芝麻繁育特性研究Ⅳ:海南南繁芝麻的天然異交率
2017-11-07 07:48:06顏廷獻樂美旺饒月亮顏小文周紅英
江西農業學報 2017年11期
孫 建,顏廷獻,樂美旺,饒月亮,顏小文,周紅英
(江西省農業科學院 作物研究所/國家油料改良中心 南昌分中心/江西省油料作物生物學重點實驗室,江西 南昌 330200)
芝麻繁育特性研究Ⅳ:海南南繁芝麻的天然異交率
孫 建,顏廷獻,樂美旺*,饒月亮,顏小文,周紅英
(江西省農業科學院 作物研究所/國家油料改良中心 南昌分中心/江西省油料作物生物學重點實驗室,江西 南昌 330200)
利用顯/隱性單基因控制的單花/三花性狀為標記,對海南南繁芝麻的天然異交率進行了測定。結果顯示:鄰近異花粉源的芝麻的天然異交率為4.44%~4.76%;天然異交率與異花粉源距離存在極顯著的負相關(r= -0.626**),隨著距離的增大,天然異交率不斷下降,大約在距離異花粉源15~20行(6~8 m)處未檢測到天然異交率;不同種植方向的天然異交率存在一定的差異。
芝麻;繁育特性;天然異交率;南繁;海南
“南繁”作為我國首創的一種獨特育種及繁育方式,是利用我國南方特別是海南島南部地區冬春季節氣候溫暖的優勢條件,將夏季在北方種植的農作物育種材料,于冬春季節在南方再種植一季或兩季的農作物育種方式,在農作物新品種培育過程中發揮著縮短育種年限、加速世代繁育的特殊作用[1-3]。在南繁作物的加代、繁育與種子生產中,天然異交率的存在不僅是作物種子純度降低、種性退化的重要因素,也是基因漂流引起環境和食品安全的潛在風險[4]。研究和明確作物南繁過程中天然異交率的大小,對于該作物南繁過程中的繁育特性和種子生產研究具有重要意義。
芝麻是海南傳統的油料作物,種植歷史悠久,明代已有記載。海南芝麻生產遍及全島各縣市,主要產區集中在瓊山、海口、澄邁等地,近年來種植面積約3300 hm2,歷史最高達13333 hm2[5]。隨著科研工作的不斷加快與深入,南繁在芝麻育種和群體構建等方面具有越來越重要的作用,越來越多的芝麻品種有著南繁的經歷,如駐芝10號、漯12和鄭芝14號等[6-8]。芝麻是具有鮮艷花冠、泌蜜量較大的開放式開花習性的主要蜜源作物之一[9-10],是蜜蜂等傳粉昆蟲的重要訪問對象。因此,在芝麻南繁中,天然異交率是一個不容忽視的客觀問題。已有的報道顯示,國外芝麻天然異交率在1%~65%,不同地區、不同品種差異較大[11-16]。筆者團隊利用多組相對性狀在我國主要芝麻產區開展了異交率測定研究,發現芝麻天然異交率在5.13%~23.35%[16],同時確定蜜蜂、切葉蜂等昆蟲是芝麻傳粉,特別是異花傳粉的主要媒介[17]。但是,關于芝麻南繁過程中的天然異交率的研究報道尚屬空白,因此,研究并明確芝麻南繁過程中的天然異交率對芝麻南繁中的群體構建、良種繁育和種子生產等工作具有十分重要的理論和實際意義,同時也對當地芝麻生產具有一定的參考價值。
