力達霉素對人宮頸癌Caski細胞增殖的影響及其機制
2017-12-11 06:02:50魯華秦燁占景瓊王艷林
山東醫藥 2017年44期
魯華,秦燁,占景瓊,王艷林
(三峽大學腫瘤微環境與免疫治療湖北省重點實驗室,湖北宜昌443002)
·論著·
力達霉素對人宮頸癌Caski細胞增殖的影響及其機制
魯華,秦燁,占景瓊,王艷林
(三峽大學腫瘤微環境與免疫治療湖北省重點實驗室,湖北宜昌443002)
目的探討力達霉素對人宮頸癌Caski細胞增殖的影響及其可能的機制。方法將對數生長期的人宮頸癌Caski細胞隨機分為低、中、高濃度組及對照組,低、中、高濃度組均加入含力達霉素的RPMI 1640完全培養基100 μL,分別調整其終濃度為1、2.5、5 ng/mL,對照組僅加入等體積RPMI 1640完全培養基。作用24 h,采用MTT法檢測細胞增殖能力(以吸光度值表示),計算細胞生存率;采用流式細胞術檢測細胞周期及線粒體膜電位降低比例;透射電子顯微鏡下觀察細胞形態學改變,計算細胞凋亡率及裂亡率。結果低、中、高濃度組細胞增殖能力及細胞生存率均低于對照組,且高濃度組降低更明顯(P均<0.01)。對照組及中濃度組S期細胞比例均高于低、高濃度組(P均<0.05)。低、中、高濃度組G0/G1期細胞比例均低于對照組,G2/M期細胞比例均高于對照組,且中、高濃度組變化更明顯(P均<0.05)。低、中、高濃度組及對照組細胞線粒體膜電位降低比例分別為(34.77±5.33)%、(37.87±5.73)%、(18.37±3.91)%、(4.17±0.65)%;低、中、高濃度組細胞線粒體膜電位降低比例均高于對照組,且低、中濃度組均高于高濃度組(P均<0.05)。透射電鏡下可見對照組細胞形態正常,低、中、高濃度組存在多種形態的死亡細胞,包括凋亡、壞死、裂亡的細胞。對照組及高、低、中濃度組細胞凋亡率及裂亡率均依次升高,兩組間比較P均<0.05。結論力達霉素可抑制人宮頸癌Caski細胞增殖,并呈濃度依賴性,其機制可能與誘導細胞凋亡或裂亡有關。
宮頸癌;力達霉素;人宮頸癌Caski細胞;細胞凋亡;細胞裂亡;線粒體膜電位
力達霉素是一種從鏈霉菌中分離的大分子肽類抗生素[1],對多種腫瘤(如肺癌、胰腺癌、肝癌等)具有強烈的細胞毒作用[2~6],其在細胞水平上的抗腫瘤活性顯著高于臨床上常用的抗腫瘤藥物,如阿霉素、絲裂霉素C、甲氨蝶呤等。研究表明,力達霉素可抑制肺癌、結腸癌、乳腺癌、胃癌、肝癌等細胞移植后的裸鼠成瘤能力,并通過多種機制抑制腫瘤生長,且可能對腫瘤細胞的增殖具有特異性影響[2~5]。2015年3月~2016年6月,本研究觀察了力達霉素對人宮頸癌Caski細胞增殖的影響,現分析結果并探討其可能的機制。
1 材料與方法
1.1 材料 細胞:人宮頸癌Caski細胞購于武漢大學細胞典藏中心,由三峽大學腫瘤微環境與免疫治療湖北省重點實驗室傳代保存。主要藥物及試劑:力達霉素由中國醫學科學院生物技術研究所甄永蘇院士惠贈,MitoProbe TM JC-1流式細胞術檢測試劑盒及RPMI 1640完全培養基均購于美國Invitrogen公司。主要儀器:全波長酶標儀購于美國Thermo Fisher公司;H-7500透射電子顯微鏡購于日本日立公司;超薄切片機購于德國萊卡公司;EPICSXL-4流式檢測儀購于美國Beckman-coulter公司。
1.2 細胞培養與分組處理 取對數生長期的人宮頸癌Caski細胞,以3 000個/孔接種至96孔板,采用RPMI 1640完全培養基,于37 ℃、5% CO2培養箱中培養20 h。將人宮頸癌Caski細胞隨機分為低、中、高濃度組及對照組,低、中、高濃度組均加入含力達霉素的RPMI 1640完全培養基100 μL,分別調整其終濃度為1、2.5、5 ng/mL,對照組僅加入等體積RPMI 1640完全培養基,37 ℃、5% CO2培養箱中培養。每組設置4個復孔。
1.3 細胞增殖情況檢測 采用MTT法。取各組作用24 h細胞,每孔加入含0.5% MTT的RPMI 1640完全培養基(終濃度為250 μg/mL),繼續培養4 h。棄上清,每孔加入250 μL二甲基亞砜,置搖床上低速振蕩10 min,充分溶解結晶物。采用酶聯免疫檢測儀測量490 nm波長處的吸光度(OD)值,以此表示細胞增殖能力,并計算細胞生存率。細胞生存率=OD藥物/OD對照×100%。
