miR-429過表達對膀胱癌細胞遷移、侵襲能力的影響及機制
2018-01-06 01:25:42孫少鵬孔廣起賢少忠蔡大偉
山東醫(yī)藥 2017年45期
孫少鵬,孔廣起,賢少忠,蔡大偉
(首都醫(yī)科大學(xué)附屬北京潞河醫(yī)院,北京通州101149)
miR-429過表達對膀胱癌細胞遷移、侵襲能力的影響及機制
孫少鵬,孔廣起,賢少忠,蔡大偉
(首都醫(yī)科大學(xué)附屬北京潞河醫(yī)院,北京通州101149)
目的探討miR-429過表達對膀胱癌細胞遷移、侵襲能力的影響及機制。方法將常規(guī)培養(yǎng)好的膀胱癌細胞T24隨機分為對照組、miR-429組,對照組轉(zhuǎn)染miR-NC(無關(guān)序列的mimics),miR-429組轉(zhuǎn)染插入miR-429的目標(biāo)片段。用劃痕試驗檢測兩組遷移能力,Transwell侵襲試驗檢測其侵襲能力。采用實時PCR技術(shù)檢測T24細胞內(nèi)E盒結(jié)合鋅指蛋白(ZEB1)、基質(zhì)金屬蛋白酶2(MMP-2)及鈣黏附蛋白E(E-cadherin)mRNA表達,用免疫印跡法檢測T24細胞內(nèi)ZEB1、MMP-2及E-cadherin蛋白表達。結(jié)果與對照組比較,miR-429組細胞遷移距離短,遷移細胞數(shù)少(P均<0.05),細胞內(nèi)ZEB1、MMP-2 mRNA及蛋白表達低,E-cadherin mRNA及蛋白表達高(P均<0.05)。結(jié)論miR-429b過表達可能通過靶向抑制膀胱癌細胞內(nèi)ZEB1、MMP-2表達,增強E-cadherin的表達阻止膀胱癌細胞遷移、侵襲。
膀胱癌;微小RNA-429;細胞遷移;細胞侵襲
膀胱癌是一種惡性腫瘤,占全部惡性腫瘤的1%~2%,其發(fā)病率與病死率均居泌尿系腫瘤的首位,且約50%的患者術(shù)后復(fù)發(fā)[1]。上皮間充質(zhì)轉(zhuǎn)化(EMT)是腫瘤細胞發(fā)生侵襲轉(zhuǎn)移的關(guān)鍵步驟,其表現(xiàn)為上皮鈣黏連蛋白的丟失與上皮鈣黏連蛋白阻遏物表達的增加[2]。在EMT過程中細胞外基質(zhì)發(fā)生溶解,是膀胱癌通過血液和淋巴系統(tǒng)發(fā)生遠處轉(zhuǎn)移的重要作用機制之一[3],其中起主要作用的酶為基質(zhì)金屬蛋白酶2(MMP-2)。微小RNA(miRNA)是一類由18~24個單核苷酸組成的非編碼RNA[4,5]。miR-200家族可靶向調(diào)控多種參與EMT基因的表達[6]。miR-200c可抑制膀胱癌細胞侵襲與增殖[7],miR-200c與miR-205協(xié)同作用抑制膀胱癌的轉(zhuǎn)移[8]。目前有關(guān)miR-429在膀胱癌中的作用尚未見明確報道。2015年9月~2016年12月,本研究對此進行了探討。
1 材料與方法
1.1 細胞、試劑、儀器及其來源 膀胱癌細胞T24由本試驗室提供。DMEM高糖培養(yǎng)基購自美國Sigmag公司;胎牛血清購自以色列Biolnd公司;胰蛋白酶購自長沙研科生物科技有限公司;一抗、二抗購自武漢三鷹科技有限公司;細胞培養(yǎng)板購自美國Costar公司;BCA蛋白濃度檢測試劑盒購自美國Pierce Chemical公司,5×SDS蛋白上樣緩沖液、SDS、Tris、甘氨酸、TEMED、30%Acr-Bis(29∶1)、Glycine、Tween20、脫脂奶粉及過硫酸銨均購自北京鼎國昌盛生物技術(shù)有限責(zé)任公司;D-Hanks緩沖液由本試驗室自行配制。倒置顯微鏡購自日本奧林巴斯公司;CO2培養(yǎng)箱購自美國Forma公司;恒溫-80 ℃冰箱購自中國海爾(Haier)集團;電泳儀、轉(zhuǎn)膜儀購自美國伯樂生物科技公司;板式酶標(biāo)儀購自中國北京新風(fēng)機電公司。