1 材料與方法
1.1 試驗材料
以花期基本一致的顯/隱性單基因控制的單花/三花性狀為標記開展異交率測定,單花芝麻品種為“金黃麻”,三花芝麻品種為“贛芝9號”。試驗材料均通過多年網帳隔離純化而來。
1.2 試驗原理及設計
1.2.1 試驗原理 利用受顯/隱性單基因控制的單花/三花性狀為標記進行天然異交率測定,以顯性單基因控制的單花性狀材料為異花粉源,將單花和三花材料按試驗設計種植,收獲三花植株上的種子進行點播(不間苗),鑒定其標記性狀,出現的單花植株即為天然異交產生,統計其出現頻率以計算天然異交率。
1.2.2 試驗設計與異交率計算 試驗在海南省三亞市天涯區江西省農業科學院南繁試驗基地進行。采用條播種植模式,在周邊無其他芝麻花粉源存在的前提下,在寬2 m、長25 m左右的東西走向的廂面上,先在中心位置種植5行異花粉源(單花)材料,在其他位置雙向條播對應的三花性狀材料,行距40 cm,田間管理如常。成熟后按行分別收獲三花性狀材料植株上的種子,每行隨機收獲2株,即重復2次。將收獲種子在江西省農科院試驗基地(南昌蓮塘)進行點播(不間苗)種植,并在花期進行性狀調查,測定距離異花粉源不同方向、不同行距下芝麻的天然異交率。以單花性狀為目標性狀,調查和計算天然異交群體內該目標性狀植株占總群體植株的百分率,即為天然異交率(%)。試驗數據整理與相關分析通過Excel和SPSS軟件進行。
2 結果與分析
2.1 不同方向、不同距離天然異交率的變化與相關分析
檢測結果(表1)顯示:東面株行的天然異交率為0%~4.44%,西面株行的天然異交率為0%~4.76%;在與異源花粉相鄰的3行中,東面株行的天然異交率為3.11%~4.44%,西面株行的天然異交率為0.76%~4.76%。圖1、圖2顯示,隨著距離異花粉源的行數的增加,東、西面株行的天然異交率呈波動下降趨勢,其中東面株行在20行(8.0 m)以后沒有檢測到天然異交率,西面株行在10行(4.0 m)以后沒有檢測到天然異交率。此外,東面株行的天然異交率整體上要高于西面株行的,而西面株行的天然異交率降低幅度明顯大于東面株行的。
相關分析結果表明,芝麻的天然異交率與異花粉源距離呈極顯著(P<0.01)負相關,相關系數為-0.626。其中,東面株行的天然異交率與異花粉源距離呈極顯著負相關(r=-0.774**),西面株行的天然異交率與異花粉源距離呈顯著負相關(r=-0.484*)。
可見,南繁芝麻存在一定程度的天然異交率,鄰近異花粉源芝麻的天然異交率可達4.76%;天然異交率與異花粉源的距離有直接關系,隨著相隔行數的增多,天然異交率逐漸減少。不同種植方向芝麻的天然異交率差異明顯,這可能與風向影響昆蟲訪問芝麻花有關。
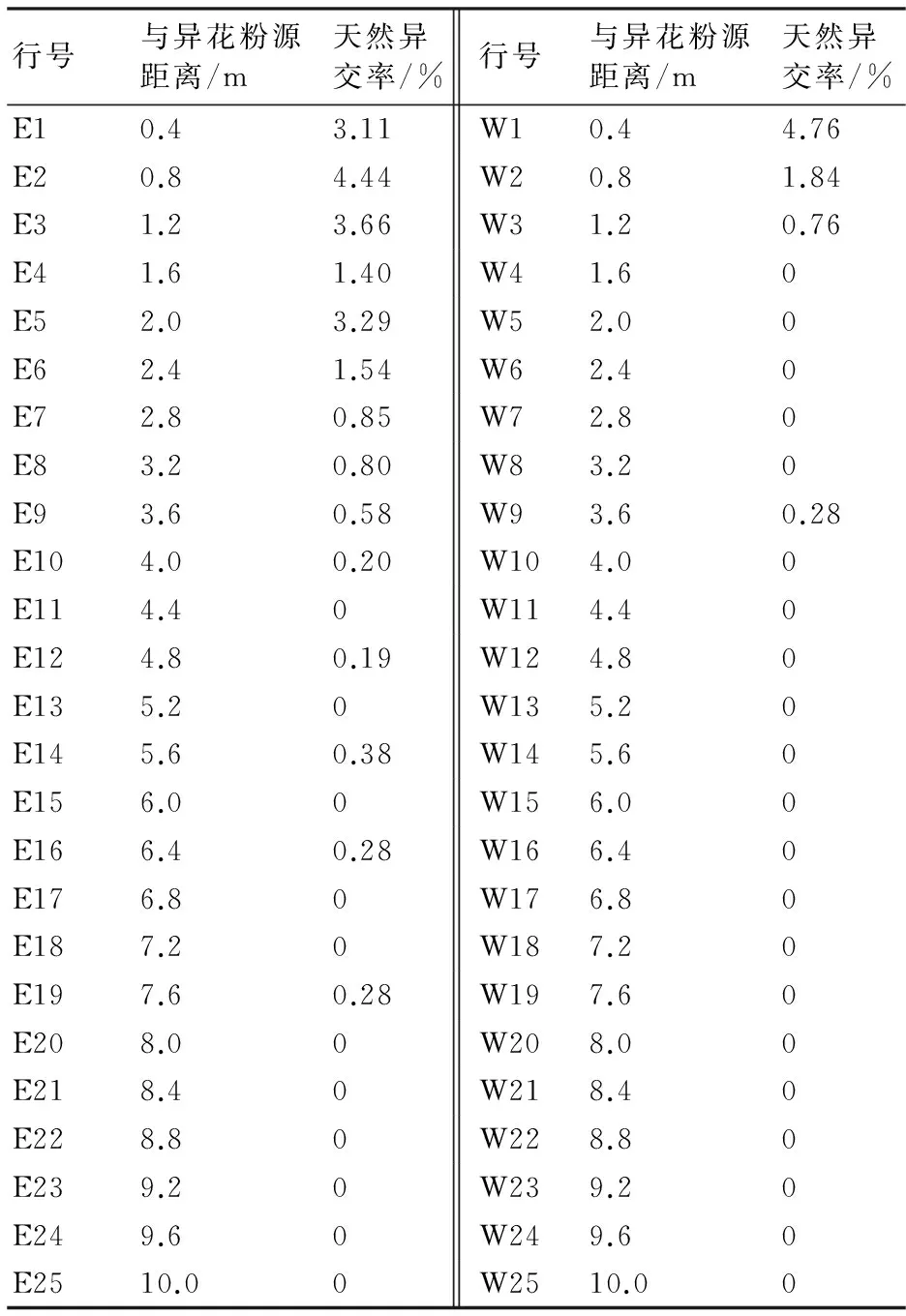
表1 不同方向、不同距離芝麻的天然異交率