1.4 細胞周期檢測 采用流式細胞術。收集各組作用24 h細胞,離心棄上清,預冷PBS沖洗2次,預冷70%乙醇4 ℃固定30 min。2 000 r/min離心5 min,棄上清,1 mL PBS沖洗1次。采用含0.2% TritonX-100和100 μg/mL RNase的PBS混合液500 μL重懸細胞,加入碘化丙啶(50 μg/ mL)避光孵育30 min,300目尼龍網進行過濾,上流式細胞儀檢測細胞周期。
1.5 細胞凋亡情況檢測 采用流式細胞術。收集各組作用24 h細胞,用預溫至37 ℃的培養基1 mL重懸細胞,邊振蕩邊加入2.5 mL熒光染料JC-1儲存液(終濃度2.5 μg/mL),37 ℃避光保存10 min。加入2 mL PBS至細胞懸液中,2 000 r/min離心5 min,棄上清,0.3 mL PBS重懸細胞。上流式細胞儀檢測細胞線粒體膜電位降低比例,以此反映細胞凋亡情況。
1.6 細胞形態學改變及死亡細胞分類 收集各組作用24 h細胞,2 000 r/min離心5 min,棄上清,戊二醛溶液固定,超薄切片機制片。以200目透射電鏡銅網為載體,隨機選取10目,采用透射電子顯微鏡在不同放大倍數下(×5 000、×6 000、×7 000)觀察細胞形態學改變,計數凋亡及裂亡細胞,計算細胞凋亡率及裂亡率。
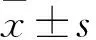
2 結果
2.1 各組細胞增殖情況比較 低、中、高濃度組細胞增殖能力及細胞生存率均低于對照組,且高濃度組降低更明顯(P均<0.01)。見表1。
2.2 各組細胞周期比較 對照組及中濃度組S期細胞比例均高于低、高濃度組(P<0.05或<0.01)。低、中、高濃度組G0/G1期細胞比例均低于對照組,G2/M期細胞比例均高于對照組,且中、高濃度組變化更明顯(P<0.05或<0.01)。見表2。
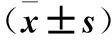
表1 各組細胞增殖情況比較
注:與對照組比較,*P<0.01;與高濃度組比較,#P< 0.01。
表2 各組細胞周期比較
注:與對照組比較,*P<0.05,#P<0.01;與低濃度組比較,△P<0.05,▽P<0.01;與中濃度組比較,▲P<0.05。
2.3 各組細胞凋亡情況比較 低、中、高濃度組及對照組細胞線粒體膜電位降低比例分別為(34.77±5.33)%、(37.87±5.73)%、(18.37±3.91)%、(4.17±0.65)%;低、中、高濃度組細胞線粒體膜電位降低的細胞比例均高于對照組,且低、中濃度組升高更明顯(P均<0.05);低、中濃度組細胞線粒體膜電位降低的細胞比例比較差異無統計學意義(P>0.05)。
2.4 各組細胞形態學變化 對照組細胞膜完整,向外形成大量纖細突起的微絨毛;細胞質染色正常,可見線狀、長桿狀線粒體;細胞核常染色質(低電子密度)、異染色質(高電子密度)分布均勻。低、中、高濃度組存在多種形態的死亡細胞,包括凋亡、壞死、裂亡的細胞。死亡細胞的主要特點:①凋亡中期細胞體積變小,細胞核深染,異染色質邊集,核膜和細胞質膜完整,細胞質內可見空泡形成,細胞膜上可見以發芽、起泡等方式形成的多個球形突起。凋亡晚期細胞體積進一步變小,細胞質少而深,細胞體周圍可見多個球形突起脫落形成的一些大小不等的凋亡小體。②壞死早期細胞核染色質邊集,核中部電子密度低;細胞質結構崩解,呈顆粒狀;線粒體、內質網崩解形成空泡狀結構,可見其中含有中等電子密度的無定形電子致密物。壞死晚期細胞膜破裂,細胞質逸出;細胞微絨毛消失,細胞質結構崩解,呈疏松顆粒狀;細胞器崩解空泡形成,染色質變淡呈顆粒狀凝集,只可見到細胞核輪廓。③裂亡早期細胞體積變大,細胞有絲分裂異常形成大小不等多個細胞核。裂亡晚期細胞體積進一步變大,細胞表面光滑,微絨毛消失,細胞質內充滿線粒體,內質網腫脹崩解形成的空泡狀結構;細胞核常染色質與異染色質差異消失,呈砂礫樣改變。
2.5 各組細胞凋亡率及裂亡率比較 對照組及高、低、中濃度組細胞凋亡率依次升高,對照組及低、中、高濃度組細胞裂亡率依次升高,兩組間比較P<0.05或<0.01。見表3。

表3 各組細胞凋亡率及裂亡率比較
注:與對照組比較,*P<0.05,#P<0.01;與低濃度組比較,△P<0.05,▽P<0.01;與中濃度組比較,▲P<0.05,▼P<0.01。
3 討論