1.2 細胞培養(yǎng)及轉(zhuǎn)染 取含胎牛血清的DMEM培養(yǎng)基,置于37 ℃、5% CO2培養(yǎng)箱中常規(guī)培養(yǎng)T24細胞。用含0.02% EDTA的胰酶消化液消化細胞,待細胞傳至第4、5代,選擇增殖旺盛、生長狀態(tài)良好的細胞進行試驗。將細胞接種入6孔板,生長至50%~60%將其分為對照組、miR-429組,采用脂質(zhì)體轉(zhuǎn)染法,對照組轉(zhuǎn)染miR-NC(無關(guān)序列的mimics),miR-429組轉(zhuǎn)染插入miR-429的目標(biāo)片段5′-TAATACTGTCTGGTAAAACCGT-3′,6 h后換液。轉(zhuǎn)染48 h后收集細胞,用實時PCR檢測mRNA表達,其過表達效率≥90%,證明轉(zhuǎn)染成功。
1.3 T24細胞遷移能力檢測 采用劃痕試驗。轉(zhuǎn)染細胞密度達90%后,用規(guī)格為10 μL的槍頭在6孔板上分別做橫向、縱向各3條劃痕,分別加入細胞增殖抑制劑羥基脲,培養(yǎng)48 h,觀察兩組細胞遷移情況并拍照,測算細胞遷移距離。
1.4 T24細胞侵襲能力檢測 采用Transwell侵襲試驗。于試驗前1 d將基質(zhì)膠放置于4 ℃恒溫冰箱中滆化、過夜。試驗前將所用槍頭在冰上預(yù)冷0.5 h,基質(zhì)膠與預(yù)冷的無血清培養(yǎng)基按1∶7的比例進行稀釋,每孔加入稀釋后的基質(zhì)膠15 μL,于37 ℃培養(yǎng)箱中孵育2~3 h;轉(zhuǎn)染1 d后,收集兩組細胞,用胰酶消化6孔板中的細胞,用濃度為10%血清培養(yǎng)基終止消化,離心,清除含血清的培養(yǎng)基,加入無血清培養(yǎng)基并重新懸浮細胞,細胞稀釋為5×105/mL,每孔加入細胞懸液200 μL;24孔板小室下層加入20% FBS血清800 μL,以供細胞進行侵襲;1~2 d后,顯微鏡下觀察到細胞已侵襲至24孔板底部時立即終止侵襲試驗,棄小室中培養(yǎng)基,PBS漂洗3次,4%多聚甲醛固定20 min,并用0.1%結(jié)晶紫對穿過小室的細胞染色10 min,清水漂洗3次,用棉簽輕輕擦掉上層未侵襲細胞。顯微鏡下觀察并拍照。計算侵襲至小室下層的細胞數(shù)。
1.5 T24細胞內(nèi)E盒結(jié)合鋅指蛋白(ZEB1)、MMP-2及鈣黏附蛋白E(E-cadherin)mRNA表達檢測 采用實時PCR技術(shù)。用TRIzol法提取兩組細胞總RNA,將其逆轉(zhuǎn)錄合成cDNA。反應(yīng)條件:42 ℃、60 min,70 ℃、10 min。將cDNA作為模板,用實時PCR技術(shù),U6為內(nèi)參照,每個樣品設(shè)置3個復(fù)孔。ZEB1引物:P1:5′-gcacaaccaagtgcagaaga-3′,P2:5′-catttgcagattgaggctga-3′;MMP2引物:P1:5′-gctccaccacctacaactttgagaa-3′;P2:5′-tgtgataggatgtgccctggaa-3′;E-cadherin引物:P1:5′-tacactgcccaggagccaga-3′,P2:5′-tggcaccagtccggatta-3′。反應(yīng)條件:95 ℃、30 s,95 ℃、15 s,60 ℃、30 s,共39個循環(huán);然后95 ℃、15 s, 60 ℃、60 s,95 ℃、15 s。通過2%瓊脂糖凝膠電泳及熔解曲線鑒定qRT-PCR產(chǎn)物,并用2-ΔΔCt法進行定量分析。
1.6 T24細胞內(nèi)ZEB1、MMP-2及E-cadherin蛋白表達檢測 采用免疫印跡法。 使用細胞裂解液提取細胞總蛋白,總蛋白經(jīng)BCA法測定濃度后,以30 μg/孔加樣,經(jīng)10% SDS-PAGE凝膠電泳分離,將轉(zhuǎn)印了蛋白的PVDF膜用5%脫脂牛奶封閉,并培育一抗及相應(yīng)的HRP標(biāo)記二抗,ECL化學(xué)發(fā)光法顯影。使用Quantityone軟件測定條帶灰度值,蛋白的相對含量=目的蛋白條帶灰度值/內(nèi)參照條帶灰度值。
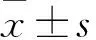
2 結(jié)果
