煙曲霉Bem46基因敲除株構建及其功能初步研究
2017-09-03 10:17:28李佳娟馮文莉
中國人獸共患病學報 2017年8期
關鍵詞:生長
李 雯,李佳娟,馮文莉,馬 彥
煙曲霉Bem46基因敲除株構建及其功能初步研究
李 雯,李佳娟,馮文莉,馬 彥
目的構建煙曲霉Bem46基因敲除株,初步明確Bem46基因在煙曲霉生長出芽和各種壓力傳導中的作用,以及對細胞壁形成的影響。方法采用生物信息學方法查找煙曲霉中Bem46基因,設計引物,并以pyrG作為篩選標記構建煙曲霉Bem46基因敲除株(ΔBem46)。觀察含不同壓力物質培養基上對照菌株和ΔBem46徑向生長速度,觀察兩菌株發芽狀況以及在含有細胞壁抑制劑培養基上的生長狀況。結果通過序列比對在煙曲霉基因組中找到了Bem46基因,其編號為Afu7g04660,由1 116 bp堿基組成,編碼311個氨基酸。使用原生質體法得到煙曲霉ΔBem46并經PCR和Southernblot確認。經觀察在含有山梨醇作為滲透壓力來源的培養基上ΔBem46生長明顯較對照菌株快。顯微鏡下觀察到在GMM液體培養基中,ΔBem46發芽速度慢于對照株。結論煙曲霉Bem46基因涉及由山梨醇引起的滲透壓壓力傳導,并且該基因對促進孢子發芽可能有作用。
Bem46;煙曲霉;發芽率;信號傳導
煙曲霉是自然界常見的一種絲狀真菌,在正常人群并不會由于經常暴露于煙曲霉而發生感染。然而近年來隨著新技術的開展和免疫抑制劑的應用,免疫力低下的患者往往由于暴露于煙曲霉而引起侵襲性曲霉病的發生,其發病率在臨床呈現明顯上升趨勢且病死率極高,因此探討煙曲霉感染的發病機制刻不容緩。煙曲霉致病同多種因素有關,而菌株在宿主體內的生長發芽,快速適應宿主環境是致病的重要環節[1]。出芽蛋白Bem46是α/β水解酶超家族成員,在進化上相對保守,在模式真菌粗糙脈孢霉(Neurosporacrassa)中出芽蛋白Bem46在其子囊孢子發芽及菌絲形成的過程中有重要作用[2]。該基因在常見的致病真菌如白念珠菌、煙曲霉中的作用目前尚不了解,為了進一步了解其在煙曲霉致病中的作用,幫助我們深入了解煙曲霉的致病機制,發現可能的新藥物靶點,我們對其進行了相關的研究。
1 材料與方法
1.1 菌株和質粒 煙曲霉Ku80作為對照菌株,煙曲霉轉化用宿主菌為Ku80 pyrG-,該菌株為嘧啶營養缺陷株,在沒有尿苷和尿嘧啶的培養基上不能生長。篩選標記pyrG來源于質粒pJW24(由杜克大學William J. Steinbach教授贈送)。