力達霉素是一種新型抗腫瘤藥物,其對肝癌BEL-7402細胞DNA、RNA合成具有強烈的抑制作用[7],對結腸癌HCT-116細胞具有明顯的細胞周期阻滯作用(阻滯于G2/M期)[8],并能顯著誘導早幼粒白血病HL-60細胞、胰腺癌細胞、骨髓瘤U266及SKO-007細胞凋亡[9~11],引起肝癌BEL-7420細胞發生裂亡[12]。本研究結果顯示,低、中、高濃度組細胞增殖能力及細胞生存率均低于對照組,且高濃度組降低更明顯;說明低濃度力達霉素處理24 h即能顯著抑制人宮頸癌Caski細胞增殖,且該作用具有濃度依賴性。
線粒體膜電位是指生物膜兩側離子濃度不同所產生的跨膜電位差,可反映線粒體功能的完整性,是評價線粒體功能的敏感指標[13]。細胞死亡方式有凋亡和裂亡等不同形式[14,15],細胞凋亡時線粒體損傷發生在細胞核凋亡特征(染色質濃縮、DNA斷裂)出現之前[16]。在凋亡因子的刺激下,線粒體膜通透性轉化孔增大,使內膜離子濃度梯度消失、呼吸鏈失偶聯,導致跨膜電位降低或耗散,促進凋亡活性物質如Cytc等從線粒體基質中釋放進入細胞質,激活Caspase級聯反應,最終引發細胞凋亡[17]。因此,線粒體膜電位降低是細胞凋亡早期的一個標志性事件,線粒體膜電位降低細胞比例可反映細胞凋亡的程度[16]。本研究結果顯示,低、中、高濃度組線粒體膜電位降低的細胞比例均高于對照組,提示不同濃度的力達霉素均可導致線粒體膜電位顯著下降,并誘導細胞發生線粒體介導的凋亡。本研究中高濃度組細胞線粒體膜電位下降比例低于中、低濃度組,細胞線粒體膜電位下降比例并沒有隨力達霉素濃度的升高而升高,結合力達霉素對人宮頸癌Caski細胞的增殖抑制作用呈濃度依賴性這一結果,推測力達霉素除了誘導人宮頸癌Caski細胞凋亡外,還可能誘導Caski細胞發生其他類型的死亡。
1989年,Molz等[18]發現在一種對熱敏感的酵母突變株中,細胞分裂時染色體分離發生異常。此后一些學者便把這種在DNA發生損害時,細胞無法進行完全分裂而導致出現四倍體或多倍體的現象稱為細胞有絲分裂災變,這種有別于典型細胞凋亡特征的細胞死亡方式也被稱為裂亡[19]。裂亡的發生可能與細胞有絲分裂的異常,如中心體過度復制、多極性紡錘體形成、多核形成等有關[20],細胞周期檢測表現為S期和G2/M期阻滯[21]。本研究結果顯示,中、高濃度組G0/G1期細胞比例均低于對照組及低濃度組,G2/M期細胞比例均高于對照組及低濃度組,對照組及中濃度組S期細胞比例均高于低、高濃度組。說明力達霉素除了誘導人宮頸癌Caski細胞凋亡外,還可誘導細胞發生裂亡。
除了生化特征外,裂亡與凋亡細胞還具有各自不同的形態學特征。凋亡細胞染色質凝集、邊緣化,細胞體皺縮、變圓,形成凋亡小體;裂亡的主要形態學特征包括有絲分裂異常,細胞體積變大,形成多核細胞。本研究透射電鏡觀察結果證實,力達霉素處理后的人宮頸癌Caski細胞形態學上發生上述凋亡、壞死及裂亡病理改變。本研究結果顯示,對照組及高、低、中濃度組細胞凋亡率依次升高,對照組及低、中、高濃度組細胞裂亡率依次升高;說明力達霉素濃度為2.5 ng/mL時對細胞凋亡的誘導作用最強,濃度為5 ng/mL時對細胞裂亡的誘導作用最強。上述結果提示,不同濃度力達霉素誘導人宮頸癌Caski細胞死亡的模式也有所不同。
綜上所述,力達霉素可抑制人宮頸癌Caski細胞增殖,并呈濃度依賴性;其機制可能與誘導人宮頸癌Caski細胞凋亡或裂亡有關。
[1] Hu JL, Xue YC, Xie MY, et al. A new macromolecular antitumor antibiotic, C1027. I. Discovery, taxonomy of producing organism, fermentation and biological activity[J]. J Antibiot, 1988,41(11):1575-1584.
[2] Liu F, Shang Y, Chen SZ. Chloroquine potentiates the anti-cancer effect of lidamycin on non-small cell lung cancer cells in vitro[J]. Acta Pharmacol Sin, 2014,35(5):645-652.
