老鴉瓣屬植物資源及分子系統(tǒng)學(xué)研究進(jìn)展
2017-11-09 01:14:22陳紹通宋向文韓邦興
皖西學(xué)院學(xué)報(bào) 2017年5期
陳紹通,劉 楓,宋向文,趙 群,韓邦興
(1.安徽中醫(yī)藥大學(xué),安徽 合肥 230012;2.皖西學(xué)院,安徽 六安 237012;3.安徽省石斛產(chǎn)業(yè)化開(kāi)發(fā)協(xié)同創(chuàng)新中心,安徽 六安 237012)
老鴉瓣屬植物資源及分子系統(tǒng)學(xué)研究進(jìn)展
陳紹通1,2,劉 楓1,2,宋向文2,3,趙 群2,3,韓邦興2,3
(1.安徽中醫(yī)藥大學(xué),安徽 合肥 230012;2.皖西學(xué)院,安徽 六安 237012;3.安徽省石斛產(chǎn)業(yè)化開(kāi)發(fā)協(xié)同創(chuàng)新中心,安徽 六安 237012)
老鴉瓣屬AmanaHonda為百合科藥用植物屬,為東亞特有類(lèi)群。有關(guān)老鴉瓣屬的研究相對(duì)較少,且多集中于物種老鴉瓣Amanaedulis(Miq.) Honda的化學(xué)成分、組培、生物學(xué)特性等方面,而對(duì)于老鴉瓣屬內(nèi)其他種的研究報(bào)道極少。本文主要綜述了該屬植物的系統(tǒng)位置、屬內(nèi)種的分類(lèi)學(xué)地位、生物學(xué)特性及分子系統(tǒng)學(xué)等多個(gè)方面的研究進(jìn)展。基于老鴉瓣屬植物已有的和本課題組前期初步研究的基礎(chǔ),并針對(duì)該屬中存在的關(guān)鍵科學(xué)問(wèn)題,對(duì)可能采用的研究方法提出展望,其研究思路為老鴉瓣屬藥用植物資源開(kāi)發(fā)奠定基礎(chǔ)。
老鴉瓣;老鴉瓣屬;植物資源;生物學(xué)特性;分子系統(tǒng)學(xué)
藥用植物老鴉瓣Amanaedulis(Miq.) Honda隸屬于郁金香屬AmanaHonda,其去除絨毛后的干燥鱗莖為中藥“光慈姑”的來(lái)源,具有解毒散結(jié)、行血化瘀的功效,主治咽喉腫痛、瘰疬和產(chǎn)后瘀滯等癥狀[1](P361)。由于文獻(xiàn)記載混亂等原因,老鴉瓣的藥用歷史也經(jīng)歷了一系列的變遷。在晉代,鱗莖作為“貝母”入藥,但因其有毒,被列為“貝母”的偽品,稱為“丹龍睛”;到了宋代,又作為藥材“山慈姑”來(lái)源入藥。此外,古代老鴉瓣鱗莖入藥并不去除絨毛,因此又稱“毛慈姑”,直到近代,人們?cè)诓墒占庸さ臅r(shí)候,為了提高藥材的療效,去除毛殼而改稱“光慈姑”[2]。
老鴉瓣屬AmanaHonda隸屬于百合科Liliaceae,由Honda從郁金香屬TulipaL.分離而成立[3],為東亞特有類(lèi)群,僅分布于中國(guó)中東部,日本和朝鮮半島[4]。老鴉瓣屬在百合科中分類(lèi)地位雖然得到了一些學(xué)者的承認(rèn)[5-6],[7](P38),[8](P925),但對(duì)于老鴉瓣屬是另立還是歸入郁金香屬Tulipa的爭(zhēng)論仍然存在。此外,面對(duì)市場(chǎng)對(duì)老鴉瓣的需求量日益增加,而野生自然資源正逐漸下降等矛盾,如何提高老鴉瓣的產(chǎn)量成為人們?cè)絹?lái)越關(guān)注的問(wèn)題。而人工栽培、組織培養(yǎng)、尋找新藥源等方法也成為當(dāng)前解決問(wèn)題的重要途徑。因此,本文基于課題組前期對(duì)老鴉瓣屬資源的調(diào)查及初步研究,對(duì)老鴉瓣屬的分類(lèi)、生物學(xué)特性及分子系統(tǒng)學(xué)進(jìn)行綜述,為老鴉瓣屬的分類(lèi)及老鴉瓣栽培提供了數(shù)據(jù)支持,也為尋找新藥源及今后產(chǎn)業(yè)化的發(fā)展奠定基礎(chǔ)。
1 老鴉瓣屬植物資源
1.1 資源特征及分布
老鴉瓣屬Amana隸屬于百合科,多年生草本,由Honda從郁金香屬分離而成立,在形態(tài)上與郁金香屬的主要區(qū)別在于:花被片背面具有紫紅色縱向條紋,花柱與子房近等長(zhǎng),具2~4個(gè)對(duì)生或輪生苞片,葉常2枚對(duì)生且兩面無(wú)毛[4]。目前,已發(fā)現(xiàn)的該類(lèi)群共5種,分別為老鴉瓣Amanaedulis、二葉郁金香Amanaerythronioides、安徽老鴉瓣Amanaanhuiensis、括蒼山老鴉瓣Amanakuocangshanica和皖浙老鴉瓣Amanawanzhensis[9](P89) [10-13]。該屬中植物均為早春植物,繁盛生長(zhǎng)期為每年的3~4月,5月開(kāi)始倒苗,喜溫暖潮濕環(huán)境,以老鴉瓣A.edulis分布最廣,而屬中其他物種分布較為狹窄且集中,主要以安徽、浙江地區(qū)為分布中心,多為各個(gè)地區(qū)的特有物種,可見(jiàn)圖1。
