長鏈非編碼RNA MYU對宮頸癌Hela細(xì)胞增殖的影響及其機(jī)制
2018-01-06 01:25:42周白云郭永利王卓王文琴
山東醫(yī)藥 2017年45期
關(guān)鍵詞:檢測
周白云,郭永利,王卓,王文琴
(黔西南州人民醫(yī)院,貴州黔西南州562400)
長鏈非編碼RNA MYU對宮頸癌Hela細(xì)胞增殖的影響及其機(jī)制
周白云,郭永利,王卓,王文琴
(黔西南州人民醫(yī)院,貴州黔西南州562400)
目的探討長鏈非編碼RNA MYU對宮頸癌Hela細(xì)胞增殖的影響及其機(jī)制。方法檢測長鏈非編碼RNA MYU在宮頸癌組織和癌旁組織中的表達(dá)差異,將Hela細(xì)胞株分為:1組:轉(zhuǎn)染對照NC序列;2組:轉(zhuǎn)染MYU siRNA;3組:轉(zhuǎn)染對照pcDNA3.1;4組:轉(zhuǎn)染pcDNA3.1-MYU過表達(dá)序列。采用CCK-8法檢測各組細(xì)胞細(xì)胞增殖活力,Western blotting和RT-PCR檢測c-myc、CDK4、CDK6基因mRNA及蛋白的表達(dá),采用RNA結(jié)合蛋白免疫沉淀試驗驗證WDR5與MYU的結(jié)合,染色質(zhì)免疫沉淀試驗檢測組蛋白H3賴氨酸4三甲基化(H3K4me3)水平。結(jié)果癌組織LncRNA MYU的表達(dá)量高于癌旁組織(P<0.05);轉(zhuǎn)染24、48 h,OD值比較,2組低于1組,4組高于3組,P均<0.05;Ki-67陽性細(xì)胞所占百分比比較,1組高于2組,3組低于4組,P均<0.05;c-myc 、CDK4、CDK6 mRNA及蛋白比較,1組高于2組,3組低于4組,P均<0.05;LncRNA MYU與WDR5的富集量高于陰性對照(P<0.05);H3K4me3水平比較,1組低于2組,3組高于4組,P均<0.05。結(jié)論長鏈非編碼RNA MYU通過募集WDR5抑制c-myc基因啟動子區(qū)H3K4me3,上調(diào)c-myc轉(zhuǎn)錄從而促進(jìn)Hela細(xì)胞增殖。
宮頸癌;Hela細(xì)胞;長鏈非編碼RNA MYU;c-myc;WDR5;組蛋白H3賴氨酸4三甲基化
宮頸癌的發(fā)生發(fā)展具有多種學(xué)說及機(jī)制,但究其根本原因,癌基因的異常過度轉(zhuǎn)錄及抑癌基因的表達(dá)抑制發(fā)揮著基礎(chǔ)作用,因此尋找新的宮頸癌致病基因,使其作為治療靶點,尤為必要[1]。非編碼RNA的產(chǎn)物是成熟的RNA轉(zhuǎn)錄本,一般認(rèn)為不翻譯成蛋白質(zhì),分為microRNA、piRNA、tRNA、長鏈非編碼RNA (LncRNA)等幾大類[2~4]。宮頸癌相關(guān)的microRNA研究較為透徹,包括microRNA-99b、microRNA-545等[4]。LncRNA是一類長度超過200 bp的轉(zhuǎn)錄本,同樣具有廣泛的生物學(xué)功能[5,6]及多種調(diào)控反式[7],然而當(dāng)前關(guān)于宮頸癌相關(guān)的LncRNA卻知之甚少。既往研究發(fā)現(xiàn),LncRNA MYU通過與hnRNP-K結(jié)合,穩(wěn)定細(xì)胞周期調(diào)控激酶CDK6轉(zhuǎn)錄,從而促進(jìn)結(jié)腸癌進(jìn)展[8]。我們的前期研究發(fā)現(xiàn)MYU在宮頸癌組織中高表達(dá),本研究中我們探討了LncRNA MYU對宮頸癌細(xì)胞增殖的影響及其機(jī)制。現(xiàn)報告如下。
1 材料與方法