[3] Chen J, Wu S, Ou-Yang ZG, et al. Synergy of gemcitabine and lidamycin associated with NF-κBdownregulation in pancreatic carcinoma cells[J]. Acta Pharmacol Sin, 2008,29(5):614-619.
[4] Huang YH, Shang BY, Zhen YS. Antitumor efficacy of lidamycin on hepatoma and active moiety of its molecule[J]. World J Gastroenterol, 2005,11(26):3980-3984.
[5] Zhang SH, Chen J, Jiang M, et al. Lidamycin induces apoptosis of human gastric carcinoma BGC823 cells and inhibits xenograft growth in nude mice[J]. Acta Pharmacol Sin, 2008,43(6):601-604.
[6] 陳淑珍,甄永蘇,邵榮光.力達霉素抗腫瘤作用及其分子機制研究新進展[J].中國抗生素雜志,2010,35(6):401-407,413.
[7] He QY, Jing B, Li Dd. Effects of lidamycin on genomic DNA in human heptatoma BEL-7402 cells[J]. Acta Pharmacol Sin, 2002,23(3):253-256.
[8] Pan Y, Ren K, He H, et al. Knockdown of Chk1 sensitizes human colon carcinoma HCT116 cells in a p53-dependent manner to lidamycin through abrogation of a G2/M checkpoint and induction of apoptosis[J]. Cancer Biol Ther, 2009,8(16):1559-1566.
[9] Chen J, Yan ZY, Li C, et al. Effects of lidamycin on proliferation and differentiation of HL-60 Cell[J]. J Chin Pharm Sci, 2010,45(23):819-1822.
[10] Chen J, Ou-Yang ZG, Zhang SH, et al. Down-regulation of the nuclear factor-κB by lidamycin in association with inducing apoptosis in human pancreatic cancer cells and inhibiting xenograft growth[J]. Oncol Rep, 2007,17(6):1445-1451.
[11] Zhen YZ, Ji CM, Hao XF, et al. Lidamycin enhances sensitivity of multiple myeloma to bortezomid by modulating expression of mitogen-activated protein kinases[J]. Cancer Res Prevent Treat, 2014,41(5):353-357.
[12] Vitale I, Galluzzi L, Castedo M, et al. Mitotic catastrophe: a mechanism for avoiding genomic instability[J]. Nat Rev Mol Cell Biol, 2011,12(6):384-391.
[13] 劉禎,李衛紅,張杰,等.魚藤酮對神經瘤細胞線粒體膜電位的影響[J].環境與職業醫學,2003,20(2):73-74,77.
[14] 高瑞娟,趙春燕,李電東.有絲分裂與有絲分裂災變[J].中國新藥雜志,2014,24(8):2854-2859,2885.