老鴉瓣Amanaedulis(Miq.) Honda為多年生草本,葉通常兩片,線形,長(zhǎng)達(dá)30 cm;花單朵頂生;苞片通常2枚對(duì)生,地下部分為橢圓形鱗莖,外包赤褐色膜質(zhì)鱗片,內(nèi)密被長(zhǎng)柔毛。我國(guó)中東部大部分地區(qū)(安徽、江蘇、浙江、山東、江西、湖北、湖南、陜西、遼寧)都有分布,且多生長(zhǎng)在丘陵、崗地等向陽(yáng)的草地或山坡上。目前也作為治療腫瘤的常用藥[14]。

圖1 中國(guó)老鴉瓣屬植物主要分布區(qū)域
二葉郁金香Amanaerythronioides(Baker) D. Y. Tan et D. Y. Hong,又稱二葉老鴉瓣、寬葉老鴉瓣,與老鴉瓣相似,同為多年生草本,以葉片和苞片類(lèi)型而區(qū)別于同屬的其他種。本種葉兩枚,較寬而短,長(zhǎng)度在7~15 cm之間,兩片葉寬也不相同,寬者常在15~22 mm,窄者常為9~15 mm;苞片通常3或4枚輪生(較少2枚對(duì)生)。多生長(zhǎng)于海拔600~1 300 m的高山,目前,除安徽、浙江之外,2012年,在江西省云居山也有發(fā)現(xiàn),為江西地理新分布物種[15]。
2001年,沈顯生在安徽天柱山發(fā)現(xiàn)郁金香屬的一新種,并命名為皖郁金香TulipaanhuiensisX. S. Shen。2008年,潭敦炎[16]等對(duì)該種新模式進(jìn)行指定和補(bǔ)充,并重新組合命名為安徽老鴉瓣Amanaanhuiensis(X. S. Shen.) D. Y. Tan et D. Y. Hong。該種為安徽天柱山地區(qū)的特有種,株高7.5~15 cm之間,生長(zhǎng)在海拔850~1250 m的落葉闊葉林下,溪溝邊或灌木叢中,陰濕環(huán)境下生長(zhǎng)較多。與同屬其余種的形態(tài)差異在于鱗莖皮薄紙質(zhì),內(nèi)側(cè)被綿毛;苞片(2~)3(~4)枚,輪生(或?qū)ι?,第一葉長(zhǎng)9.6~22.4 cm,寬1.6~2.5 cm,且從基部向上3/4處最寬(潭敦炎等, 2008)。沈顯生認(rèn)為本種與二葉郁金香A.erythronioides近緣,但花被片披針形,花藥淡紫色,花柱1~2 mm,柱頭歪斜可與之區(qū)別[11]。
2007年,Tan等在浙江括蒼山地區(qū)發(fā)現(xiàn)一新種——括蒼山老鴉瓣AmanakuocangshanicaD. Y. Tan & D. Y. Hong,分布在海拔600~1 100 m。在形態(tài)學(xué)上,該種與安徽老鴉瓣和二葉郁金香相似,不同的是地下鱗莖部分皮內(nèi)側(cè)無(wú)毛,葉片最寬處在基部向上的2/3處,果喙長(zhǎng)(6.4±0.8)mm。此外,TAN通過(guò)17個(gè)形態(tài)特征對(duì)A.edulis、A.erythronioides、A.anhuiensis、A.kuocangshanica進(jìn)行UPGAM聚類(lèi)分析,結(jié)果發(fā)現(xiàn)A.edulis聚在最外支,A.anhuiensis與A.kuocangshanica互為姐妹群,并與A.erythronioides形成并系分支[12]。
2014年,Han等人在安徽寧國(guó)的仙霞鎮(zhèn)發(fā)現(xiàn)老鴉瓣屬的另一新種——皖浙老鴉瓣AmanawanzhensisL. Q. Huang, B. X. Han & K. Zhang。該種多生長(zhǎng)在600~800 m的竹林下、灌叢中或草地上,鱗片卵形,直徑1.5~2.5 cm,內(nèi)部棕色紙質(zhì),柔毛,莖高15~30 cm,苞片通常3枚,不輪生,帶狀,長(zhǎng)0.1~0.5 cm。Han等認(rèn)為該種與二葉郁金香A.erythronioides相似,但通過(guò)苞片長(zhǎng)度較短(0.1~0.5cm),花藥黃色,花被片易脫落可與其區(qū)分開(kāi)。而該結(jié)論也獲得了分子系統(tǒng)學(xué)的支持,通過(guò)ITS+trnL intron聯(lián)合序列對(duì)該屬(除括蒼山老鴉瓣A.kuocangshanica)之外的其余四種進(jìn)行分子發(fā)育研究,結(jié)果顯示該種獨(dú)自形成單支,且獲得了高度的支持(BS=93, PP=1.00),相對(duì)于老鴉瓣,皖浙老鴉瓣與二葉郁金香聚為一支(BS=79, PP=0.95),由此也說(shuō)明了,該新種是成立的,且與二葉郁金香親緣關(guān)系更近[13]。