1.1 組織、細(xì)胞株、試劑及儀器 宮頸癌組織及對應(yīng)癌旁組織20例份均來自于本院收集的樣本,起止時間為2012年2月~2016年5月。Hela細(xì)胞株購自上海中科院生化細(xì)胞所細(xì)胞庫,培養(yǎng)基為DMEM basic培養(yǎng)基+10%胎牛血清+雙抗。siRNA由廣州銳博生物技術(shù)有限公司合成,pcDNA3.1-MYU過表達(dá)質(zhì)粒由上海生工生物技術(shù)有限公司構(gòu)建,啟動子采用巨細(xì)胞病毒,Lipofectamine 2000TM(Invitrogen公司),熒光染料SYBR Green (TAKATA公司),c-myc抗體、CDK4抗體、CDK6抗體、Ki-67抗體、WDR5抗體、組蛋白H3賴氨酸4三甲基化(H3K4me3)抗體(abcam公司),引物(上海生工生物公司),protein G(Santa Cruz公司),細(xì)胞裂解及蛋白提取液(碧云天公司),CCK8試劑盒(日本同仁公司),RNA提取試劑盒、逆轉(zhuǎn)錄試劑盒(TAKARA公司),RNA結(jié)合蛋白免疫沉淀(RIP)試劑盒(Millipore公司),染色質(zhì)免疫沉淀(CHIP)試劑盒(Millipore公司)。熒光定量PCR檢測儀(Bio-Rad公司,CFX96),Western blotting電泳及轉(zhuǎn)膜儀(Bio-Rad公司)。
1.2 組織及細(xì)胞LncRNA MYU、c-myc、CDK4、CDK6 mRNA表達(dá)的檢測 使用TRIzol法提取組織及細(xì)胞的RNA,反轉(zhuǎn)錄反應(yīng)體系20 μL,反應(yīng)條件為:37 ℃、30 min,85 ℃、5 s,4 ℃、10 min。采用熒光定量PCR檢測基因表達(dá)情況,反應(yīng)條件為92 ℃、3 min預(yù)變性,92 ℃、30 s,58 ℃、10 s(39個循環(huán)),72 ℃、2 min, 4 ℃、10 min。所有操作嚴(yán)格按照使用說明書進(jìn)行,采用2-ΔΔCT表示目的基因的相對表達(dá)量。引物序列:MYU:F:5′CTTCAGGCGTGTTTTCCCTG3′;R:5′CGACACCTCCAAGATGCTGA3′;c-myc:F:5′CCCTCCACTCGGAAGGACTA3′;R:5′GCTGGTGCATTTTCGGTTGT3′;CDK4:F:5′CAGCTGGTCACATGGTGAGG3′;R:5′TTCCTACGGCCCCATACACC3′;CDK6:F:5′TGGTAACTCCTTCCCCAGAGT3′;R:5′CGAAGCGAAGTCCTCAACAC3′;GAPDH:F:5′AATGGGCAGCCGTTAGGAAA3′;R:5′GCGCCCAATACGACCAAATC3′。
1.3 細(xì)胞中c-myc、CDK4、CDK6蛋白表達(dá)的檢測 采用Western blotting法。轉(zhuǎn)染結(jié)束后,使用RIPA弱型裂解液裂解細(xì)胞,冰上孵育30 min后,12 000 r/min轉(zhuǎn)速離心30 min,吸取上層蛋白,測量濃度,加入蛋白loading buffer高溫變性。隨后按試劑盒步驟操作。一抗工作濃度為1∶500,相應(yīng)熒光二抗工作濃度為1∶20 000。Odessay軟件對Western blotting膜進(jìn)行曝光掃描后,采用Quantity One軟件分析各蛋白的灰度值,并校正于內(nèi)參GAPDH,以作為各組相應(yīng)蛋白表達(dá)量。
1.4 細(xì)胞分組及轉(zhuǎn)染 將Hela細(xì)胞株分為:1組:轉(zhuǎn)染對照NC序列;2組:轉(zhuǎn)染MYU siRNA;3組:轉(zhuǎn)染對照pcDNA3.1;4組:轉(zhuǎn)染pcDNA3.1-MYU過表達(dá)序列。siRNA及過表達(dá)質(zhì)粒的轉(zhuǎn)染均采用Lipofectamine 2000TM包裹,細(xì)胞密度約70%時進(jìn)行轉(zhuǎn)染,每組分別轉(zhuǎn)染對應(yīng)序列,轉(zhuǎn)染前2 h,將細(xì)胞培養(yǎng)基換為無血清培養(yǎng)基,細(xì)胞轉(zhuǎn)染8 h換為10%血清的培養(yǎng)基,轉(zhuǎn)染48 h后檢測轉(zhuǎn)染效率及細(xì)胞表型變化。