[15] Jiang B, Li DD, Zhen YS. Induction of apoptosis by enediyne antitumor antibiotic C1027 in HL-60 human promyelocytic leukemia cells[J]. Biochem Biophys Res Com, 1995,208(1):238-244.
[16] Haeberlein SL. Mitochondrial function in apoptotic neuronal cell death[J]. Neurochem Res, 2004,29(3):521-530.
[17] Garrido C, Galluzzi L, Brunet M, et al. Mechanisms of cytochrome crelease from mitochondria[J]. Cell Death Differ, 2006,13(9):1423-1431.
[18] Molz L, Booher R, Young P, et al. cdc2 and the regulation of mitosis: six interacting mcs genes[J]. Genetics, 1989,5(4):365.
[19] Margottin-Goguet F, Hsu JY, Loktev A, et al. Prophase destruction of emi1 by the SCF βTrCP/Slimb, ubiquitin ligase activates the anaphase promoting complex to allow progression beyond prometaphase[J]. Dev Cell, 2003,4(6):813-826.
[20] Liang YX, Zhang W, Li DD, et al. Mitotic cell death in BEL-7402 cells induced by enediyne antibiotic lidamycin is associated with centrosome overduplication[J]. World J Gastroenterol, 2004,10(18):2632-2636.
[21] Ianzini F, Mackey MA. Spontaneous premature chromosome condensation and mitotic catastrophe following irradiation of HeLa S3 cells[J]. Int J Radiat Biol, 1997,72(4):409-421.
EffectoflidamycinonproliferationofhumancervicalcancerCaskicells
LUHua,QINYe,ZHANJingqiong,WANGYanlin
(ChinaThreeGorgesUniversity,HubeiKeyLaboratoryofTumorMicroenvironmentandImmunotherapy,Yichang443002,China)
ObjectiveTo investigate the effect of lidamycin on the proliferation of human cervical cancer Caski cells and its underlying molecular mechanisms.MethodsHuman cervical cancer Caski cells were randomly divided the experimental groups (including the low-dose, medium-dose and high-dose groups) which were separately treated with 1, 2.5 and 5 ng/mL lidamycin (100 μL) for 24 h, and the control group which was added with the same volume of RPMI-1640. MTT assay was used to detect the inhibition rate of proliferation. The cell cycle and mitochondrial membrane potential of CasKi cells were measured by flow cytometry. Transmission electronic microscope was used to observe the morphological changes of Caski cells. We calculated the apoptosis rate and mitotic catastrophe rate.ResultsThe proliferation rate and survival rate of the experimental groups were significantly lower than those of the control group, and the decrease in the high-dose group was more significant (allP<0.05). The percentage of cells in the S phase of the control group and the medium-dose group was higher than those in the low-dose and high-dose groups (allP<0.05). The percentage of cells in the G0/G1phase of the experimental groups was lower than that of the control group and the percentage of cells in the G2/M phase was higher than that of the control group, especially in the medium-dose and high-dose groups (allP<0.05). The ratios of mitochondrial membrane potential decrease in the low-dose, medium-dose and high-dose groups and control group were 34.77%±5.33%, 37.87%±5.73%, 18.37%±3.91%, and 4.17%±0.65%, respectively. The mitochondrial membrane potential decreased ratios of the experimental groups were higher than that in the control group, especially in the low-dose and medium-dose groups (allP<0.05). Transmission electronic microscopy showed that the control group had normal morphology while the experimental groups had many forms of death cells, including apoptosis, necrosis and mitotic catastrophe. The apoptosis rate and mitotic catastrophe rate of the experimental groups were both higher than those of control group (bothP<0.05).ConclusionLidamycin can significantly inhibit the proliferation of Caski cells with an dose-dependent manner by inducing the apoptosis and mitotic catastrophe of cells.
cervical carcinoma; lidamycin; human cervical cancer Caski cells; apoptosis; mitotic catastrophe; mitochondrial membrane potential
國家自然科學基金資助項目(81372265)。
魯華(1978-),男,講師,研究方向為腫瘤病理學。E-mail: 4554080@qq.com
王艷林(1954-),男,教授,研究方向為腫瘤分子生物學。E-mail: 2244524933@qq.com
10.3969/j.issn.1002-266X.2017.44.001
R737.33
A
1002-266X(2017)44-0001-04
2017-02-09)