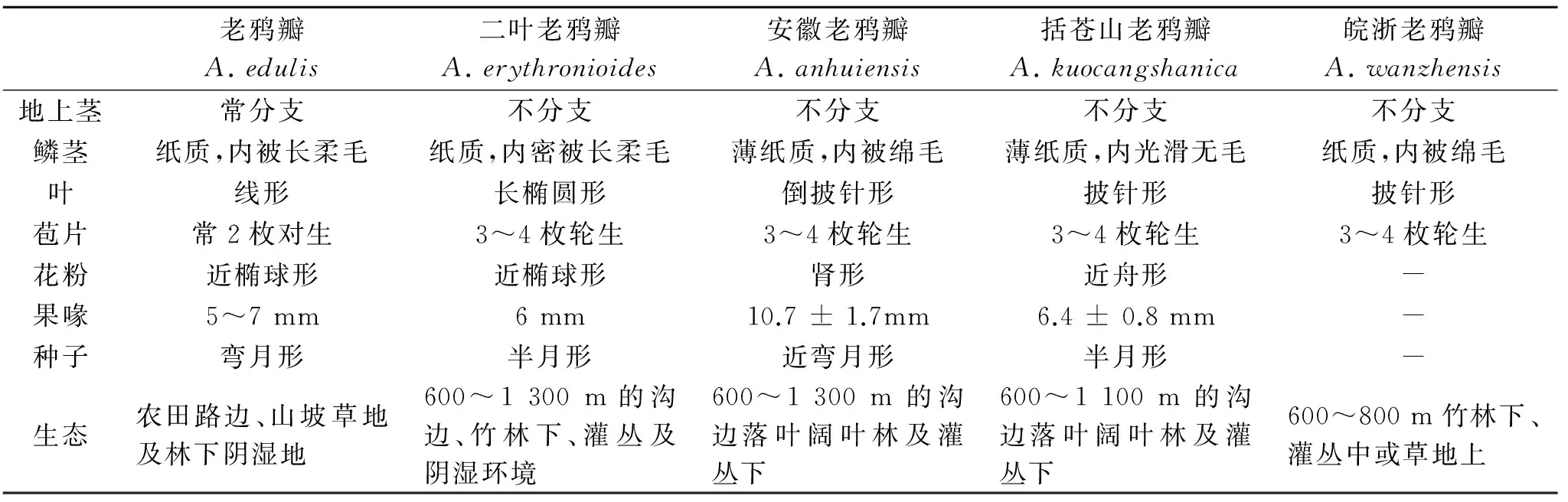
表1 老鴉瓣屬內(nèi)部形態(tài)特征比較[17]
1.2 老鴉瓣屬植物分類(lèi)沿革
近年來(lái),對(duì)于老鴉瓣屬Amana的系統(tǒng)發(fā)育和分類(lèi)地位一直備受爭(zhēng)議。自Honda建屬以來(lái),關(guān)于該類(lèi)群是隸屬于郁金香屬Tulipa還是另立的問(wèn)題也一直是爭(zhēng)論的焦點(diǎn)。其中,較為重要的分類(lèi)方法當(dāng)屬M(fèi)iquel[18],Baker[19],Honda[5],Hall[3],Sealy[20],Hoog[21],毛祖美[22](P86-97),Kitamura[6],Satake[7],Ohwi和Kitagawa[23],梁松筠[24],Tamura[25],吳征鎰[8],譚敦炎[4],韓邦興[13]等,可見(jiàn)表2。

表2 老鴉瓣屬分類(lèi)研究
1.3 生物學(xué)特性
老鴉瓣屬為早春植物,對(duì)光照、溫度等自然條件要求較高,多生長(zhǎng)在向陽(yáng)的山坡、草地和水邊[26]。除了老鴉瓣之外,其余種均分布在海拔較高的山區(qū),如二葉郁金香多生長(zhǎng)在海拔600~1300 m的落葉闊葉林下,安徽老鴉瓣生長(zhǎng)在海拔850~1250 m的竹林、落葉闊葉林或水邊,部分也長(zhǎng)在向陽(yáng)的石頭含有腐殖質(zhì)較多的潮濕處。皖浙老鴉瓣多生長(zhǎng)在600~800 m的竹林下、灌叢中或草地上。而老鴉瓣在平原、路邊及高海拔地區(qū)都有分布。目前,由于老鴉瓣屬物種分布的限制,在對(duì)于老鴉瓣屬的生物學(xué)特性的研究中,以老鴉瓣的研究最多,而其余種的研究極少見(jiàn)到報(bào)道。
老鴉瓣具有典型的百合科花部特征,屬兼性異交類(lèi)型,異交為主,自交為輔,主要以昆蟲(chóng)為老鴉瓣傳粉媒介[27]。梁麗[28]等對(duì)老鴉瓣的光合特性進(jìn)行研究發(fā)現(xiàn),老鴉瓣的光補(bǔ)償點(diǎn)為1.782 umol/(m2·s),光飽和點(diǎn)約為1 500 umol/(m2·s),表現(xiàn)光量子產(chǎn)額為0.025,說(shuō)明老鴉瓣具有陰性的特點(diǎn),對(duì)光能的生態(tài)適應(yīng)范圍較廣,但不宜強(qiáng)光直射。此外有研究發(fā)現(xiàn),在78%左右的光強(qiáng)最適宜老鴉瓣生長(zhǎng)[29]。
老鴉瓣種子屬胚形態(tài)休眠為主的綜合休眠型,且剛成熟老鴉瓣種子胚率僅為10%左右。吳正軍[27]等為提高種子萌發(fā)率,對(duì)老鴉瓣種子的萌發(fā)特性進(jìn)行研究,結(jié)果發(fā)現(xiàn),通過(guò)300 mg·L-1赤霉素浸泡處理24 h后,25 ℃恒溫層積70天,再轉(zhuǎn)入5 ℃低溫層積可使種胚成熟最快,破眠之后的種子在10、15 ℃的黑暗條件下萌發(fā)率可達(dá)90%以上。
除了種子繁殖之外,老鴉瓣也可通過(guò)鱗莖繁殖,而且相對(duì)于種子繁殖,鱗莖繁殖過(guò)程所需周期更短。但是,與種子繁殖相同的是,如何打破鱗莖休眠,延長(zhǎng)老鴉瓣的生長(zhǎng)發(fā)育時(shí)間是提高產(chǎn)量的關(guān)鍵。楊穎[30]等發(fā)現(xiàn)冷藏處理有利于鱗莖提前出苗,并顯著縮短出苗時(shí)間且利于苗齊,但對(duì)出苗率影響不明顯,且冷藏后花發(fā)育不正常,結(jié)果率為0,并認(rèn)為(4±0.5)℃冷藏6~8周為老鴉瓣鱗莖打破休眠的關(guān)鍵時(shí)期,冷藏可延長(zhǎng)老鴉瓣生育期并提高鱗莖產(chǎn)量。繆媛媛[31]等通過(guò)摘蕾和人工授粉處理老鴉瓣,發(fā)現(xiàn)摘蕾有利于老鴉瓣鱗莖生物量的積累從而提高藥材產(chǎn)量,而人工授粉顯著提高老鴉瓣結(jié)實(shí)率及種子數(shù)量和單粒重。楊小花[32]等根據(jù)鱗莖的重量設(shè)4個(gè)級(jí)別:1級(jí)(重量>2 g)、2級(jí)(重量=1~2 g)、3級(jí)(重量=0.5~1 g)、4級(jí)(重量<0.5 g),并在生長(zhǎng)旺期測(cè)量老鴉瓣的生長(zhǎng)指標(biāo),結(jié)果發(fā)現(xiàn)3級(jí)和4級(jí)種球增產(chǎn)效果明顯,作為鱗莖繁殖最佳。同時(shí),作者也對(duì)老鴉瓣的采收期研究發(fā)現(xiàn),老鴉瓣應(yīng)在枯萎期或休眠期初期采收為佳[33]。除了上述之外,對(duì)于老鴉瓣的組織培養(yǎng)也有報(bào)道[34-35]。
2 老鴉瓣屬分子系統(tǒng)學(xué)研究