1.2 培養基 GMM基礎培養基(1L):20×鹽溶液50 mL,微量元素液1 mL,D-葡萄糖10 g,瓊脂15 g(1.5%),pH值6.5;溶壁酶: Vinoflow FCE (Novozymes公司);Osmotic Media:1.2 mol/L MgSO4, 10 mmol/L sodium phosphate,4 ℃保存;STC溶液:1.2 mol/Lsorbitol, 10 mmol/L CaCl2, 10 mmol/L Tris-HCl pH 7.5,在4 ℃保存;PEG-CaCl2溶液:60% PEG, 50 mmol/L CaCl2, 50 mmol/L Tris-HCl pH 7.5在室溫(25 ℃)保存。0.7%頂層瓊脂培養基(SMM) : 20×鹽溶液 50 mL,微量元素液1 mL,葡萄糖10 g,瓊脂7 g,山梨醇218.6 g,酵母提取物 1g,pH值6.5,加水至1 L,高壓滅菌20 min。
1.3 生物信息學分析 在煙曲霉基因組中(www.aspergillusgenome.org)找出與N.crassa同源的Bem46基因開放讀碼框及其上下游各約1 000 bp序列,進行分析,并與常見致病真菌進行同源性比對。
1.4 構建Bem46基因敲除株
1.4.1Bem46基因克隆及敲除載體的構建 煙曲霉Ku80 pyrG-在過夜培養,收集菌絲;采用冷凍干燥法提取基因組DNA。設計引物分別擴增Bem46基因5′和3′側翼序列各約1.0 kb的DNA序列。使用限制性內切酶EcoRI,SalI,XbaI和BamHI,將相應克隆片段連接入質粒pJW24,最后得到目的片段產物大小4 869 bp,定量5μg后進行煙曲霉原生質體的轉化,具體實驗方法見文獻[3]。試驗中用到的引物見表1。
1.4.2 原生質體法構建Bem46基因敲除株 Ku80 pyrG-菌懸液2 mL,30 ℃,250 r/min搖床培養過夜培養(16~20 h)。40mLOsmotic Media中溶解3 g Vinoflow FCE,加入收集的菌絲并混勻,28 ℃,75 r/min搖床培養3~4 h。4 ℃,3 500 r/min離心10 min,吸出離心管夾層的原生質體,并加3倍體積的STC混勻,離心。吸出原生質體層,用1 mL的STC重懸原生質體。取5 μg的融合酶切產物加入到200 μL的原生質體中,冰上靜置1 h。將200 μLDNA-原生質體放入到1.25 mL的PEG-CaCl2中混勻,室溫(25 ℃)放置20 min,加入4 mLSTC混勻,取400 μL與10 mL SMM頂層培養基混合后鋪板,放置于37 ℃孵箱中培養。具體實驗方法見文獻[4]。
1.5 Bem46基因敲除株驗證