1.5 細(xì)胞增殖活力檢測 采用CCK-8法及Ki-67染色法。CCK-8法:細(xì)胞種植于96孔板中,干擾/過表達(dá)48 h后,加入CCK-8試劑,置于37 ℃細(xì)胞孵箱孵育1 h,熒光酶標(biāo)儀檢測450 nm吸光度(OD)值。Ki-67染色法:細(xì)胞轉(zhuǎn)染后48 h后,以4%多聚甲醛固定,Triton-100打孔,隨后按照1∶100的濃度稀釋Ki-67抗體,4 ℃孵育過夜后進(jìn)行熒光二抗染色。高倍鏡下隨機(jī)選擇5個視野,計數(shù)Ki-67染色陽性細(xì)胞及總細(xì)胞數(shù)量,計算其在總細(xì)胞數(shù)量中所占百分比。
1.6 WDR5與MYU結(jié)合的鑒定 采用RIP實驗。嚴(yán)格按照Millipore公司試劑盒說明書操作步驟操作。實驗中分組:WDR5組,IgG組,分別孵育WDR5抗體及IgG。細(xì)胞裂解后,加入檢測抗體,抗體的工作濃度為每反應(yīng)體系8 μg,4 ℃搖床孵育過夜后,室溫復(fù)溫1 h,加入protein G磁珠捕獲復(fù)合體,洗滌buffer洗滌之后,提取RNA,逆轉(zhuǎn)錄PCR之后,熒光定量PCR檢測MYU富集于WDR5蛋白上的RNA水平。
1.7 c-myc基因啟動子H3K4me3水平的檢測 采用CHIP實驗。嚴(yán)格按照Millipore公司試劑盒說明書操作步驟操作。細(xì)胞超聲破碎、交聯(lián)之后,加入H3K4me3抗體孵育,4 ℃搖床孵育過夜后,再次磁珠捕獲、提取DNA,PCR檢測c-myc基因啟動子區(qū)H3K4me3水平。
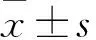
2 結(jié)果
2.1 LncRNA MYU在宮頸癌患者癌組織及癌旁組織的表達(dá)比較 癌組織LncRNA MYU的表達(dá)量為2.35±0.23,癌旁組織為1.12±0.19,兩者比較,P<0.05。
2.2 敲除或過表達(dá)LncRNA MYU對Hela細(xì)胞增殖的影響 轉(zhuǎn)染24、48 h,OD值比較,2組低于1組,4組高于3組,P均<0.05。見表1。1、2、3、4組Ki-67陽性細(xì)胞所占百分比分別為52.23%±5.77%、24.39%±4.58%、42.11%±5.24%、72.94%±6.83%,Ki-67陽性細(xì)胞所占百分比比較,1組高于2組,3組低于4組,P均<0.05。
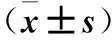
表1 轉(zhuǎn)染0、24、48 h各組細(xì)胞OD值比較
2.3 敲除或過表達(dá)LncRNA MYU對Hela細(xì)胞c-myc、CDK4、CDK6基因mRNA及蛋白表達(dá)的影響 c-myc、CDK4、CDK6 mRNA及蛋白表達(dá)比較,1組高于2組,3組低于4組,P均<0.05。見表2。
表2 轉(zhuǎn)染各組細(xì)胞c-myc、CDK4、CDK6基因mRNA及蛋白表達(dá)比較
2.4 LncRNA MYU與WDR5在Hela細(xì)胞中的結(jié)合情況 WDR5組LncRNA MYU與WDR5的富集量為0.032 0±0.003 1,IgG組為0.001 2±0.000 1,兩組比較,P<0.05。
2.5 敲除或過表達(dá)LncRNA MYU對Hela細(xì)胞c-myc基因啟動子H3K4me3水平變化的影響 1、2、3、4組c-myc基因啟動子H3K4me3水平分別為0.033±0.006、0.078±0.004、0.164±0.011、0.082±0.007,H3K27me3水平比較,1組低于2組,3組高于4組,P均<0.05。