近年來(lái),隨著測(cè)序技術(shù)的迅猛發(fā)展和大量的DNA數(shù)據(jù)積累,生物系統(tǒng)學(xué)也由早期的利用化石證據(jù)、表型形態(tài)特征或者解剖結(jié)構(gòu)性狀推演生物間系統(tǒng)關(guān)系,轉(zhuǎn)入分子系統(tǒng)學(xué)時(shí)代[36]。目前,分子系統(tǒng)學(xué)已成為植物系統(tǒng)發(fā)育中的研究熱點(diǎn)。相對(duì)于傳統(tǒng)的方法,分子系統(tǒng)學(xué)在面對(duì)部分種間形態(tài)相似或種內(nèi)變異較大的疑難物種時(shí),成為學(xué)者們選擇的重要技術(shù)手段,既能彌補(bǔ)傳統(tǒng)的不足,又能為植物的系統(tǒng)發(fā)育研究提供新的層面的數(shù)據(jù)支持。在傳統(tǒng)意義上,分子系統(tǒng)學(xué)可分為兩部分,即在種上水平的系統(tǒng)發(fā)生學(xué)(Phylogenetics)和種下水平的群體遺傳學(xué)(Population genetics),但隨著系統(tǒng)地理學(xué)、DNA barcoding理論的相繼出現(xiàn),系統(tǒng)發(fā)生學(xué)的體系也得到了迅速擴(kuò)大[37]。
2.1 系統(tǒng)發(fā)育研究
自21世紀(jì)以來(lái),老鴉瓣屬的新種相繼被發(fā)現(xiàn),由最初建屬的2個(gè)種(A.edulis和A.erythronioides)到目前的5個(gè)種:沈顯生[11]發(fā)現(xiàn)該屬的第3個(gè)物種皖郁金香(即安徽老鴉瓣);Tan[12]等發(fā)現(xiàn)該屬的第4個(gè)物種括蒼山老鴉瓣;Han[13]等發(fā)現(xiàn)第5個(gè)物種皖浙老鴉瓣。越來(lái)越多的新種被發(fā)現(xiàn),也為老鴉瓣屬的分類(lèi)研究提供了更多的材料。但是,相對(duì)于同科的其他的屬來(lái)說(shuō),如郁金香屬Tulipa[38]、百合屬Lilium[39]、豬牙花屬Erythronium[40]等,該屬的系統(tǒng)發(fā)育研究近乎空白,僅在對(duì)百合科其他類(lèi)群的系統(tǒng)發(fā)育研究中,對(duì)該屬少數(shù)種類(lèi)有過(guò)一些零星報(bào)道。
譚敦炎[4]等利用ITS和trnL-F序列對(duì)廣義郁金香屬的5個(gè)組進(jìn)行系統(tǒng)發(fā)育分析,其中sect.Tulipanum、sect.Leiostemones、sect.Eriostemone和sect.Orithyia聚為一支,而老鴉瓣屬Amana類(lèi)群(包括A.edulis、A.erythronioides、A.anhuiensis、A.kuocangshanica)單獨(dú)聚為一支,且獲得高度的支持(BP=99),并與豬牙花屬Erythronium形成并系,聚為一支(BP=100)。但研究結(jié)果只說(shuō)明了老鴉瓣屬和郁金香屬的物種位于不同的譜系分支,并沒(méi)有對(duì)老鴉瓣屬內(nèi)的系統(tǒng)發(fā)育關(guān)系進(jìn)行分析,且未涉及現(xiàn)今發(fā)現(xiàn)的所有老鴉瓣屬物種。
Allen[41]根據(jù)matK基因序列對(duì)Erythronium屬分析時(shí)發(fā)現(xiàn),老鴉瓣Amanaedulis與郁金香屬中的其他兩個(gè)種Tulipamontanavar. chrysantha和Tulipaturkestanica并未形成一支。同樣,JOHN[40]利用聯(lián)合片段(ITS,matKandrps16 introns)對(duì)Erythronium屬分析,也得到相同的結(jié)果,都表明Amana類(lèi)群獨(dú)立成一支。但不足的是,研究所用的物種只是包含了老鴉瓣屬中的一個(gè)或兩個(gè),而對(duì)于老鴉瓣屬整體的研究尚未見(jiàn)到報(bào)道。Christenhusz等[38]對(duì)郁金香屬進(jìn)行了分子系統(tǒng)學(xué)研究,其中老鴉瓣屬僅作為外群參與了系統(tǒng)發(fā)育樹(shù)的構(gòu)建,并沒(méi)有做進(jìn)一步的系統(tǒng)發(fā)育分析,且老鴉瓣屬僅包括老鴉瓣和二葉郁金香2個(gè)種。Nguyen等[42]以6個(gè)DNA分子片段為分子標(biāo)記,對(duì)百合目的Colchicaceae科進(jìn)行了系統(tǒng)發(fā)育分析,老鴉瓣同樣僅作為其中一個(gè)外群參加了構(gòu)樹(shù),并沒(méi)有分析和其他物種之間的系統(tǒng)發(fā)育關(guān)系。