1.5.1 首先使用PCR法進行驗證 在GMM液體培養基中過夜培養單克隆轉化子,并提取基因組DNA。并設計相關引物進行驗證,驗證用引物見表1。1.5.2 通過Southernblot進行驗證 設計探針,具體實驗步驟按照試劑盒說明進行。探針設計上下游引物序列命名為Bem46-probe-F,Bem46-probe-R 具體序列見表1。
1.6 Bem46相關功能研究
1.6.1 Ku80與ΔBem46在不同生長條件下的生長狀況對比。生長條件分別為:37 ℃時GMM基礎培養基、含卡泊芬凈(1 μg/mL和4 μg/mL)和尼可霉素(1 μg/mL)的培養基、含過氧化氫(1 mmol/L, 2 mmol/L,3 mmol/L)和甲酰胺(10 μmol/L和50 μmol/L)的培養基、含有氯化鈉(1 mol/L)和山梨醇(1.2 mol/L)的培養基,及55 ℃時GMM基礎培養基。分別將Ku80與ΔBem46孢子10 μL( 1×106個)點種于平皿中央,每日記錄菌落直徑,并拍照。
1.6.2 孢子發芽率實驗 分別取100 μL濃度為1×107個/mL的Ku80及ΔBem46菌懸液,并分別加入到10 mL的GMM液體培養基,在37 ℃以200 r/min轉速的搖床孵育。從腫脹孢子階段(大約4 h)開始,每30 min在顯微鏡下觀察一次,并計數孢子的發芽百分率,重復3次。
1.6.3 統計學軟件 采用SPSS22.0重復測量方差分析軟件對結果進行統計學分析,P<0.05為差異有統計學意義。
2 結 果
2.1 生物信息學分析結果 在煙曲霉基因組中通過序列比對找到了Afu7g04660,1 116 bp個堿基編碼311個氨基酸。380-715堿基間編碼水解酶的保守區域。在細胞極性,孢子發芽,以及定位在內質網膜都有重要的意義。序列比對見圖1,可知煙曲霉Bem46基因同常見致病菌種的同源基因有較強的同源性。
2.2 構建Bem46基因敲除載體結構 如圖2Bem46全基因使用篩選標記pyrG替代。設計探針并使用KpnI酶切, Suthernblot驗證時在對照菌株和ΔBem46產生不同條帶,分別為4.8 kb和2.3 kb。
2.3 基因敲除株驗證結果 經過原生質體法轉化煙曲霉共得到6株轉化子,轉化子的驗證主要通過PCR法和Southernblot進行驗證。

表1 試驗中使用到的引物Tab.1 Primer were used in the construction of ΔBem46
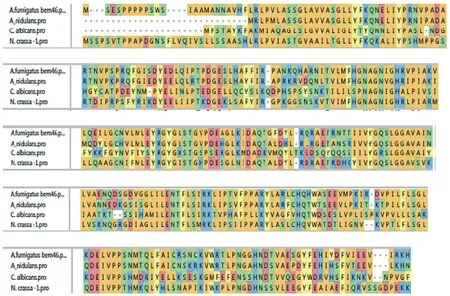
