TIAM1在兒童神經母細胞瘤中的研究進展
2017-12-21 01:08:25邱艷麗綜述趙強審校
中國腫瘤臨床 2017年22期
邱艷麗 綜述 趙強 審校
TIAM1在兒童神經母細胞瘤中的研究進展
邱艷麗 綜述 趙強 審校
隨著分子診斷技術的不斷發展,精準治療已進入惡性腫瘤的治療范疇,靶向治療是近年來較為熱門的研究方向。神經母細胞瘤(neuroblastoma,NB)是兒童期最常見的顱外實體瘤。研究表明,很多基因參與NB的發生發展過程,T淋巴瘤侵襲轉移誘導因子1(T lymphoma invasion and metastasis inducing factor 1,TIAM1)是其中之一。TIAM1主要與下游的RAC1(Ras-related C3 botulinum toxin substrate 1)結合,作用于TrkA/TIAM1/RAC1通路,激活下游相關因子,參與調節神經元軸突的分化過程。因此,深入的研究和實驗或許可以更透徹的闡明具體的機制,為NB未來的診療提供一個新的方向。
神經母細胞瘤 兒童 TIAM1 分化 研究進展
神經母細胞瘤(neuroblastoma,NB)是兒童最常見的顱外惡性實體瘤,起源于腎上腺髓質或椎旁交感神經系統。在美國每年大約確診700~800例[1],中國大約每百萬人群中有兒童患兒7.7例,占兒童實體腫瘤的9.8%[2]。其生物學行為多樣,病因復雜,腫瘤異質性明顯,治療較為困難,被稱為“兒童腫瘤之王”。目前在多模式高強度的治療下,由于常規治療效果不佳,不良反應較為明顯,幾乎無空間去增加現有的治療強度,分層診斷和靶向治療或許是打破目前治療的瓶頸。近年來,對兒童惡性腫瘤相關基因的研究越來越多,T淋巴瘤侵襲轉移誘導因子1(T lymphoma invasion and metastasis inducing factor 1,TIAM1)是其中之一。研究發現,TIAM1在正常組織中表達較低,但具有多種生物學活性,在多種腫瘤細胞系中表達,如胃癌[3]、肝細胞癌[4]、前列腺癌[5]、胰腺癌[6]、導管性乳腺癌[7]、喉鱗狀細胞癌[8]、肺腺癌[9]、卵巢癌[10]等。目前關于TIAM1在NB中的研究越來越多,但研究結果尚未達成統一共識。本文就近年來關于TIAM1在NB形成過程中的影響進行綜述,旨在為兒童NB的診療提供一個新的方向。
1 TIAM1的生物學特點
TIAM1最初在小鼠T淋巴瘤細胞高侵襲變異株中分離鑒定而來。小鼠TIAM1位于第16號染色體的遠側端[11],人TIAM1位于第21號染色體q22上。TIAM1有1 591個氨基酸,蛋白分子量大小為177 KDa[12]。在成人的大腦、睪丸及表皮中相對高表達,在小鼠胚胎發育中沉默TIAM1基因的表達,發現小鼠胚系大腦發育嚴重缺陷,表明TIAM1蛋白在胚胎發生中起關鍵作用[13]。