本課題組前期也對(duì)老鴉瓣屬的5個(gè)種系統(tǒng)發(fā)育進(jìn)行初步研究(未發(fā)表),基于nrDNA和cpDNA聯(lián)合序列構(gòu)建系統(tǒng)發(fā)育樹(shù),結(jié)果顯示,5個(gè)來(lái)自老鴉瓣屬的種都各自形成單系分支。在系統(tǒng)發(fā)育樹(shù)中,老鴉瓣屬起先形成兩大分支,一個(gè)分支由老鴉瓣和括蒼山老鴉瓣組成;另一分支由二葉郁金香、安徽老鴉瓣和皖浙老鴉瓣組成,其中安徽老鴉瓣與皖浙老鴉瓣親緣關(guān)系更近,形成姐妹類(lèi)群,與二葉郁金香再形成并系。
2.2 生物地理學(xué)研究
生物地理學(xué)(Biogeography)是地理學(xué)和生物學(xué)的交叉學(xué)科,是研究生物分布及分布規(guī)律的學(xué)科,可分為歷史生物地理學(xué)和生態(tài)生物地理學(xué)。其中,生態(tài)生物地理學(xué)主要以在短時(shí)間跨度內(nèi)空間因子的作用,即自然因素的作用;而歷史生物地理學(xué)是在大的時(shí)間跨度內(nèi)(百萬(wàn)年),討論生物的進(jìn)化歷程[43]。近30年來(lái),隨著分子生物學(xué)的技術(shù)改革與進(jìn)步,促進(jìn)了歷史生物地理學(xué)的迅猛發(fā)展。1987年,Avise等在早期歷史生物學(xué)基礎(chǔ)上,建立了新的分支—譜系生物地理學(xué)(Phylogeography),主要通過(guò)對(duì)基因的譜系分析和單倍型的時(shí)空分布探討生物與古地質(zhì)事件(如冰川、地理隔離等)之間的關(guān)系,推測(cè)物種的起源、分化過(guò)程[44-46]。
Han[13]等基于核序列ITS和質(zhì)體序列trnL intron的部分DNA片段分析了老鴉瓣屬4個(gè)物種之間的系統(tǒng)發(fā)育關(guān)系,并證明皖浙老鴉瓣為有效種。然而,在分子系統(tǒng)發(fā)育分析中并未包括蒼山老鴉瓣,且每個(gè)種的樣本量較少,同時(shí)也沒(méi)有對(duì)該屬的屬內(nèi)分化時(shí)間和生物地理學(xué)進(jìn)行分析。根據(jù)不同物種間的系統(tǒng)發(fā)育關(guān)系以及各物種的地理分布,可以分析物種的生物地理格局,探討各物種形成現(xiàn)有地理格局的原因,能夠更好地理解物種宏進(jìn)化機(jī)制,同時(shí)也能對(duì)重要的地質(zhì)歷史事件進(jìn)行印證[47-51]。所以,有必要對(duì)老鴉瓣屬的分子系統(tǒng)發(fā)育學(xué)和生物地理學(xué)進(jìn)行深入的研究。
Vinnersten和Bremer[52]基于葉綠體rbcL序列對(duì)百合目主要分支分化時(shí)間和生物地理學(xué)進(jìn)行了研究,結(jié)果表明郁金香屬和豬牙花屬的分化時(shí)間在13±5.6 mya(million years ago;百萬(wàn)年前)。盡管沒(méi)有老鴉瓣屬的分化時(shí)間,但在后續(xù)研究中可以首先建立老鴉瓣屬與郁金香屬和豬牙花屬之間的系統(tǒng)發(fā)育關(guān)系,并利用建立的系統(tǒng)發(fā)育樹(shù),以郁金香屬和豬牙花屬的分化時(shí)間為時(shí)間標(biāo)定點(diǎn)計(jì)算老鴉瓣屬的分化時(shí)間和屬內(nèi)各物種之間的分化時(shí)間。由于老鴉瓣屬和豬牙花屬的親緣關(guān)系更近[4][39],可以推測(cè)老鴉瓣屬可能起源于中新世后期,情況是否如此,需要進(jìn)一步研究才能確定。
譚敦炎[4]等認(rèn)為中國(guó)的浙江和安徽是老鴉瓣屬植物的分布中心,Wu[8]等認(rèn)為老鴉瓣屬在中國(guó)、日本和朝鮮半島的隔離分布顯示其古老性。由于地質(zhì)史上較為嚴(yán)重的冰期會(huì)導(dǎo)致海平面下降,在此期間臺(tái)灣可能會(huì)和亞洲大陸通過(guò)陸地相連,而琉球群島南部則可能和臺(tái)灣東北部的陸地相連。這樣,在某一個(gè)冰盛期,亞洲大陸、臺(tái)灣和琉球群島可能是短暫相連的;當(dāng)冰期結(jié)束后,海平面上升,三者之間又會(huì)被海面隔離[53]。自從中新世這些島嶼形成以來(lái),短暫的陸地通道的形成和消失已經(jīng)重復(fù)了很多次。因此,越海擴(kuò)散技能很差的生物物種可能會(huì)通過(guò)以上冰盛期時(shí)短暫的陸地連接通道在亞洲大陸、臺(tái)灣和琉球群島之間擴(kuò)散[54-55]。而老鴉瓣屬的分化中心地理位置,以及該屬的隔離分布是否存在中國(guó)、朝鮮半島和日本之間在冰盛期的跨海擴(kuò)散,目前未見(jiàn)報(bào)道,需要在這方面做更多的研究。