注:E為東面株行,W為西面株行。
2.2 零異交率的理論距離估算
分別對表1數據和圖1、圖2進行趨勢線(對數)分析,結果表明:東面株行芝麻天然異交率的趨勢線公式為y=-1.4069ln(x)+4.1044,在距離異花粉源約7.6 m(19行)時天然異交率為0;西面株行芝麻天然異交率的趨勢線公式為y=-0.8986ln(x)+2.3908,在距離異花粉源約6.0 m(15行)時天然異交率為0。該結果較其他生態區的估算值[16]略小,可能與試驗基地(種植作物以水稻為主)的昆蟲種類和數量等環境因素有關。因此,在芝麻海南南繁過程中,不同材料間其相鄰15~20行之內(6~8 m)所獲種子存在雜合的可能性較高,這為芝麻南繁工作提供了理論參考。
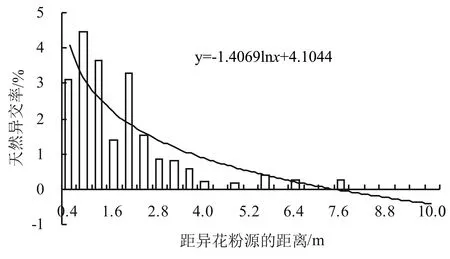
圖1 東面株行不同距離芝麻的天然異交率

圖2 西面株行不同距離芝麻的天然異交率
3 討論
一直以來,芝麻都被認為是自花授粉作物,但在科研和生產實踐中,較高頻率出現的天然異交現象一直困擾著芝麻繁育與種子生產。不同國家和地區的研究者也對芝麻的天然異交率先后進行了報道,例如:Langham檢測到委內瑞拉的芝麻存在4.6%的天然異花授粉作用;Rheenen發現尼日利亞芝麻的天然異交率為3.5%~9.6%;Khidir估算的蘇丹芝麻的天然異交率為3.1%~6.7%; Musa等研究結果顯示墨西哥芝麻的異交率為4.4%~24.8%[12-15];孫建等[16]首次通過多年多點試驗對我國芝麻主要產區的異交率進行了測定,結果顯示混合種植模式下芝麻的天然異交率為5.13%~23.35%,平均12.69%,在不同生態區、不同年份、不同品種間均存在較大的差異;Yermanos更是認為在特定條件下,芝麻的天然異交率可以高達65%[13]。本試驗檢測到芝麻在海南南繁過程中,緊鄰異花粉源的天然異交率為4.44%~4.76%,該結果與前人的研究結果[12-16]相近,說明了芝麻天然異交率的普遍存在。但該結果較我國主產區的芝麻天然異交率[16]稍低,可能與昆蟲數量和環境因素存在一定的關系。首先,由于風媒對芝麻傳粉基本上不起作用,所以昆蟲是芝麻主要的傳粉媒介[17];冬季海南三亞氣溫不高且常受大陸冷空氣的影響,傳粉昆蟲的數量和活動頻率并不太高,直接導致天然異交率的降低。其次,海南是我國冬季蔬菜的主要生產地,在本試驗實施地的周邊地區種植有大量的豇豆、黃秋葵及瓜果類等蔬菜,而緊鄰試驗地核心區則種植了大量的水稻,農藥的使用和蔬菜花粉對昆蟲的吸引,直接導致芝麻訪花昆蟲的減少,從而使得天然異交率稍低于主產區。
