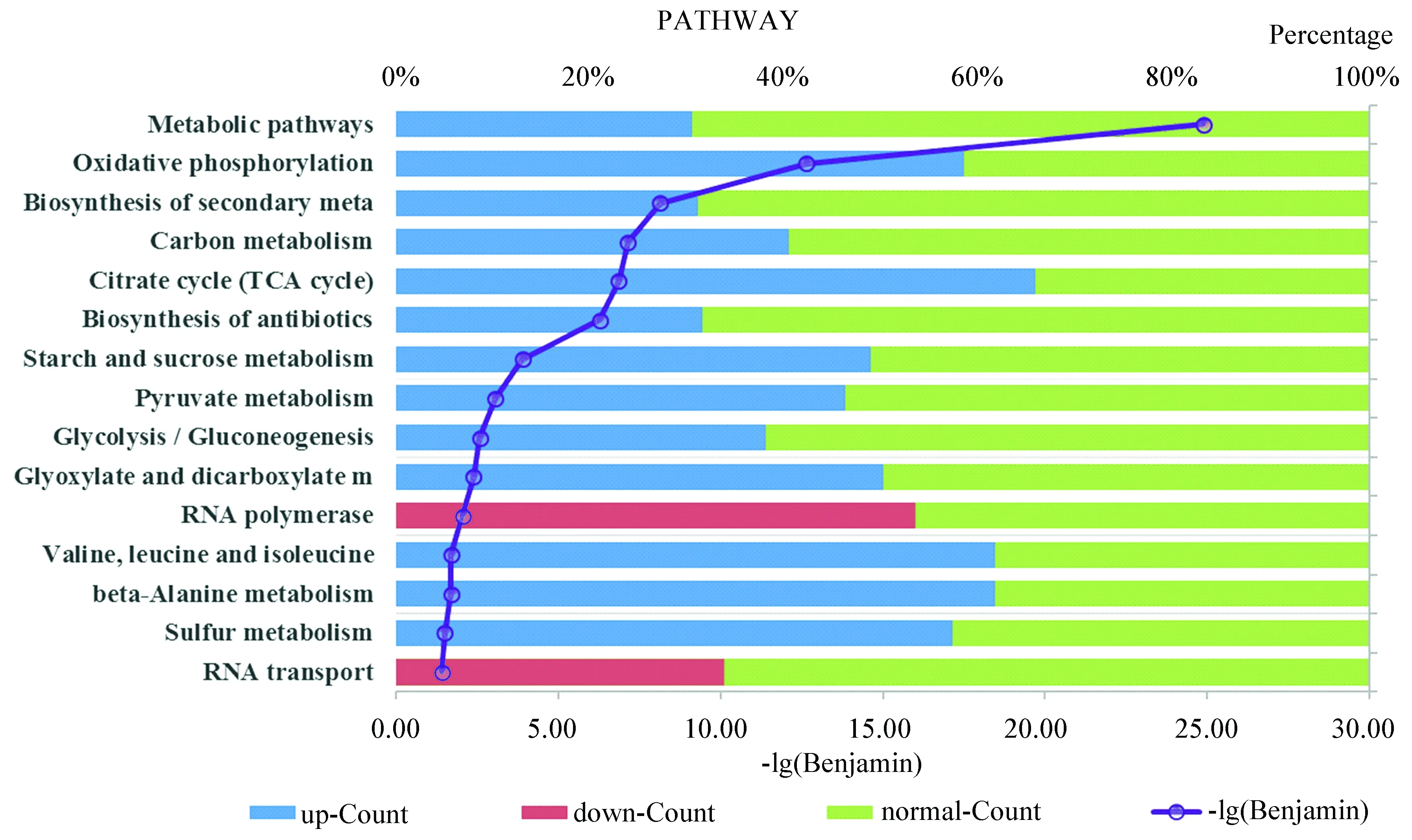
圖4 通路分析Fig.4 Pathway analysis
注:圖中左側為不同的通路,右側為不同表達水平(藍色表示上調、紅色表示下調、綠色表示正常)的差異表達基因在某通路中所占比例。對p取負常用對數值,按大小排序并取值最大的15條通路繪制圖4。
3 討 論
3.1 主成分分析
主成分分析結果顯示單一藥物處理組(包括Myr, Rap)與無藥物處理組在轉錄組水平無顯著差異,說明此低劑量藥物對轉錄組產生極小影響,與實驗中發現的低劑量藥物對細胞壽命影響極小的結論相吻合。
3.2 差異表達基因數量分析
對差異表達基因列表分析顯示,協同效應組表達水平發生顯著變化的2 546個基因中,有超過89%的差異表達基因在Myr組、Rap組未出現。僅在協同藥物處理時表達水平發生顯著變化的基因,簡稱為Unique to Syn。本文認為,導致該情況發生的原因為:1)Unique to Syn的表達受到單一藥物雷帕霉素或多球殼菌素的調控,但由于實驗中選用的藥物濃度極低(45 ng/ml Myr、450 pg/ml Rap),未能使該類基因的表達水平達到本文選取的統計學篩選閾值,即未能使其顯著性差異表達;2)Unique to Syn的表達不受單一藥物雷帕霉素或多球殼菌素的調控,僅僅在兩種小分子同時處理時才會對Unique to Syn起到調控。這兩種猜想還需進一步通過實驗加以判斷。
3.3 差異表達基因聚類分析
差異基因本體聚類及通路分析結果均顯示,線粒體功能,包括線粒體相關的組分(如線粒體內膜、外膜)和生化過程(如氧化磷酸化、三羧酸循環),可能是多球殼菌素和雷帕霉素協同效應調控的重要靶點,后續將從實驗角度加以重點研究。線粒體的氧化反應可能參與衰老和長壽的過程。線粒體是細胞內活性氧的主要來源,線粒體或其DNA受到氧化損傷可能縮短壽命[13]。因此,推測線粒體相關的分子功能、細胞組分、生物學過程和信號通路是此協同效應的一個主要靶點,其與衰老具有密不可分的關系。
4 結 論
1)研究表明,線粒體內蛋白及核酸等氧化損傷是機體衰老的重要原因。Kovalenko等[14]提出線粒體DNA突變在組織細胞衰老過程中起著重要作用,發現線粒體DNA突變引起細胞凋亡才是導致衰老的根本因素,而并非由自由基增多引起的細胞損傷所導致的。
2)近年研究發現,線粒體DNA的突變可以引起某種通路中的信號級聯放大,最終導致細胞程序性死亡[13, 15]。這一研究結果為本文中所考察的協同效應實驗現象的生物學解釋提供了思路,即雷帕霉素和多球殼菌素的組合調控了某種級聯放大通路,導致細胞壽命的顯著延長。
References)
[1]LOPEZ-OTIN C, BLASCO M A, PARTRIDGE L, et al. The hallmarks of aging[J]. Cell, 2013, 153(6): 1194-1217. DOI: 10.1016/j.cell.2013. 05.039.
