mmu-miR-3475-3P在心臟發育中的生物信息學分析*
2017-10-12 10:21:18周揮銘劉玲娟
重慶醫學 2017年26期
沈 興,潘 博,周揮銘,劉玲娟,田 杰
(1.重慶醫科大學附屬兒童醫院心內科,重慶 400014;2.西南醫科大學附屬醫院兒二科,四川瀘州 646000)
·生物信息學·
mmu-miR-3475-3P在心臟發育中的生物信息學分析*
沈 興1,2,潘 博1,周揮銘1,劉玲娟1,田 杰1
(1.重慶醫科大學附屬兒童醫院心內科,重慶 400014;2.西南醫科大學附屬醫院兒二科,四川瀘州 646000)
目的探討mmu--miR-3475-3P可能參與的生物學過程及信號通路。方法用實時熒光定量PCR方法測定mmu-miR-3475-3P在小鼠胚胎心臟和成熟心臟中的表達。再綜合應用TargetScan、miRDB、miRanda等常用microRNA在線數據庫對miR3475-3p進行靶基因預測,對所得靶基因進行基因功能注釋(Go)和信號通路分析。結果mmu-miR-3475-3P在小鼠胚胎心臟和成熟心臟中存在明顯的表達差異,運用TargetScan、miRDB、miRanda對miR3475-3P進行生物信息學分析發現該microRNA可能調控441個靶基因。結論mmu-miR-3475-3P在小鼠胚胎心臟高表達,其預測的靶基因富集于多個信號通路及細胞生物學過程。
mmu-miR-3475-3P;生物信息學;靶基因;心臟發育
目前,心臟疾病的發病率逐年上升,已成為威脅人類健康的主要疾病之一[1]。對正常心臟發育過程的深入解析可為實現對心臟疾病的早期防治提供理論依據。心臟的構建涉及多細胞、多基因,也受到遺傳與環境因素的共同影響。表觀遺傳作為遺傳與環境因素之間的橋梁在胚胎心臟發育過程中具有重要作用,特別是表觀遺傳中的microRNA被認為廣泛參與了心臟的發育及疾病的發生、發展[2-5]。mmu-miR-3475-3P是本課題組在前期研究中發現的在胎鼠心肌中高表達的microRNA,目前對其的研究較少。本研究擬通過生物信息學分析,預測mmu-miR-3475-3P的關鍵靶基因和涉及的信號通道,為進一步行mmu-miR-3475-3P在心臟發育中的功能、機制研究奠定基礎。
1 材料與方法
1.1材料 實驗動物采用清潔級昆明孕鼠,購于重慶醫科大學實驗動物中心[合格證號:SYXK(渝)2007-0016]。實驗用引物均購自廣州市銳博生物科技有限公司,RNA酶抑制劑購自美國Thermo Scientific公司,脫氧核糖核苷三磷酸購自Tiangen公司,綠色熒光染料購自美國Bio-Rad公司。
1.2方法
1.2.1mmu-miR-3475-3P表達量在胚胎鼠和成年鼠心臟中的檢測 以胚胎14.5 d的昆明小鼠(n=3)和生后21 d的成年昆明小鼠(n=3)為對象,用Trizol試劑提取總RNA,并利用紫外分光光度計K5500在OD260/280,OD260/230波長下進行RNA質檢,以該RNA作為模板,以1 μg總RNA反轉錄成CDNA第1鏈,并以其為模板進行PCR擴增,根據小鼠mmu-miR-3475-3P(MIMAT0015219)的編碼序列5′-UCU GGA GGC ACA UGG UUU GAA-3′設計合成PCR引物,按如下條件進行PCR反應:95 ℃ 20 s、95 ℃ 10 s、60 ℃ 30 s、70 ℃ 1 s,共40個循環,繪制溶解曲線。以U6作為反應內參,microRNA的含量通過2-△△ct法定量計算。
1.2.2mmu-miR-3475-3P生物信息學分析 綜合應用TargetScan、miRDB、miRanda等常用microRNA在線數據庫對mmu-miR3475-3p進行靶基因預測。對所得靶基因進行基因功能注釋(gene ontology,Go)(http://www.geneontology.org/),并建立起統計模型,分析整個基因組中最顯著的功能,計算這些基因在某(多)個特定的分支的超幾何關系,根據基因的GO 注釋,選擇本物種的所有基因作為背景基因,用超幾何分布計算P值,以P<0.05為顯著性閾值分別得到相對于背景具有統計意義的高頻率注釋,從而得到基因集合在GO類別上的分布信息和顯著性情況。將mmu-miR3475-3p的預測靶基因進行基于Kyoto encyclopedia of genes and genomes(KEGG,http://www.genome.jp/)的生物學通路數據分析,通過Fisher Exact Test計算P值,以P<0.05為顯著性閾值得到基因集合相對于背景具有統計意義的信號轉導及疾病通路。
“喂!放我出去!”步凡用力地敲擊著門板,但回應他的,只有門外的嬉笑聲:“黃皮仔,你就在這里好好待著吧!明天早上掃地的人會來營救你的!”