本試驗結果顯示,東西不同方向的芝麻的異交率存在較明顯的差距,東面芝麻的天然異交率隨著與異源花粉距離的增加而逐步下降,在距離異花粉源8 m之后沒有檢測到天然異交率,而西面芝麻的天然異交率則隨著與異源花粉距離的增加而陡然下降,在距離異花粉源4 m之后便沒有檢測到天然異交率。這種因種植方向帶來的天然異交率差異可能與三亞氣候條件和昆蟲的生活習性相關,冬春季節三亞市天涯區受氣候和山體的影響,試驗基地以東北風為主。而蜜蜂等大多數訪花昆蟲均為晝出性昆蟲,其具有對日光的正趨性,同時,在風速較低時,昆蟲有逆風飛行擴散的習性,在訪花時,下風處的昆蟲有靠風傳氣味尋找蜜源的特點[18-19]。訪花昆蟲的這些特性和試驗地的風向,直接導致訪花昆蟲將異源花粉優先傳送到東面植株,使東面芝麻的天然異交率要普遍高于西面的。因此,訪花昆蟲的訪花習性影響了芝麻天然異交率的高低,日照方向和花期風向在一定程度上可以作為良種繁育和種子生產的考慮因素。
農作物良種繁育對種子純度有嚴格的要求,尤其對原原種純度,一般要求在繁育過程中排除機械混雜等人為因素之外,其天然異交率應該控制在0.1%以內[20]。本試驗檢測到海南南繁芝麻天然異交率達到4.44%~4.76%,因此通過有效隔離降低或排除天然異交率在芝麻南繁中顯得較為重要。距離隔離是芝麻良種繁育過程中主要方式之一[11]。孫建等[16]在多年多點監測我國主產區芝麻天然異交率的基礎上,提出了400 m空間距離加8~12 m保護行的理論隔離方法。本試驗在條播種植模式下,綜合東西方向異交率出現的特點,估算出距離異源花粉6~8 m的情況下,南繁芝麻的天然異交率幾乎為0。參考油菜、棉花等其他作物在歐洲、加拿大和澳大利亞的相關規定[21-25],我們提出,芝麻南繁中理論隔離距離為400 m,加上6~8 m的保護行,這樣可滿足良種繁育種99.9%的純度要求,但其實際隔離效果則需要進一步試驗驗證。
[1] 佟屏亞.異地培育理論的創立與南繁發展[J].中國種業,2000,28(2):11-13.
[2] 董照輝,張應祿,劉繼芳,等.我國南繁基地建設問題的探討與建議[J].中國農業科技導報,2010(1):52-55.
[3] 周晶,羅勁梅,徐雪榮,等.南繁基地建設的戰略意義及發展方向[J].熱帶農業工程,2014,38(3):45-48.
[4] 胡凝,姚克敏,袁潛華,等.海南南繁區水稻基因飄流的最大閾值距離及其時空分布特征[J].中國農業科學,2014,47(23):4551-4562.
[5] 符齋,云白楊.海南省油料作物生產存在的問題與發展對策[J].中國農技推廣,2010,26(2):10-12.
[6] 張仙美.芝麻新品種漯12的選育及栽培要點[J].河南農業科學,2002(7):18-19.
[7] 崔向華,段瑩,闞躍峰,等.黑芝麻新品種駐芝10號的選育[J].中國種業,2008(7):60-61.
[8] 張體德,張海洋,衛雙玲,等.芝麻新品種鄭芝14號的選育及應用[J].農業科技通訊,2010(12):178-180.
[9] 劉炳侖.我國春夏秋冬44種主要蜜源植物的花粉形態[J].養蜂科技,2001(4):4-6.