[2]KENNEDY B K, BERGER S L, BRUNET A, et al. Geroscience: linking aging to chronic disease[J]. Cell, 2014, 159(4): 709-713. DOI: 10.1016/j.cell.2014.10. 039.
[3]HUANG Xinhe, LIU Jun, DICKSON R C. Down-regulating sphingolipid synthesis increases yeast lifespan[J]. PLoS Genet, 2012, 8(2): e1002493. DOI:10.1371/journal.pgen.1002493.
[4]HUANG Xinhe, LIU Jun, WITHERS B R, et al. Reducing signs of aging and increasing lifespan by drug synergy[J]. Aging Cell, 2013, 12(4): 652-660. DOI:10.1111/acel.12090.
[5]IRIZARRY R A, BOLSTAD B M, COLLIN F, et al. Summaries of Affymetrix GeneChip probe level data[J]. Nucleic Acids Res, 2003, 31(4): e15. DOI: 10.1093/nar/gng015.
[6]PEPPER S D, SAUNDERS E K, EDWARDS L E, et al. The utility of MAS5 expression summary and detection call algorithms[J]. BMC Bioinformatics, 2007, 8(273). DOI:10.1186/1471- 2105-8-273.
[7]劉建勝, 張善鎮, 姚志洪,等. 不同表達譜芯片分析方法在藥效機制研究中的應用[J]. 計算機應用與軟件,2015,32(6):57-61,108. DOI:10.3969/j.issn.1000-386x.2015.06.014.
LIU Jiansheng,ZHANG Shanzhen,YAO Zhihong, et al.Application of different microarray analysis methods in pharmacological mechanism research[J].Computer Applications and Software, 2015,32(6):57-61,108. DOI:10.3969/j.issn.1000-386x.2015.06.014.
[8]賈曉東, 陳秀杰, 吳欣, 等. 基于基因表達變異性的通路富集方法研究[J]. 生物化學與生物物理進展, 2013, (12): 1256-1264. DOI:10.3724/SP.J.1206.2012.00410.
JIA Xiaodong, CHEN Xiujie, WU Xin, et al A method of pathway enrichment analysis based gene expression variability[J]. Progress in Biochemistry and Biophysics, 2013,(12):1256-1264.DOI:10.3724/SP.J.1206.2012. 00410.
[9]HUANG D W, SHERMAN B T, LEMPICKI R A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources[J]. Nat Protoc, 2009, 4(1): 44-57. DOI:10.1038/nprot.2008.211.
[10]紀相禹. 基于R語言的差異表達基因檢測研究[D]. 長春:吉林大學, 2011.
JI Xiangyu. The study of differentially expressed gene detection based on r language[D]. Changchun: Jilin University, 2011.
[11]ASHBURNER M, BALL C A, BLAKE J A, et al. Gene ontology: tool for the unification of biology. The gene ontology consortium[J]. Nat Genet, 2000, 25(1): 25-29. DOI:10.1016/j.jcz.2007.05.001.
[12]KANEHISA M, GOTO S. KEGG: kyoto encyclopedia of genes and genomes[J]. Nucleic Acids Research, 2000, 28(1): 27-30.
[13]莫菲, 管德龍, 韓燕,等. 線粒體DNA及相關基因與衰老的關系[J]. 中國老年學雜志, 2016, 36(11): 2796-2799. DOI:10.3969/j.issn.1005- 9202. 2016.11.109.
[14]龍建綱. 衰老及相關疾病中線粒體損傷與保護機制研究[D]. 上海:第二軍醫大學, 2006. DOI:10.7666/d.y897183.
[15]RAIMUNDO N, SONG Lei, SHUTT T E, et al. Mitochondrial stress engages E2F1 apoptotic signaling to cause deafness[J]. Cell, 2012, 148(4): 716-726. DOI:10. 1016/j.cell.2011.12.027.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