2 結 果
人們根據第二摩崖石刻“永樂十八年領軍至此”“洪熙元年領軍至此”“宣德七年領軍至此”,普遍認為劉清共三次來到船廠。

圖1 mmu--miR-3475-3P靶基因文恩交集圖
2.2mmu-miR-3475-3P生物信息學分析 運用TargetScan、miRDB、miRanda對mmu-miR-3475-3P進行靶基因預測分別獲得3 797、518、8 431個靶基因,設3個數據庫中共同預測的靶基因為候選基因,共441個(圖1)。針對靶基因的GO分析提示mmu-miR-3475-3P涉及多種生物學過程(圖2),其中多個基因涉及心臟、血管發育、跨膜離子通道轉運等(表1)。利用KEGG經典生物通路數據庫對mmu-miR-3475-3P的靶基因進行信號通路富集分析,結果提示441個靶基因參與廣泛的信號轉導通路,包括代謝通路、內吞作用、Wnt信號通路、Hippo信號通路等,而其中與心血管發育密切相關的Wnt信號通路共涉及15個基因,Hippo信號通路共涉及8個基因,Notch信號通路共涉及2個基因,見表2。
2.1mmu-miR-3475-3P在胚胎小鼠和成年小鼠心肌中的表達量分析 mmu-miR-3475-3P在胎鼠14.5 d的心肌中的表達量明顯高于成年鼠心肌中的表達量,在胎鼠中的表達量為17.41±5.74,在成年鼠心肌中的表達量為1.00±0.54,二者比較差異有統計學意義(P<0.05)。
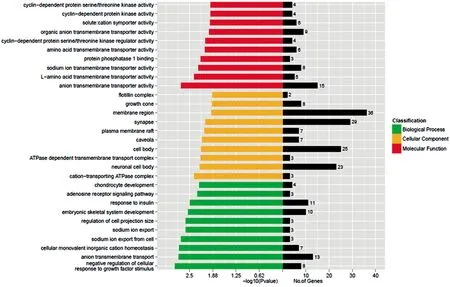
圖2 mmu--miR-3475-3P靶基因生物學GO分析
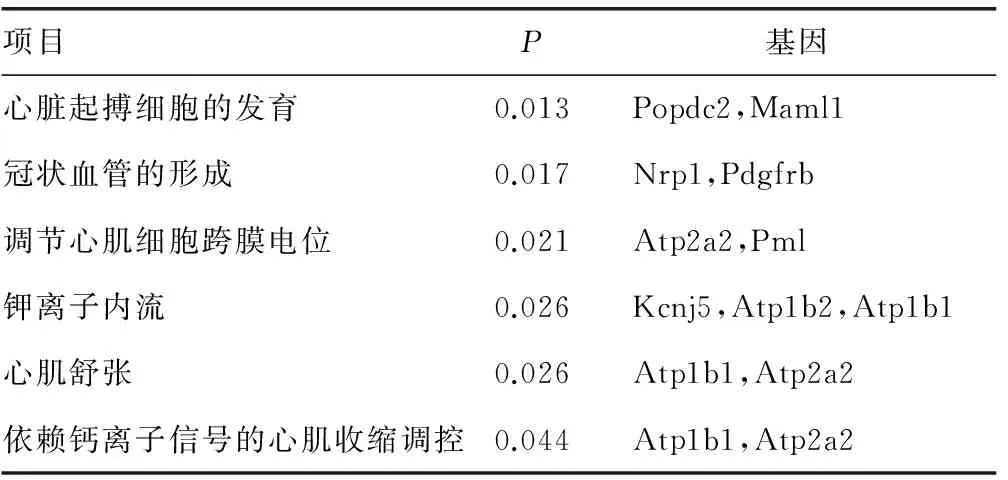
項目P基因心臟起搏細胞的發育0.013Popdc2,Maml1冠狀血管的形成0.017Nrp1,Pdgfrb調節心肌細胞跨膜電位0.021Atp2a2,Pml鉀離子內流0.026Kcnj5,Atp1b2,Atp1b1心肌舒張0.026Atp1b1,Atp2a2依賴鈣離子信號的心肌收縮調控0.044Atp1b1,Atp2a2

表2 與心血管發育密切相關的KEGG經典生物通路示例
3 討 論
microRNA是一類小分子非編碼的RNA分子,起到對基因表達的負性調控作用,廣泛參與細胞的分化、遷徙,組織器官的形成等。生物信息學預測microRNA可能控制著約70%的生理過程,但只有小部分microRNA的靶基因及功能得到了初步闡述,大部分microRNA的功能仍然未知。研究未知microRNA對組織器官的發育及調控機制仍然十分重要。小鼠心臟因為與人類心臟相似,一直作為研究人類心臟的較好的動物模型[6]。