3 討論
宮頸癌是最常見婦科惡性腫瘤,HPV感染是其主要的危險因素。既往研究表明宮頸癌高發(fā)年齡為30~35歲,浸潤癌為45~55歲,但是近年隨著環(huán)境、飲食等因素改變,其發(fā)病開始出現(xiàn)年輕化趨勢。
疾病的發(fā)生是外在環(huán)境和內(nèi)在基因異常轉(zhuǎn)錄共同作用的結(jié)果,但是基因的異常轉(zhuǎn)錄扮演著始動角色。過去,研究者認(rèn)為編碼基因才起關(guān)鍵作用,因為編碼基因的產(chǎn)物是蛋白質(zhì),非編碼基因的產(chǎn)物只是核酸,而蛋白質(zhì)是最終效應(yīng)器,因此,占基因座上絕大部分位置的非編碼基因被認(rèn)為是一片荒漠,其轉(zhuǎn)錄產(chǎn)物被稱之為“轉(zhuǎn)錄垃圾”。非編碼RNA分為microRNA、piRNA、tRNA、LncRNA等幾大類別,其中microRNA的相關(guān)研究目前開展的非常廣泛,其主要發(fā)揮抑制作用。既往研究證實,眾多microRNA分子在宮頸癌演進(jìn)中扮演關(guān)鍵角色,例如microRNA-140-5p通過靶向IGF2BP1抑制宮頸癌增殖和轉(zhuǎn)移,miR-143/145與宮頸癌預(yù)后相關(guān)。
LncRNA是一類長度超過200 bp的轉(zhuǎn)錄本,作為新興的非編碼物質(zhì),被認(rèn)為具有相較microRNA更為復(fù)雜的作用方式,例如HotAir、Xist等可以與蛋白修飾復(fù)合體互相作用,影響組蛋白H3K27表觀遺傳學(xué)改變,而H19則可以作為分子篩,發(fā)揮類似“海綿作用”吸附microRNA分子,最終影響靶mRNA。LncRNA既可以促進(jìn)基因的轉(zhuǎn)錄,也可以發(fā)揮抑制作用;既可以結(jié)合核酸,也可以結(jié)合蛋白;既可以修飾蛋白質(zhì),也可以影響mRNA的成熟和穩(wěn)定。
本研究中,我們發(fā)現(xiàn)LncRNA MYU在宮頸癌組織中高表達(dá),提示可能存在生物學(xué)效能。Kawasaki等[8]發(fā)現(xiàn),LncRNA MYU通過與RNA結(jié)合蛋白hnRNP-K結(jié)合,穩(wěn)定細(xì)胞周期調(diào)控激酶CDK6轉(zhuǎn)錄,從而發(fā)揮促進(jìn)細(xì)胞增殖的作用。Ki-67是細(xì)胞增殖相關(guān)的核抗原,可對整個細(xì)胞有絲分裂周期的細(xì)胞進(jìn)行標(biāo)記,其陽性率越高,表明細(xì)胞增殖越活躍。本研究發(fā)現(xiàn)敲除LncRNA MYU之后, Hela細(xì)胞的增殖活力及Ki-67陽性細(xì)胞數(shù)降低,而在病毒載體過表達(dá)LncRNA MYU之后,Hela細(xì)胞的增殖活力及Ki-67陽性細(xì)胞數(shù)升高。進(jìn)一步研究證實,敲除LncRNA MYU將下調(diào)c-myc、CDK4、CDK6的轉(zhuǎn)錄水平、翻譯水平表達(dá),而過表達(dá)則上調(diào)其表達(dá)。基因的異常轉(zhuǎn)錄是由染色體的開閉狀態(tài)決定的,開放狀態(tài)的染色體結(jié)構(gòu)利于轉(zhuǎn)錄因子與基因啟動子的結(jié)合,啟動轉(zhuǎn)錄,而關(guān)閉狀態(tài)的染色體則限制了轉(zhuǎn)錄因子的結(jié)合,轉(zhuǎn)錄則受到抑制。組蛋白H3的不同賴氨酸殘基的甲基化狀態(tài)對基因的轉(zhuǎn)錄激活或抑制發(fā)揮不同的作用,H3K4的三甲基化(H3K4me3)、H3k9的三甲基化(H3K9me3)提示基因的轉(zhuǎn)錄激活狀態(tài),而H3K27的三甲基化(H3K27me3)則提示轉(zhuǎn)錄抑制。通過CHIP實驗,我們發(fā)現(xiàn)LncRNA MYU可以作為表觀遺傳學(xué)的調(diào)控元件,影響H3K4me3,從而影響c-myc基因的轉(zhuǎn)錄,上調(diào)CDK4、CDK6,從而促進(jìn)Hela細(xì)胞增殖。RIP實驗提示,LncRNA MYU與TrxG/MLL復(fù)合體中的WDR5可直接結(jié)合,WDR5是組蛋白H3的重要調(diào)節(jié)因子,通過輔助TrxG/MLL復(fù)合體抑制甲基化H3K4,從而發(fā)揮轉(zhuǎn)錄激活作用。