1.1 TIAM1的結構域及表達
TIAM1具有細胞質蛋白的特性,有酪氨酸、絲氨酸及蘇氨酸等多個潛在的磷酸化位點,主要在細胞質中表達,有較大程度的親水性。TIAM1是一個多結構域的蛋白質,在氨基末端區域,存在一個豆蔻酰化位點(Myr結構域),此位點可能參與蛋白質的定位和穩定性;其下游是兩個相鄰的PEST結構域,可能參與調節蛋白質的穩定性。C1199-TIAM1,即N端豆蔻酰化位點Myr及兩個PEST結構域缺失,已有研究顯示這種突變體較TIAM1全長片段更穩定,更具有活性[14]。C1199-TIAM1片段包括一個PHn-CC-Ex單位,其對質膜的相互聯系及下游支架蛋白(Eph,Spinophilin,JIP2/IB2,Par3,CD44,Ankyrin)的信號轉導起決定性作用。PHn-CC-Ex下游的結構域包括與Ras蛋白結合的RBD結構域及與DHR同源的PDZ結構域。在羧基端區域,有一個集GEFs家族所有成員特點所在的DH-PH(PHc)結構域,不同于僅一個PH結構域的Dbl家族,TIAM1存在兩個PH結構域,一個為C末端的PHc結構域,另一個為N末端的PHn結構域[15]。TIAM1蛋白中的DH-PH可介導蛋白質復合物的形成及TIAM1的膜定位,參與蛋白質間的相互作用。其中有研究表明PHn對膜皺褶起主要作用[16];另有研究發現,PHc和PHn結構域共同參與細胞偽足的形成。這兩個PH結構域可能互相協同、互相補充,共同發揮不可替代的生物學作用(表1)[17]。
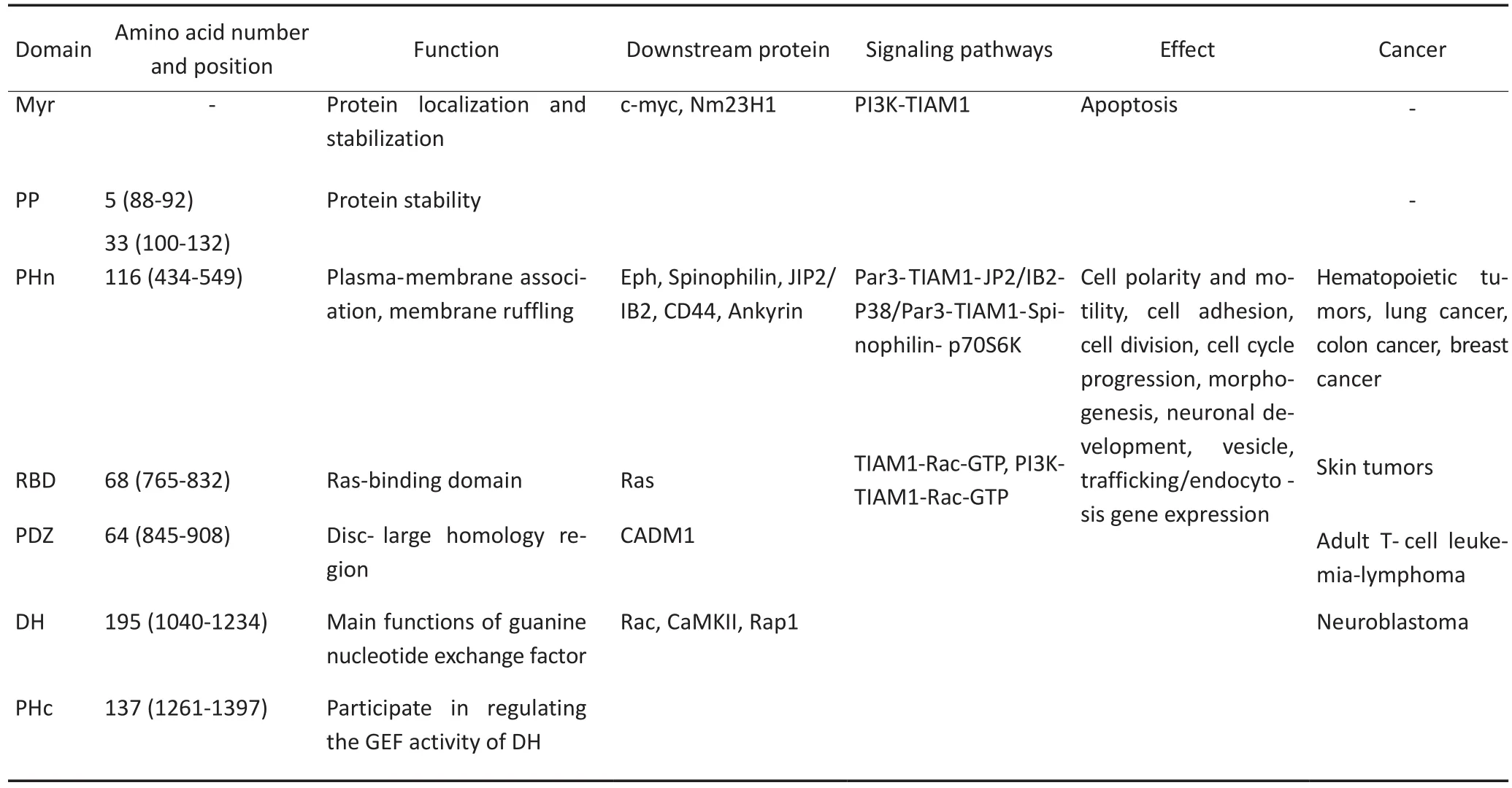
表1 TIAM1主要結構域的基本功能及信號通路Table 1 The basic functions and signaling pathways of the main domain of the TIAM1
1.2 TIAM1的生物學功能
TIAM1參與一系列分子事件,其中最重要的方面在于其可作為小G蛋白Rho家族重要成員RAC1的特異性鳥嘌呤核苷酸交換因子,激活RAC1,調節RAC1參與的信號通路,RAC1又活化一系列的下游底物,促進腫瘤侵襲轉移。TIAM1活化后促進GDP釋放使GTP結合到Rho家族成員中,Rho樣GTPase可以促使整合素聚集至細胞表面,將信號傳遞至肌動蛋白細胞骨架,影響細胞骨架的組裝、運動及細胞形體極化過程,從而影響細胞的形態、遷移、黏附、生長、存活、極性以及內吞作用和膜轉運。因此,該基因在細胞侵襲、轉移和癌變過程中起到重要作用[18-19]。有研究顯示,活化的TIAM1可以誘導肌動蛋白微絲在質膜上聚集,產生片狀和絲狀偽足,也可以誘導膜皺褶,調節E-鈣黏素介導的細胞黏附,和Rho一起參與黏附復合物的組裝,從而誘導腫瘤細胞侵襲轉移。也有研究得出在神經生長過程中,該基因參與了神經元的遷移、軸突的生長和神經元的極化[20-21]。過表達的TIAM1誘導NB細胞的擴散和影響神經軸突生長,在原始的海馬趾和中樞神經中,沉默TIAM1表達后導致樹突,棘突數量的減少和密度的下降[22-24],但具體作用機制尚不明確。