本研究在前期研究中發現mmu-miR-3475-3P在小鼠胚胎期高表達,提示其可能參與了心臟的早期發育。對該microRNA的Go分析發現該microRNA的靶基因涉及popdc2、pdgfrb、Atp1b1、Atp2a2等基因。而目前的研究表面大部分基因與心臟發育及疾病形成有關。如研究表明popdc2是雞胚胎心臟發育成熟的標志基因[7],而且popdc2存在于大部分哺乳動物的心臟,沉默popdc2基因表達可導致斑馬魚胚胎心臟和骨骼肌發育障礙[8],沉默小鼠popdc2基因表達可導致心臟傳導系統在應激誘導下功能障礙,可誘發小鼠出現竇性心動過緩[9],下調popdc2表達可促進新生大鼠心肌細胞增殖,促其進入細胞循環參與心肌修復,且popdc2在心肌中表達很高,表達量在胚胎期較少,但生后隨著大鼠出生時間增加表達量逐漸升高[10],與mmu-miR-3475-3P的表達呈負相關關系。對microRNA的信號通路分析發現mmu-miR3475-3p的靶基因富集于心臟發育密切相關的Notch、Wnt、Hippo信號通路,已有大量研究揭示這些通路調控著心臟的發育進程。如Notch信號通路調控心肌分化及心臟瓣膜、心室小梁,以及動脈血管發育;Notch信號通路中基因JAG的突變將引起法洛四聯癥的發生[11-12]。而Wnt通路的異常是先天性心臟病的高風險因素[13-14],持續激活內皮細胞的Wnt經典信號通路可導致心力衰竭的發生等[15]。
(2)膠凝材料水化生成的凝膠對Pb2+的吸附作用,Pb2+以類質同相進入AFt和復鹽結構內部等的協同作用,是其固鉛效率顯著高于水泥的重要原因。
本研究證實了mmu-miR3475-3p在小鼠胚胎期和成年心肌中的表達有明顯差異。通過生物信息學分析揭示其可能參與心臟發育及疾病形成。但在生物信息學預測分析中,存在著假陽性或者假陰性結果。mmu-miR3475-3p是否在心臟發育中起到重要的調控作用仍需進一步研究證實,包括經典的過表達和沉默策略研究mmu-miR3475-3p對心肌細胞功能的影響,以及其他分子機制的研究等。
[1]Benziger CP,Roth GA,Moran AE.The global burden of disease study and the preventable burden of NCD[J].Glob Heart 2016,11(4):393-397.
[2]Michael GK,Anthony SF,Andrew PK,et al.The role of microRNAs in cardiac development and regenerative capacity [J].Am J Physiol Heart Circ Physiol,2016,310(5):H528-541.
[3]Bao MH,Feng X,Zhang YW,et al.Let-7 in Cardiovascular diseases,heart development and cardiovascular differentiation from stem cells [J].Int J Mol Sci,2013,14(11):23086-23102.
[4]Jian D,Mao N,Jianming L,et al.Trbp is required for differentiation of myoblasts and normal regeneration of skeletal muscle[J].PLoS One,2016,11(5):e0155349.