3 結(jié)語(yǔ)
3.1 分類(lèi)問(wèn)題
目前,老鴉瓣屬Amana在百合科中分類(lèi)地位雖然得到了一些學(xué)者的承認(rèn),但對(duì)于老鴉瓣屬是另立還是歸入郁金香屬Tulipa的爭(zhēng)論仍然存在。隨著新種的相繼發(fā)現(xiàn),老鴉瓣屬已有5個(gè)物種,以我國(guó)的安徽分布最廣,包含4個(gè)種。由于物種分布的限制,目前,對(duì)于老鴉瓣屬的分子系統(tǒng)學(xué)研究相對(duì)較少,相關(guān)的研究也只是對(duì)該屬內(nèi)部分種,而針對(duì)老鴉瓣屬整體來(lái)說(shuō),并未見(jiàn)到報(bào)道。因此,本課題組對(duì)該屬5個(gè)植物種類(lèi),用4個(gè)DNA序列為分子標(biāo)記構(gòu)建老鴉瓣屬的系統(tǒng)發(fā)育樹(shù),確定系統(tǒng)樹(shù)中各個(gè)節(jié)點(diǎn)(祖先物種)的分化時(shí)間,同時(shí)確定各個(gè)節(jié)點(diǎn)的祖先物種可能的地理分布區(qū),結(jié)合各祖先物種當(dāng)時(shí)的古地質(zhì)條件和古氣候條件分析該屬的地理起源和物種擴(kuò)散路徑和方向,探討該屬的起源及屬內(nèi)分化原因。
3.2 資源及產(chǎn)業(yè)化
近年來(lái),市場(chǎng)對(duì)老鴉瓣的需求量日益增加,而野生自然資源正逐漸下降,供求失衡,矛盾日益尖銳。如何提高老鴉瓣的產(chǎn)量成為人們?cè)絹?lái)越關(guān)注的問(wèn)題。而人工栽培、組織培養(yǎng)、尋找新藥源等方法也成為當(dāng)前解決問(wèn)題的重要途徑。目前,對(duì)于老鴉瓣的生物學(xué)特性的研究已經(jīng)進(jìn)入成熟階段,不論在栽培和組織培養(yǎng)方面,都有研究報(bào)道。特別是在如何提高老鴉瓣產(chǎn)量的人工栽培上,其方法學(xué)也已成熟。此外,本課題組已對(duì)老鴉瓣屬的分子系統(tǒng)學(xué)展開(kāi)初步研究,并對(duì)于市場(chǎng)中常見(jiàn)的老鴉瓣偽品進(jìn)行分子鑒別的研究。所以,大量的基礎(chǔ)研究為老鴉瓣的栽培提供了數(shù)據(jù)支持,也為尋找新藥源及今后產(chǎn)業(yè)化的發(fā)展奠定基礎(chǔ)。
[1]謝宗萬(wàn).全國(guó)中草藥匯編(上冊(cè))[M].第2版.北京:人民衛(wèi)生出版社,1996,361.
[2]王惠民.老鴉瓣藥用歷史的變遷[J].中藥材,1992(6):35-36.
[3]Honda M.Amana, A New Genus of Liliaceae[J]. Bulletin of the Biogeographical Society of Japan, 1935(6): 19-21.
[4]譚敦炎,張震,李新蓉,等.老鴉瓣屬(百合科)的恢復(fù):以形態(tài)性狀的分支分析為依據(jù)[J].植物分類(lèi)學(xué)報(bào),2005(3):262-270.
[5] Hall A D, Beal J M. The Genus Tulipa.[J]. International Journal of Plant Sciences, 1940(1):143-146.
[6] Br.-Bl. Coloured Illustrations of Herbaceous Plants of Japan (Monocotyledoneae), by Kitamura S.; Murata G.; Koyama T.[J]. Vegetatio, 1966(5):289-289.
[7]Satake Y, et al.Wild Flowers of Japan (Herbaceous Plants) [M]. Tokyo: Heibonsha Ltd. Publishers, 1982.
[8]吳征鎰,路安民,湯彥承,等.中國(guó)被子植物科屬綜論[M].北京:科學(xué)出版社,2003.
[9]陳心啟,許介眉,梁樺筠,等.中國(guó)植物志:第14卷,被子植物門(mén)(單子葉植物綱),百合科(一)[M].北京:科學(xué)出版社,1980.
[10]梁松筠.百合科(狹義)植物的分布區(qū)對(duì)中國(guó)植物區(qū)系研究的意義[J].植物分類(lèi)學(xué)報(bào),1995(33):27-51.
[11]沈顯生.中國(guó)郁金香屬一新種[J].植物分類(lèi)與資源學(xué)報(bào),2001(23):39-40.