2 TIAM1在NB中的變異情況及對NB預后的影響
隨著TIAM1在腫瘤細胞中的異常表達被逐步證實,TIAM1在NB中的作用也逐步被揭示。Sanmartin等[25]對106例初治的NB患兒行二代測序,發現TI-AM1突變率占11%,而且在MYC、RAS及RAC涉及的3條信號通路中,TIAM1突變的患兒預后較好,同時發現3個MYCN與TIAM1一同突變的高危NB患兒均未復發,其中2例已經治愈。對TIAM1突變的這部分患兒進一步分析發現,TIAM1基因突變通過影響該基因中3個主要結構域,進而影響其參與的信號通路,使蛋白質的功能發生障礙,表達量并無明顯變化。Pugh等[26]研究發現,在240例高危NB患兒中未發現任何TIAM1突變,可能篩查的重點是致瘤性的體細胞突變,忽略了TIAM1保護性的胚系變異所致。相反,Nishimura等[27]對87例NB患兒行全基因組測序結果發現3例TIAM1突變,其中1例伴隨MYCN擴增,預后較差。Cao等[28]對33例兒童NB患兒行二代測序時發現7例TIAM1胚系突變及1例TIAM1缺失。上述研究結果均表明TIAM1在NB形成過程中可能影響其發生和進展,但其具體的作用機制尚不明確。
2.1 TIAM1參與NB形成的結構域
據報道,TIAM1主要通過以下3個結構域影響NB:1)N端區域,結合MYC家族成員促進轉錄激活并且抑制細胞凋亡;2)RAS結合域(RBD)轉導信號的下游效應;3)催化結構域(DH-PH2)激活RAC1,從而激活細胞遷移和軸突形成。TIAM1的代謝轉換由泛素-蛋白酶體途徑調節:對泛素化敏感的兩個酪氨酸殘基(K1404 and K1420)與C端DH-PHc結構域結合后立即起作用[29]。K1469定位接近兩泛素化的賴氨酸,在1例患兒中發現K1469R變異,并且這種變異可能改變TIAM1的定位或代謝轉換,還有可能減小其與MYCN轉錄激活的能力,因為這種變異可以與其他轉錄激活因子相互作用,如NCOA3和雌激素受體[30]。另外,突變的K1469R可以產生新的剪接位點,這種改變可能導致mRNA的減少及蛋白質穩定性的下降。蛋白酶體抑制劑硼替佐米與維甲酸已被建議作為選擇性聯合輔助治療NB防止復發[31]。然而,這例患兒及伴隨TIAM1突變R788Q的患兒均未復發,進一步表明TIAM1在NB預后結果中是一種保護性的變異。