[10] 于先覺,韓開菊,李莉.芝麻花期的蜂群管理[J].中國蜂業,2009,60(6):21.
[11] Weiss E A. Castor, sesame and safflower [M]. London: Leonard Hill, 1971: 311-525.
[12] Brar G S, Ahuja K L. Sesame, its culture, genetics, breeding and biochemistry: annual review [J]. Plant Science, 1979(1):245-313.
[13] Yermanos D M. Sesame [M]//Fehr W, Hadleys H. Hybridization of crop plants. Madison: Agronomy-crop Science Society of America, 1980: 549-563.
[14] 馮祥運,張秀榮,劉越英.基因庫芝麻種子繁殖更新方法研究[J].中國油料,1997,19(1):63-65.
[15] 翟志席,詹英賢.雜交芝麻[M].北京:中國農業出版社,1997:147-150.
[16] 孫建,顏廷獻,高德學,等.芝麻繁育特性研究Ⅰ:天然異交率的測定[J].中國油料作物學報,2015,37(4):462-466.
[17] 孫建,魏星,樂美旺,等.芝麻繁育特性研究Ⅱ:異花傳粉媒介的確定[J].作物雜志,2015(3):36-40.
[18] 王立和,張青文,楊淑霞,等.螟黃赤眼蜂在棉田的擴散方向及其與風速風向的關系[J].昆蟲學報,1998,41(增刊):76-81.
[19] 張青文,張巍巍,蔡青年,等.苜蓿切葉蜂授粉擴散行為及苜蓿種子增產效應的研究[J].應用生態學報,1999,10(5):606-608.
[20] 王志興,王旭靜,賈士榮.主要農作物轉基因飄流頻率和距離的數據調研與分析Ⅰ:背景、調研目的及所考慮的問題[J].中國農業科技導報,2011,13(3):26-29.
[21] 李允靜,盧長明,王旭靜,等.主要農作物轉基因飄流頻率和距離的數據調研與分析Ⅴ:油菜[J].中國農業科技導報,2012,14(1):49-56.
[22] 王旭靜,賈士榮,王志興.主要農作物轉基因飄流頻率和距離的數據調研與分析Ⅵ:棉花[J].中國農業科技導報,2012,14(6):19-22.
[23] Bilsborrow P E, Evans E J, Bowman J, et al. Contamination of edible double-low oilseed rape crops via pollen transfer from high erucic cultivars [J]. J Sci Food Agne, 1998, 76: 17-22.
[24] SCIMAC. Guidelines for growing newly developed herbicide tolerant crops [Z/OL]. http://www.scimac.org.uk/files/onfarm_guidelines.pdf, 1999.
[25] Staniland B K, McVetty P B E, Friesen L F, et al. Effectiveness of border areas in confining the spread of transgenicBrassicanapuspollen [J]. Can J Plant Sci, 2000, 80: 521-526.
StudyonBreedingCharacteristicsofSesame(Sesamumindicum) Ⅳ:SpontaneousOutcrossingRateofSesameinWinterMultiplicationinHainan
SUN Jian, YAN Ting-xian, YUE Mei-wang*, RAO Yue-liang, YAN Xiao-wen, ZHOU Hong-ying
(Crop Research Institute, Jiangxi Academy of Agricultural Sciences / Nanchang Branch, National Center of Oil Crop Improvement / Jiangxi Provincial Key Laboratory of Oil Crop Biology, Nanchang 330200, China)
Using the mono-floral / tri-floral character controlled by dominant / recessive single gene as marker, we measured the spontaneous outcrossing rate (SOR) of sesame (Sesamumindicum) in winter multiplication in Hainan. The results showed that: the SOR of sesame adjacent to different pollen sources was 4.44%~4.76%, and it was very significantly negatively correlated with the distance from different pollen source (r=-0.626**); the SOR of sesame gradually decreased with the increase in the distance from different pollen source, and it could not be detected beyond 15~20 lines (6~8 m) from different pollen sources; there was a certain difference in SOR among sesame plants planted in different directions.
Sesame; Breeding characteristics; Spontaneous outcrossing rate; Winter multiplication; Hainan
2017-07-20
國家特種油料產業技術體系建設項目(CARS-14)。
孫建(1981─),男,江西高安人,副研究員,主要從事芝麻遺傳育種與良種繁育研究。*通訊作者:樂美旺。
S565.3
A
1001-8581(2017)11-0017-04
(責任編輯:黃榮華)