[12]Tan DY, Li XR, Hong DY.AmanaKuocangshanica(Liliaceae), a New Species from South-east China [J]. Botanical Journal of the Linnean Society, 2007(154): 435-442.
[13]Han BX, Zhang K, Huang LQ.AmanaWanzhensis (Liliaceae), a New Species from Anhui, China[J]. Phytotaxa, 2014(177):118-124.
[14]阮小麗,施大文.山慈菇的抗腫瘤及抑菌作用[J].中藥材,2009,32(12):1886-1888.
[15]熊宇,錢(qián)萍,黃萌,等.江西百合科一新記錄種——二葉郁金香[J].江西科學(xué),2012(30):317-318.
[16]譚敦炎,李新蓉,洪德元.安徽老鴉瓣新模式的指定與補(bǔ)充描述[J].西北植物學(xué)報(bào),2008(28):393-395.
[17]譚敦炎.中國(guó)郁金香屬(廣義)的系統(tǒng)學(xué)研究[D].北京:中國(guó)科學(xué)院(博士學(xué)位論文),2005.
[18]Tan D, Xinrong L I, Hong D. Amana Kuocangshanica (Liliaceae), A New Species from South-east China[J]. Botanical Journal of the Linnean Society, 2007, 154(3):435-442.
[19]Baker JG. Revision of the Genera and Species of Tulipeae[J]. Botanical Journal of the Linnean Society, 1874(14): 275-296.
[20]Sealy JR.Tulipaedulis[J] Curtis’s Botanical Magazine, 1957(171):293-293.
[21]Fiedler P L. Life History and Population Dynamics of Rare and Common Mariposa Lilies (Calochortus Pursh: Liliaceae)[J]. Journal of Ecology, 1988, 75(4):977-977.
[22]Mao ZM. Tulipa L. In: Flora Reipublicae Popularis Sinicae[M]. Beijing: Science Press, 1980.
[23]Nakaike T. New Flora of Japan[J]. Pteridophyta.rev. & Enlarg. 1992(1):417-418.
[24]梁松筠. 百合科(狹義)植物的分布區(qū)對(duì)中國(guó)植物區(qū)系研究的意義[J].植物分類(lèi)學(xué)報(bào),1995(33):27-51.
[25]Paul Wilkin. The Families and Genera of Vascular Plants. Volume III. Flowering Plants Monocotyledons Lilianae (Except Orchidaceae), by K. Kubitzki; H. Huber; P. J. Rudall; P. S. Stevens; T. Stützel[J]. Kew Bulletin, 1999, 54(4):1013-1013.
[26]宋向文,王德群,韓邦興.安徽省老鴉瓣屬藥用植物資源[J].中國(guó)野生植物資源,2014(33):51-52.
[27]吳正軍,朱再標(biāo),郭巧生,等.老鴉瓣傳粉生物學(xué)初步研究[J].中國(guó)中藥雜志,2012(37):293-297.
[28]梁麗,田霞,何云核.老鴉瓣光合生理特性研究[J].林業(yè)科技開(kāi)發(fā),2009(23):72-74.
[29]徐紅建,朱再標(biāo),郭巧生,等.光強(qiáng)對(duì)老鴉瓣生長(zhǎng)發(fā)育及光合特性的影響[J].中國(guó)中藥雜志,2012(37):442-446.
[30]楊穎,朱再標(biāo),郭巧生,等.冷藏對(duì)老鴉瓣鱗莖休眠解除及栽后生長(zhǎng)情況的影響[J].中國(guó)中藥雜志,2015(40):48-52.
[31]繆媛媛,朱再標(biāo),郭巧生,等.摘蕾和人工授粉對(duì)老鴉瓣生長(zhǎng)和產(chǎn)量的影響[J].中國(guó)中藥雜志,2014(39):2016-2018.
[32]楊小花,繆媛媛,郭巧生,等.不同等級(jí)老鴉瓣生長(zhǎng)繁殖特征研究[J].中草藥,2015(46):3746-3750.
[33]楊小花,郭巧生,朱再標(biāo),等.不同采收期老鴉瓣生物量積累及藥材品質(zhì)研究[J].中國(guó)中藥雜志,2016(41):624-629.
[34]朱麗芳,史俊,朱再標(biāo),等.老鴉瓣芽莖組織培養(yǎng)初步研究[J].中草藥,2014(45):563-568.
[35]朱麗芳,徐超,朱再標(biāo),等.TDZ和NAA對(duì)老鴉瓣不定芽誘導(dǎo)和叢生芽增殖的影響[J].中國(guó)中藥雜志,2014(39):3030-3035.
[36]Suárez Díaz E, Anaya Muoz VH. History, Objectivity, and the Construction of Molecular Phylogenies[J]. Stud Hist Phil Biol Biomed Sci, 2008(39): 451-68.
[37]徐宏發(fā),王靜波.分子系統(tǒng)學(xué)研究進(jìn)展[J].生態(tài)學(xué)雜志,2001(20):41-46.
[38]Christenhusz M J M, Govaerts R, David J C, et al. Tiptoe through the Tulips-cultural History, Molecular Phylogenetics and Classification of Tulipa (Liliaceae)[J]. Botanical Journal of the Linnean Society, 2013, 172(3):280-328.
[39]R?nsted N, Law S, Thornton H, et al.. Molecular Phylogenetic Evidence for the Monophyly of Fritillaria and Lilium (Liliaceae; Liliales) and the Infrageneric Classification of Fritillaria[J]. Molecular Phylogenetics & Evolution, 2005, 35(3):509-527.
[40]JOHN CB, CLENNET, et al. Phylogenetic Systematics of Erythronium (Liliaceae):Morphological and Molecular Analyses[J]. Botanical Journal of the Linnean Society, 2012(170): 504-528.