2.2 TIAM1參與小鼠NB神經元細胞極性的調節
神經元細胞是高度極化的細胞之一,極化的形式主要有兩種即軸突和樹突。PAR-3、PAR-6和非典型的蛋白激酶C在多種細胞極化事件中發揮作用,研究發現TIAM1/TIAM2與PAR-3-aPKC-PAR-6-Cdc42-GTP形成復雜的復合物在小鼠NB中介導板狀偽足的形成[27]。在小鼠NB細胞系NIE-115中,過表達的TIAM1可以誘導NB細胞的擴散及軸突的形成,并且層黏連蛋白明顯增加了上述作用。過表達的TIAM1可以招募層黏連蛋白受體a6b1整聯蛋白遷移到腫瘤細胞周邊特異的黏著部位,促進異質型黏附;也可將a6b1募集于運動細胞前部和偽足處,傳遞調節信號至細胞骨架,增強腫瘤細胞的運動和侵襲,上述作用主要依賴于PI3K-TIAM1-RAC1信號通路[32]。另有研究發現,在海馬神經元中活躍的TIAM1可以誘導多軸突樣突起的分化,促進神經元突起的延伸[33],并且在此過程中,細胞骨架與細胞外基質的相互作用對軸突的形成具有一定的影響。TIAMl可以定位在成神經細胞瘤細胞的生長端胞膜的內側,參與生長端形體重塑及調解神經突的形成。當成神經元細胞與基質層黏連蛋白黏附后,可以通過TIAM1-RACl信號途徑調節瘤細胞形體的延展與神經突形成,促進其運動和侵襲[33]。
2.3 TIAM1可能參與調節神經生長因子與軸突間的信號通路
許多研究支持TIAM1在細胞膜皺裂,神經細胞信息傳遞,軸突的形成中發揮一定的作用[22,34]。近期有研究顯示,RAC1-GEF TIAM1復雜的信號通路在神經營養因子NGF、NT-3和BDNF介導的肌動蛋白細胞骨架重塑中起到關鍵的作用,不同的神經營養因子對應的受體不同,作用也各不相同。NGF/TrkA及NT-3/TrkC分別作用于神經突的向外延伸和施萬細胞的遷移,而BDNF/TrkB則參與皮層神經細胞的神經突生長[35-36]。雖然激活神經生長因子Trk的受體TrkA可以觸發細胞內涉及RAC1的信號通路,但是關于神經生長因子受體TrkB和TrkC對RAC1特異性因子TIAM1的不同激活機制也已有報道。盡管TIAM1調節神經生長因子NT-3和BDNF介導的RAC1活性,但是通過此途徑激活的RAC1是否對神經生長因子NGF及其受體TrkA有生物學反應還尚不明確。在神經生長因子NGF的作用下,分化中的交感神經元及神經元細胞系PC12明顯表達TIAM1,推測TIAM1可能參與神經生長因子受體TrkA的下游信號通路及神經突的延伸[20],并且神經元軸突的缺失可能促使NB的形成[37]。