[5]Xiaomin Z,Gohar A,Scott AH,et al.Regulation of cardiac microRNAs by serum response factor[J].J Biomed Sci,2011,18(1):15-28.
[6]Anita K,Rajeev S,Preeta DH,et al.Detailed comparison of mouse and human cardiac development[J].Pediatr Res,2014,76(6):500-507.
[7]Breher SS,Mavridou E,Brenneis C,et al.Popeye domain containing gene 2(Popdc2) is a myocyte-specific differentiation marker during chick heart development[J].Dev Dyn,2004,229(3):695-702.
[8]Kirchmaier BC,Poon KL,Schwerte T,et al.The Popeye domain containing 2(popdc2) gene in zebrafish is required for heart and skeletal muscle development[J].Dev Biol,2012,363(2):438-450.
[9]Simrick S,Schindler RF,Poon KL,et al.Popeye domain-containing proteins and stress-mediated modulation of cardiac pacemaking[J].Trends Cardiovasc Med,2013,23(7):257-263.
[10]魏飛宇,呂麗,馬芳芳,等.Popdc2表達下調通過Akt磷酸化促進新生大鼠心肌細胞增殖[J].南京醫科大學學報(自然科學版),2015,35(7):938-944.
[11]Kerr BA,West XZ,Kim YW,et al.Stability and function of adult vasculature is sustained by Akt/Jagged1 signalling axis in endothelium[J].Nat Commun,2016(7):10960.
[12]Hofmann JJ,Briot A,Enciso J,et al.Endothelial deletion of murine Jag1 leads to valve calcification and congenital heart defects associated with Alagille syndrome[J].Development,2012,139(23):4449-4460.
[13]Soemedi R,Wilson IJ,Bentham J,et al.Contribution of global rare copy-number variants to the risk of sporadic congenital heart disease[J].Am J Hum Genet,2012,91(3):489-501.
[14]Nagy II,Railo A,Rapila R,et al.Wnt-11 signalling controls ventricular myocardium development by patterning N-cadherin and beta-catenin expression[J].Cardiovasc Res,2010,85(1):100-109.
[15]Nakagawa A,Naito AT,Sumida T,et al.Activation of endothelial β-catenin signaling induces heart failure[J].Sci Rep,2016(6):25009.
Bioinformaticsanalysisformmu-miR-3475-3Pincardiacdevelopment*
ShenXing1,2,PanBo1,ZhouHuiming1,LiuLinjuan1,TianJie1
(1.DepartmentofCardiology,AffiliatedChildren′sHospitalofChongqingMedicalUniversity,Chongqing400014,China;2.SecondDepartmentofPediatrics,AffiliatedHospitalofSouthwestMedicalUniversity,Luzhou,Sichuan646000,China)
ObjectiveTo investigate the mmu-miR-3475-3P possible participating in biology process and signal pathway.MethodsThe expression of mmu-miR-3475-3P in the mature mouse heart and in the embryonic mouse heart was measured by qRT-PCR.The target genes were predicted through comprehensively using the common microRNA on-line databases such as TargetScan,miRDB and miRanda,and then the obtained targeted genes were performed the gene function annotation and signal pathway analysis.ResultsThere was significant difference between the expression of mmu-miR-3475-3P in the embryonic mouse heart and the expression in the mature heart.Bioinformatics analysis by using TargetScan,miRDB and MiRanda on miR3475-3P revealed that microRNA was likely to regulate 441 target genes.Conclusionmmu-miR-3475-3P is highly expressed in the embryonic mouse heart.The target genes predicted by mmu-miR-3475-3P are enriched in multiple signal pathways and cellular biological processes.
mmu-miR-3475-3P;bioinformatics analysis;target gene;cardiomyogenesis
R725.4
A
1671-8348(2017)26-3671-03
2017-02-22
2017-06-21)
《重慶醫學》對臨床研究論文醫學倫理學要求
《重慶醫學》編輯部
10.3969/j.issn.1671-8348.2017.26.025
瀘州市科技廳-西南醫科大學聯合課題[2015lzcyds05(9/12)]。
沈興(1976-),副教授,在讀博士,主要從事兒童心血管疾病基礎與臨床方面研究。
凡投本刊的涉及人的生物醫學研究論文,作者應說明所在用的試驗程序是否經過倫理審查委員會(單位性的、地區性的或國家性的)評估,注明批準號。涉及患者(受試者)的應簽訂知情同意書。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