2.1 miR-429過表達對T24細胞遷移、侵襲能力的影響 培養(yǎng)48 h,對照組、miR-429組遷移距離分別為(10.09±0.4)、(5.2±0.23)mm。miR-429組細胞遷移距離短于對照組(P<0.05)。對照組、miR-429組遷移細胞數(shù)分別為(102±3)、(36±8)個。miR-429組遷移細胞數(shù)少于對照組(P<0.05)。
2.2 miR-429過表達對T24細胞ZEB1、MMP-2、E-cadherin表達的影響 見表1。
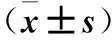
表1 兩組ZEB1、MMP-2、E-cadherin表達比較
注:與對照組比較,*P<0.05。
3 討論
膀胱癌為泌尿系常見的惡性腫瘤之一,年新增病例38.63萬,病死病例15.02萬,發(fā)病率在惡性腫瘤中居第9位,致死率列全球男性泌尿生殖系統(tǒng)腫瘤第2位。75%的患者為非肌層浸潤膀胱癌,5年生存率較高;25%為肌層浸潤膀胱癌,在首次積極治療后容易發(fā)生轉(zhuǎn)移。探尋基于分子網(wǎng)絡(luò)的新型治療模式、早期診斷及預(yù)后標(biāo)志物成為近年研究的熱點。EMT是指具有極性結(jié)構(gòu)的上皮細胞喪失緊密連接結(jié)構(gòu)的過程。在該過程中,上皮細胞轉(zhuǎn)變?yōu)殚g質(zhì)細胞,獲得了間質(zhì)細胞的分子標(biāo)記物,且具有更強的侵襲和遷移能力。EMT與腫瘤轉(zhuǎn)移具有相同的特點,即調(diào)節(jié)因素多、機制復(fù)雜。
miRNA通過與其靶mRNA分子3′非編碼區(qū)特異堿基部分完全或不完全配對結(jié)合,從而導(dǎo)致其發(fā)生降解或翻譯抑制,于轉(zhuǎn)錄后水平調(diào)控基因表達。研究證實,miRNA可調(diào)控>60%的人類蛋白編碼基因,并在多種病理生理進程中發(fā)揮關(guān)鍵作用。miRNA既可作為抑癌基因又可作為原癌基因參與腫瘤的發(fā)生發(fā)展。鑒于其疾病/組織特異性表達圖譜及驚人的調(diào)控潛能,miRNA已成為腫瘤診斷及預(yù)后的標(biāo)志物。miR-429是miR-200家族中具有抑癌作用的成員之一。本研究證實,miR-429過表達對T24細胞遷移能力起到了一定的抑制作用,并同時抑制細胞侵襲,其主要原因可能為miR-429過表達抑制腫瘤細胞EMT的發(fā)生,延緩其進程。miR-429能夠抑制ZEB1的表達同時恢復(fù)E-cadherin的表達,從而阻滯膀胱癌發(fā)生侵襲轉(zhuǎn)移。目前研究認為,miR-429在多種腫瘤組織中表達下調(diào),如膀胱癌[9]、胃癌[10]、腎癌[11]、結(jié)直腸癌[12]和鼻咽癌[13],在其他腫瘤組織中miR-429的表達也發(fā)生上調(diào)。miR-200家族可直接靶向ZEB1和ZEB2,進而抑制EMT的發(fā)生。過表達miR-200c能夠升高E-cadherin的表達[14]。本研究也發(fā)現(xiàn),過表達miR-429能夠抑制ZEB1的表達,恢復(fù)E-cadherin的表達。在miR-429所介導(dǎo)的EMT信號通路中E-cadherin是十分重要的指標(biāo),ZEB1與ZEB2在膀胱癌組織中的表達異常升高[15],并且ZEB2的高表達與膀胱癌患者的不良預(yù)后存在相關(guān)性,同時E-cadherin的丟失也會導(dǎo)致患者預(yù)后不良。
綜上所述,miR-429能夠通過靶向抑制ZEB1的表達實現(xiàn)對EMT的抑制,從而遏制腫瘤細胞的擴散和轉(zhuǎn)移。本研究為今后膀胱癌的臨床治療提供了新方向,而miR-429很可能作為膀胱癌臨床治療的新靶點。
[1] Kausch I, Bohle A. Molecular aspects of bladder cancer Ⅲ. Prognostic markers of bladder cancer[J]. Eur Urol, 2002,41(1):15-29.