對于胚胎性NB來說,其發生主要是因為胚胎發育異常,即分化障礙致神經干細胞不能正常分化為成熟的神經元及神經膠質細胞[38]。既往研究表明,TrkA/TIAM1/RAC1調控神經細胞的極性,軸突的延伸以及膠質的形成,有促進正向分化的作用,當TIAM1缺失后導致神經細胞去分化的發生。而與此相反的另一信號通路P75NGFR/RhoA,起到負調控TrkA/TIAM1/RAC1的作用,將向外突出的軸突退縮,細胞變圓,起到了去分化的作用。因此,推斷上述兩種信號通路共同參與調控NB的分化(圖1)[14]。
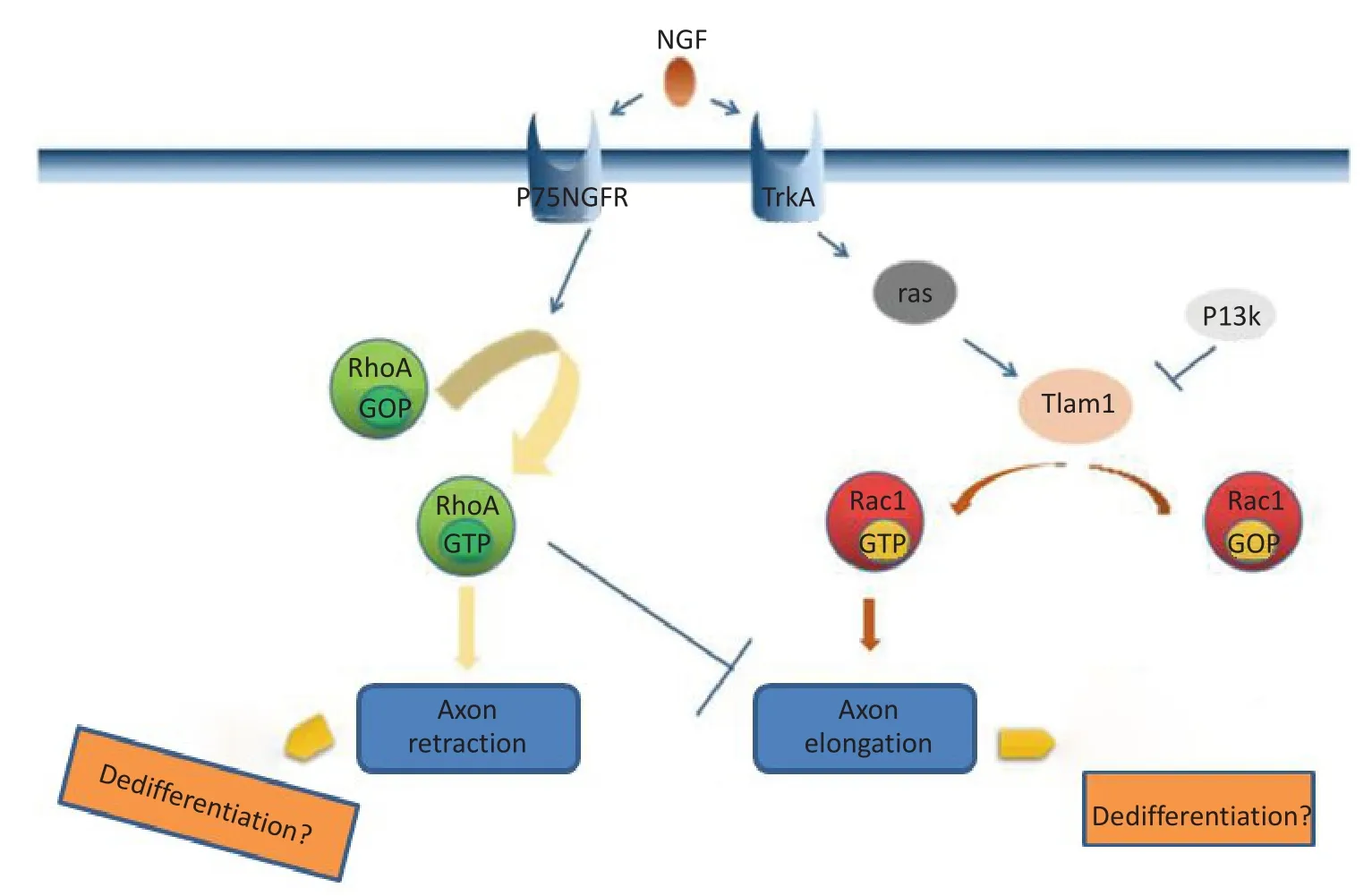
?圖1 TrkA/TIAM1/RAC1和P75NGFR/RhoA信號通路示意圖Figure 1 Schematic of TrkA/TIAM1/RAC1 and P75NGFR/RhoA signaling pathways
3 總結與展望
20年前有研究發現了TIAM1基因,其在細胞遷移、黏附、浸潤、凋亡、腫瘤形成過程中發揮重要的作用。本文重點綜述了TIAM1在NB形成中的作用。部分TIAM1突變的NB預后較好,但也有過表達的TIAM1可以促進NB細胞的轉移與浸潤,誘導NB的發生與進展。目前為止,在NB形成過程中TIAM1明確的分子機制尚不明確,亟需大量的研究證明TIAM1在NB發生發展中的具體作用,旨在為NB的相關治療診斷提供幫助,同時為NB的化療、放療及預后提供更多的依據。
[1]Coughlan D,Gianferante M,Lynch CF,et al.Treatment and survival of childhood neuroblastoma:evidence from a population-based study in the United States[J].Pediatr Hematol Oncol,2017:1-11.
[2]He J,Zou Y,Wang T,et al.Genetic variations of GWAS-identified genes and neuroblastoma susceptibility:a replication study in southern Chinese children[J].Transl Oncol,2017,10(6):936-941.
[3]Li Z,Yu X,Wang Y,et al.By downregulating TIAM1 expression,microRNA-329 suppresses gastric cancer invasion and growth[J].Oncotarget,2015,6(19):17559-17569.
[4]Chen G,Lu L,Liu C,et al.MicroRNA-377 suppresses cell proliferation and invasion by inhibiting TIAM1 expression in hepatocellular carcinoma[J].PLoS One,2015,10(3):e0117714.