綜上所述,LncRNA MYU在宮頸癌組織中高表達(dá),通過與WDR5結(jié)合,募集TrxG/MLL復(fù)合體至組蛋白,改變組蛋白H3的表觀遺傳學(xué)狀態(tài),抑制H3K4me3發(fā)生,從而上調(diào)c-myc基因的轉(zhuǎn)錄,進(jìn)一步上調(diào)CDK4、CDK6,發(fā)揮促Hela細(xì)胞增殖的作用。
[1] Hammer A, Rositch A, Qeadan F,et al. Age-specific prevalence of HPV16/18 genotypes in cervical cancer: a systematic review and meta-analysis[J]. Int J Cancer, 2016,138(12):2795-2803.
[2] Chen LL. Linking long noncoding RNA localization and function[J]. Trends Biochem Sci, 2016,41(9):761-772.
[3] Stuwe E, Tóth KF, Aravin AA. Small but sturdy: small RNAs in cellular memory and epigenetics[J]. Genes Dev, 2014,28(5):423-431.
[4] Duval M, Cossart P, Lebreton A. Mammalian microRNAs and long noncoding RNAs in the host-bacterial pathogen crosstalk[J]. Semin Cell Dev Biol, 2016,65(16):11-19.
[5] Li L, Liu B,Wapinski OL,et al. Targeted disruption of Hotair leads to homeotic transformation and gene derepression[J]. Cell Rep, 2013,5(1):3-12.
[6] Sarma K, Cifuentes-Rojas C, Ergun A, et al. ATRX directs binding of PRC2 to Xist RNA and Polycomb targets [J]. Cell, 2014,159(4):869-883.
[7] Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways[J]. Cancer Cell, 2016,29(4):452-463.
[8] Kawasaki Y, Komiya M, Matsumura K, et al. MYU, a target lncRNA for Wnt/c-Myc signaling, mediates induction of CDK6 to promote cell cycle progression[J]. Cell Rep, 2016,16(10):2554-2564.
10.3969/j.issn.1002-266X.2017.45.012
R737.33
A
1002-266X(2017)45-0039-03
2017-08-09)
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