[41]Allen GA, Soltis DE, Pamela S. Soltis. Phylogeny and Biogeography of Erythroniutn (Liliaceae) Inferred from Chloroplast matK and Nuclear rDNA ITS Sequences[J]. Systematic Botany, 2003(28): 512-523.
[42]Nguyen TPA, Kim JS, Kim JH. Molecular Phylogenetic Relationships and Implications for the Circumscription of Colchicaceae (Liliales)[J]. Botanical Journal of the Linnean Society, 2013(172): 255-269.
[43]趙鐵橋.歷史生物地理學(xué)進(jìn)展[J].昆蟲(chóng)分類(lèi)學(xué)報(bào),1992(1):35-48.
[44] Avise JC, Arnold J, Ball RM, et al.. Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics[J]. Ecology, Evolution, and Systematics, 1987(18): 489-522.
[45]Reem E, Douek J, Paz G, et al.Phylogenetics, Biogeography and Population Genetics of the Ascidian Botryllus Schlosseri, in the Mediterranean Sea and beyond[J].Molecular Phylogenetics & Evolution, 2017(107):221-231.
[46]Cardoso D, Harris D J, Wieringa J J, et al.. A Molecular-dated Phylogeny and Biogeography of the Monotypic Legume Genus Haplormosia, a Missing African Branch of the otherwise American-Australian Brongniartieae Clade[J].Molecular Phylogenetics & Evolution, 2017(107):431-442.
[47]Sakaguchi S, Qiu Y X, Liu Y H, et al. Climate Oscillation during the Quaternary Associated with Landscape Heterogeneity Promoted Allopatric Lineage Divergence of a Temperate Tree Kalopanax Septemlobus, (Araliaceae) in East Asia[J]. Molecular Ecology, 2012, 21(15):3823-3823.
[48]Wang W, Lin L, Xiang X G, et al.. The Rise of Angiosperm-dominated Herbaceous Floras: Insights from Ranunculaceae[J]. Scientific Reports, 2016(6):27259-27259.
[49]Koch R A, Wilson A W, Séné O, et al.. Resolved Phylogeny and Biogeography of the Root Pathogen Armillaria and its Gasteroid Relative, Guyanagaster[J]. Bmc Evolutionary Biology, 2017,17(1):33-33.
[50]Chen LY, Chen JM, Gituru RW, et al. Eurasian Origin of Alismatidae Inferred from Statistical Dispersal-vicariance Analysis[J]. Molecular Phylogenetics and Evolution, 2013(67): 38-42.
[51]Burke JM, Bayly MJ, Adams PB, et al. Molecular Phylogenetic Analysis ofDendrobium(Orchidaceae), with Emphasis on the Australian SectionDendrocoryne, and Implications for Generic Classification[J]. Australian Systematic Botany, 2008(21): 1-14.
[52]Vinnersten A, Bremer K. Age and Biogeography of Major Clades in Liliales[J]. American Journal of America, 2001(88): 1695-1703.
[53] Br.-Bl. Coloured Illustrations of Herbaceous Plants of Japan (Monocotyledoneae), by Kitamura S.; Murata G.; Koyama T.[J]. Vegetatio, 1966(5):289.
[54]Toda M, Nishida M, Matsui M, et al. Allozyme Variation Among East Asian Populations of the Indian Rice Frog Rana Limnocharis, (Amphibia: Anura)[J]. Biochemical Systematics & Ecology, 1997, 25(2):143-159.
[55]Lin SM, Chen CA, Lue KY. Molecular Phylogeny and Biogeography of the Grass Lizards GenusTakydromus(Reptilia: Lacertidae) of East Asia[J]. Molecular Phylogenetics and Evolution, 2002(22): 276-288.
ResearchProgressofPlantResourcesofAmanaHondaandItsMolecularSystematics
CHEN Shaotong1,2, LIU Feng1,2, SONG Xiangwen2,3, ZHAO Qun2,3, HAN Bangxing2,3
( 1.AnhuiUniversityofChineseMedicine,Hefei230012,China;2.WestAnhuiUniversity,Lu’an237012,China;3.SynergeticInnovationCenterofDendrobiaIndustrializationDevelopmentinAnhuiProvince,Lu’an237012,China)
The genus Amana Honda belongs to the tribe Lilieae (Liliales: Liliaceae) and it is a group confined to East Asia. At present, there are relatively few studies on the Amana Honda, and more concentrated on the chemical composition, tissue culture and biological characteristics of Amana edulis (Miq.) Honda, and few reports on other species in this genus. In this paper, we reviewed the advances in the phylogenetic position, the taxonomic status of the genus, biological characteristics and molecular systematics of the genus. Based on the existing research and the preliminary research the group, and the key scientific problems existing in this genus, it is of great significance for the development, utilization and protection of Amana Honda medicinal plant resources.
Amana edulis (Miq.) Honda; Amana Honda; plant resources; biological characteristics; molecular systematics
R284
A
1009-9735(2017)05-0102-07
2017-09-12
安徽省自然科學(xué)基金(1508085MH203);中央本級(jí)重大增減支項(xiàng)目(2060302);中央財(cái)政林業(yè)科技推廣示范項(xiàng)目(2016TG05);安徽高校自然科學(xué)研究項(xiàng)目(KJ2016SD61);現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系建設(shè)專項(xiàng)(CARS-21)。
陳紹通(1992-),女,河南原陽(yáng)人,碩士研究生,研究方向:中藥資源開(kāi)發(fā)與利用;通信作者:韓邦興(1978-),男,安徽樅陽(yáng)人,碩士生導(dǎo)師,教授,研究方向:中藥資源。
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車(chē)觀察(2018年10期)2018-11-06 07:05:26