[5]Engers R,Mueller M,Walter A,et al.Prognostic relevance of Tiam1 protein expression in prostate carcinomas[J].Br J Cancer,2006,95(8):1081-1086.
[6]Guo X,Wang M,Zhao Y,et al.Par3 regulates invasion of pancreatic cancer cells via interaction with Tiam1[J].Clin Exp Med,2016,16(3):357-365.
[7]Li Z,Liu Q,Piao J,et al.Clinicopathological implications of Tiam1 overexpression in invasive ductal carcinoma of the breast[J].BMC Cancer,2016,16(1):681.
[8]Wang S,Li S,Tang Q,et al.Overexpression of Tiam1 promotes the progression of laryngeal squamous cell carcinoma[J].Oncol Rep,2015,33(4):1807-1814.
[9]Liu S,Li Y,Qi W,et al.Expression of Tiam1 predicts lymph node metastasis and poor survival of lung adenocarcinoma patients[J].Diagn Pathol,2014,(9):69.
[10]Li H,Cui X,Chen D,et al.Clinical implication of Tiam1 overexpression in the prognosis of patients with serous ovarian carcinoma[J].Oncol Lett,2016,12(5):3492-3498.
[11]Habets GG,Ra VDK,Jenkins NA,et al.The invasion-inducing TIAM1 gene maps to human chromosome band 21q22 and mouse chromosome 16[J].Cytogene Geno Res,2008,70(1-2):48-51.