[2] McConkey DJ, Choi W, Marquis L, et al. Role of epithelial-to-mesenchymaltransition (EMT) in drug sensitivity and metastasis in bladder cancer[J]. Cancer Metastasis Rev, 2009,28(3):335-344.
[3] Szarvas T, vom Dorp F, Ergun S, et al. Matrix metalloproteinases and their clinical relevance in urinary bladder cancer[J]. Nat Rev Urol, 2011,8(5):241-254.
[4] Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function[J]. Cell, 2004,116(2):281-297.
[5] Zaravinos A. The regulatory role of microRNAs in EMT and cancer[J]. J Oncol, 2015,2015(25):865816.
[6] Adam L, Zhong M, Choi W, et al. miR-200 expression regulates epithelial-tomesenchymal transition in bladder cancer cells and reverses resistance to epidermal growth factor receptor therapy[J]. Clin Cancer Res, 2009,15(16):5060-5072.
[7] Liu L, Qiu M, Tan G, et al. miR-200c inhibits invasion, migration and proliferation of bladder cancer cells through downregulation of BMI-1 and E2F3[J]. J Transl Med, 2014,12(1):305.
[8] Wiklund ED, Bramsen JB, Hulf T, et al. Coordinated epigenetic repression of the miR-200 family and miR-205 in invasive bladder cancer[J]. Int J Cancer, 2011,128(6):1327-1334.
[9] Chen Z, Liu X, Hu Z, et al. Identification and characterization of tumor suppressor and oncogenic miRNAs in gastric cancer[J]. Oncol Lett, 2015,10(1):329-336.
[10] Karlsson R, Pedersen ED, Wang Z, et al. Rho GTPase function in tumorigenesis[J]. Biochim Biophys Acta, 2009,1796(2):91-98.
[11] Qiu M, Liang Z, Chen L, et al. MicroRNA-429 suppresses cell proliferation, epithelial-mesenchymal transition, and metastasis by direct targeting of BMI1 and E2F3 in renal cell carcinoma[J]. Urol Oncol, 2015,33(7):332.e9-e18.
[12] Sun Y, Shen S, Tang H, et al. miR-429 identified by dynamic transcriptome analysis is a new candidate biomarker for colorectal cancer prognosis[J]. OMICS, 2014,18(1):54-64.
[13] Thiery JP, Acloque H, Huang RY, et al. Epithelialmesenchymal transitions in development and disease[J]. Cell, 2009,139(5):871-890
[14] Hurteau GJ, Carlson JA, Spivack SD, et al. Overexpression of the microRNA hsa-miR-200c leads to reduced expression of transcription factor 8 and increased expression of E-cadherin[J]. Cancer Res, 2007,67(17):7972-7976.
[15] Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition[J]. Nat Rev Mol Cell Biol, 2014,15(3):178-196.
10.3969/j.issn.1002-266X.2017.45.013
R737.14
A
1002-266X(2017)45-0042-03
2017-10-11)
猜你喜歡
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫(yī)科大學(xué)學(xué)報(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