[12]Xu Z,Gakhar L,Bain FE,et al.The Tiam1 guanine nucleotide exchange factor is auto-inhibited by its pleckstrin homology coiledcoil extension domain[J].2017,292(43):17777-17793.
[13]Yoo S,Kim Y,Lee H,et al.A gene trap knockout of the Tiam-1 protein results in malformation of the early embryonic brain[J].Mole Cells,2012,34(1):103-108.
[14]Boissier P,Huynhdo U.The guanine nucleotide exchange factor Tiam1:a janus-faced molecule in cellular signaling[J].Cellular Signalling,2014,26(3):483-491.
[15]Xu Z,Gakhar L,Bain FE,et al.The Tiam1 guanine nucleotide exchange factor is auto-inhibited by its pleckstrin homology coiledcoil extension domain[J].2017.[Epub ahead of print].
[16]Stam JC,Sander EE,Michiels F,et al.Targeting of Tiam1 to the plasma membrane requires the cooperative function of the N-terminal pleckstrin homology domain and an adjacent protein interaction domain[J].J Biol Chem,1997,272(45):28447-28454.
[17]Crompton AM,Foley LH,Wood A,et al.Regulation of Tiam1 nucleotide exchange activity by pleckstrin domain binding ligands[J].J Biol Chem,2000,275(33):25751-25759.
[18]Worthylake DK,Rossman KL,Sondek J.Crystal structure of Rac1 in complex with the guanine nucleotide exchange region of Tiam1[J].Nature,2000,408(6813):682-688.
[19]Shepherd TR,Klaus SM,Liu X,et al.The Tiam1 PDZ domain couples to Syndecan1 and promotes cell-matrix adhesion[J].J Mol Biol,2010,398(5):730-746.
[20]Fard SS,Kele J,Vilar M,et al.Tiam1 as a signaling mediator of nerve growth factor-dependent neurite outgrowth[J].PLoS One,2010,5(3):e9647.
[21]Kawauchi T,Chihama K,Nabeshima Y,et al.The in vivo roles of STEF/Tiam1,Rac1 and JNK in cortical neuronal migration[J].Embo J,2003,22(16):4190-4201.
[22]Zhang H,Macara IG.The polarity protein PAR-3 and TIAM1 cooperate in dendritic spine morphogenesis[J].Nat Cell Biol,2006,8(3):227-237.
[23]Tolias KF,Bikoff JB,Burette A,et al.The Rac1-GEF Tiam1 couples the NMDA receptor to the activity-dependent development of dendritic arbors and spines[J].Neuron,2005,45(4):525-538.
[24]Tolias KF,Bikoff JB,Kane CG,et al.The Rac1 guanine nucleotide exchange factor Tiam1 mediates EphB receptor-dependent dendritic spine development[J].Proc Natl Acad Sci USA,2007,104(17):7265-7270.
[25]Sanmartin E,Yanez Y,Fornes-Ferrer V,et al.TIAM1 variants improve clinical outcome in neuroblastoma[J].Oncotarget,2017,8(28):45286-45297.
[26]Pugh TJ,Morozova O,Attiyeh EF,et al.The genetic landscape of high-risk neuroblastoma[J].Nat Genet,2013,45(3):279-284.
[27]Nishimura T,Yamaguchi T,Kato K,et al.PAR-6-PAR-3 mediates Cdc42-induced Rac activation through the Rac GEFs STEF/Tiam1[J].Nat Cell Bio,2005,7(3):270.
[28]Cao Y,Jin Y,Yu J,et al.Clinical evaluation of integrated panel testing by nextgeneration sequencing for somatic mutations in neuroblastomas withMYCNunamplification[J].Oncotarget,2017.[Epubaheadofprint].
[29]Genau HM,Huber J,Baschieri F,et al.CUL3-KBTBD6/KBTBD7Ubiquitin ligase cooperates with GABARAP proteins to spatially restrict TIAM1-RAC1 signaling[J].Molecular Cell,2015,57(6):995-1010.
[30]Ferrero M,Avivar A,Garcíamacías MC,et al.Phosphoinositide 3-kinase/AKT signaling can promote AIB1 stability independently of GSK3 phosphorylation[J].Cancer Res,2008,68(13):5450.
[31]Barbara H,Yania Y,Sarai P,et al.Targeting neuroblastoma stem cells with retinoic acid and proteasome inhibitor[J].PLoS One,2013,8(10):e76761-e76761.
[32]Keely PJ,Westwick JK,Whitehead IP,et al.Cdc42 and Rac1 induce integrin-mediated cell motility and invasiveness through PI(3)K[J].Nature,1997,390(6660):632.
[33]Matsuo N,Terao M,Nabeshima Y,et al.Roles of STEF/Tiam1,guanine nucleotide exchange factors for Rac1,in regulation of growth cone morphology[J].Mol Cell Neurosci,2003,24(1):69-81.
[34]Leeuwen FN,Kain HE,Kammen RA,et al.The guanine nucleotide exchange factor Tiam1 affects neuronal morphology;opposing roles for the small GTPases Rac and Rho[J].J Cell Biol,1997,139(3):797-807.
[35]Yamauchi J,Miyamoto Y,Tanoue A,et al.Ras activation of a Rac1 exchange factor,Tiam1,mediates neurotrophin-3-induced Schwann cell migration[J].Neurosci Res,2005,102(41):14889-14894.
[36]Miyamoto Y,Yamauchi J,Tanoue A,et al.TrkB binds and tyrosinephosphorylates Tiam1,leading to activation of Rac1 and induction of changes in cellular morphology[J].Proce Nat Acade Sci USA,2006,103(27):10444-10449.
[37]Molenaar JJ,Koster J,Zwijnenburg DA,et al.Sequencing of neuroblastoma identifies chromothripsis and defects in neuritogenesis genes[J].Nat Int Week J Sci,2012,483(7391):589-593.
[38]Speleman F,Park JR,Henderson TO.Neuroblastoma:a tough nut to crack[J].Am Soci Clin Oncol Edu Book Am Soci Clin Oncol Meet,2016,(35):e548.
Research progress on TIAM1 in pediatric neuroblastoma
Yanli QIU,Qiang ZHAO
Department of Pediatric Oncology,Tianjin Medical University Cancer Institute and Hospital;National Clinical Research Center for Cancer;Key Laboratory of Cancer Prevention and Therapy,Tianjin;Tianjin's Clinical Research Center for Cancer,Tianjin 300060,China
Qiang ZHAO;E-mail:qiangzhao169@sina.com
With continuous development of molecular diagnosis,"precision treatment"has entered the therapeutic category for malignant tumors,and targeted gene therapy has been an intense research topic in recent years.Neuroblastoma is the most common extracranial solid tumor that develops during childhood.Studies show that many genes,such as TIAM1,are involved in the development and progression of neuroblastoma.TIAM1 mainly combines with RAC1 to activate downstream factors that mediate differentiation via the TrkA/TIAM1/RAC1 signaling pathway,which is involved in the regulation of neurite.Therefore,further studies and experiments may reveal the specific mechanisms and provide a new direction for the future treatment and development of neuroblastoma.
neuroblastoma,children,TIAM1,differentiation,research progress
10.3969/j.issn.1000-8179.2017.22.032
天津醫科大學腫瘤醫院兒童腫瘤科,國家腫瘤臨床醫學研究中心,天津市腫瘤防治重點實驗室,天津市惡性腫瘤臨床醫學研究中心(天津市300060)
趙強 qiangzhao169@sina.com
(2017-09-08收稿)
(2017-10-31修回)
(編輯:孫喜佳 校對:周曉穎)

邱艷麗 專業方向為兒童惡性實體瘤的分子診斷。
E-mail:qyl0611@126.com
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
汽車工程學報(2017年2期)2017-07-05 08:13:02
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