圖1 煙曲霉,構巢曲霉,白念珠菌和粗糙脈孢霉中 Bem46基因序列同源性比對Fig.1 Homologous sequence alignment results of Bem46 gene among the A. fumigatus, A. nidulans, Candida albicans and Neurospora crassa
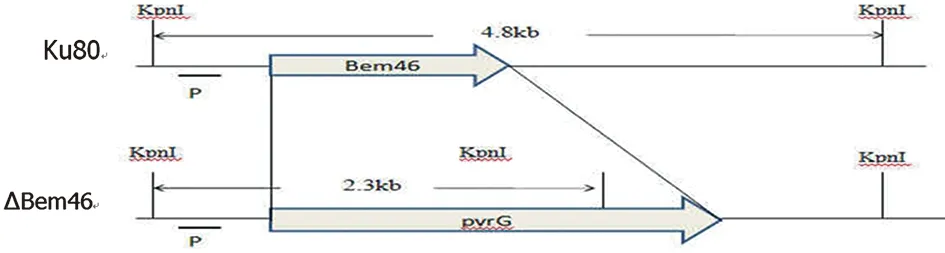
圖2 構建Bem46基因敲除載體示意圖Fig.2 Strategy for deleting Bem46 gene by replacing with pyrG gene by homologous recombination
2.3.1 驗證 根據Bem46序列設計了如下引物擴增和驗證使用了2對引物來進行轉化子的驗證,第1對引物證明有篩選標記的插入,第2對引物證明基因敲除株同對照之間的不同。具體實驗用引物見表1。
2.3.2 用PCR及Southernblot驗證ΔBem46 圖3a為PCR驗證所需引物擴增Ku80及ΔBem46引物的相應片段大小;圖3b為使用第1對引物PCR驗證結果;圖3c為使用第2對引物PCR驗證結果;圖3d為Southernblot驗證結果。
由圖3b可見對照菌株未擴增出該基因,而轉化子1,2,4,5,6,7均有篩選標記擴增成功,說明轉化子中有pyrG成功插入;由圖3c可見只有2號和4號擴增出大約5.3 kb的片段;根據煙曲霉Bem46基因敲除構建示意圖可見對基因DNA酶切后進行探針標記驗證,在對照菌株和煙曲霉Bem46基因敲除株應該有2條明顯不同的條帶,在煙曲霉Bem46基因敲除大約可見2.3 kb大小條帶,而在對照菌株可見約為4.8 kb大小的條帶。由圖3d可見在2號菌株確實只有一條大小約為2.3 kb的條帶,而對照菌株和1、3、5號基因敲除株均有大小約為4.8 kb的條帶,說明只有2號菌株為正確的基因敲除株。該菌株用來進行后續的實驗。
2.4 觀察Bem46基因對煙曲霉相關功能的影響:
2.4.1 對照菌株與ΔBem46在不同培養基上的生長狀況 圖4a為二者在普通GMM培養基上的生長比較;圖4b為二者在含有卡泊芬凈和尼可霉素2種藥物的培養基上生長結果比較;圖4c為在含有不同濃度過氧化氫培養基上二者菌落直徑生長速度比較;圖4d為在含有不同濃度甲酰胺培養基上二者生長速度比較;圖4e為在含有氯化鈉、山梨醇的培養基上和不同溫度條件下二者生長結果比較。
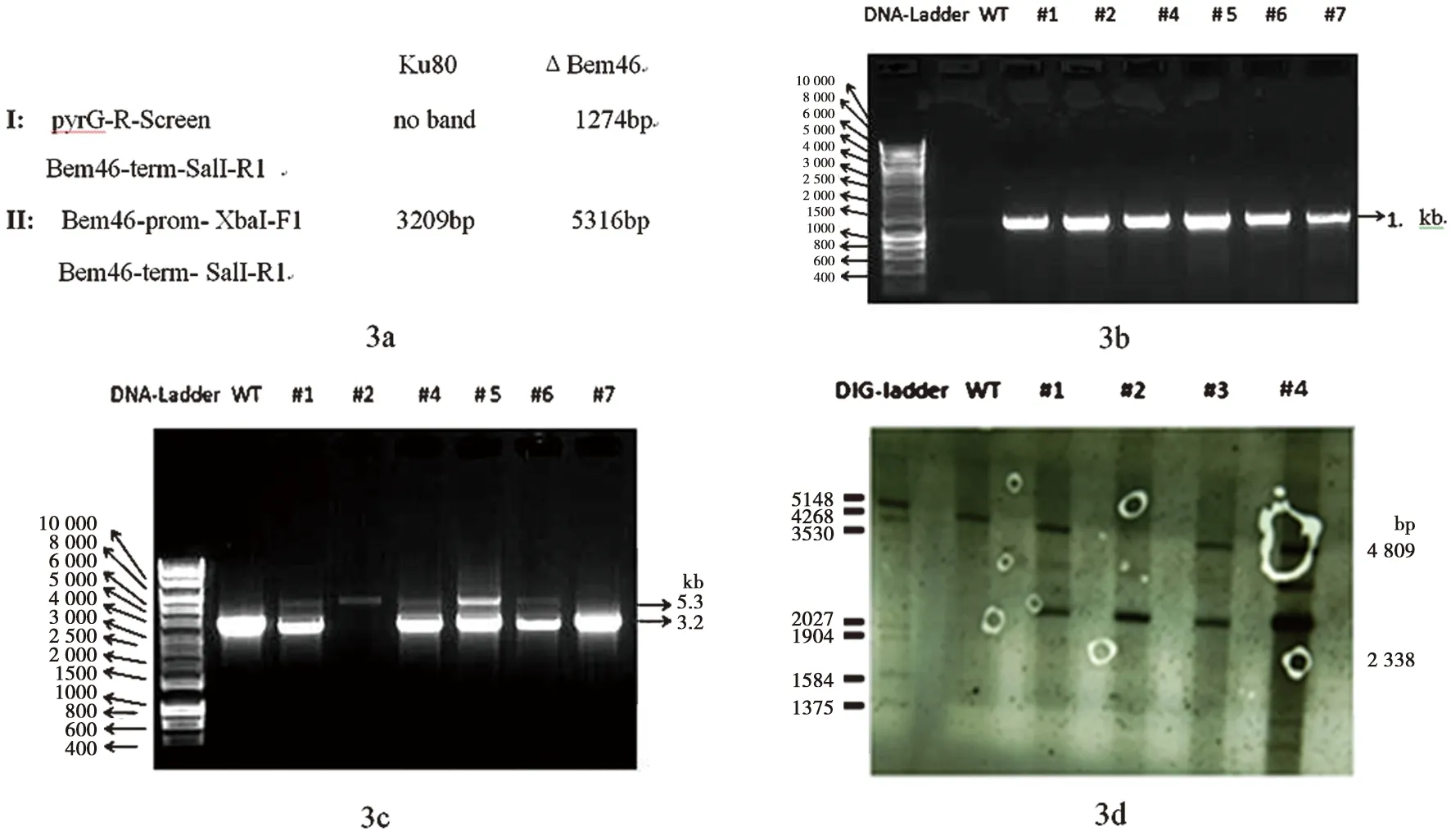
圖3 PCR及Southernblot驗證ΔBem46Fig.3 Confirmation of ΔBem46 by PCR and Southernblot
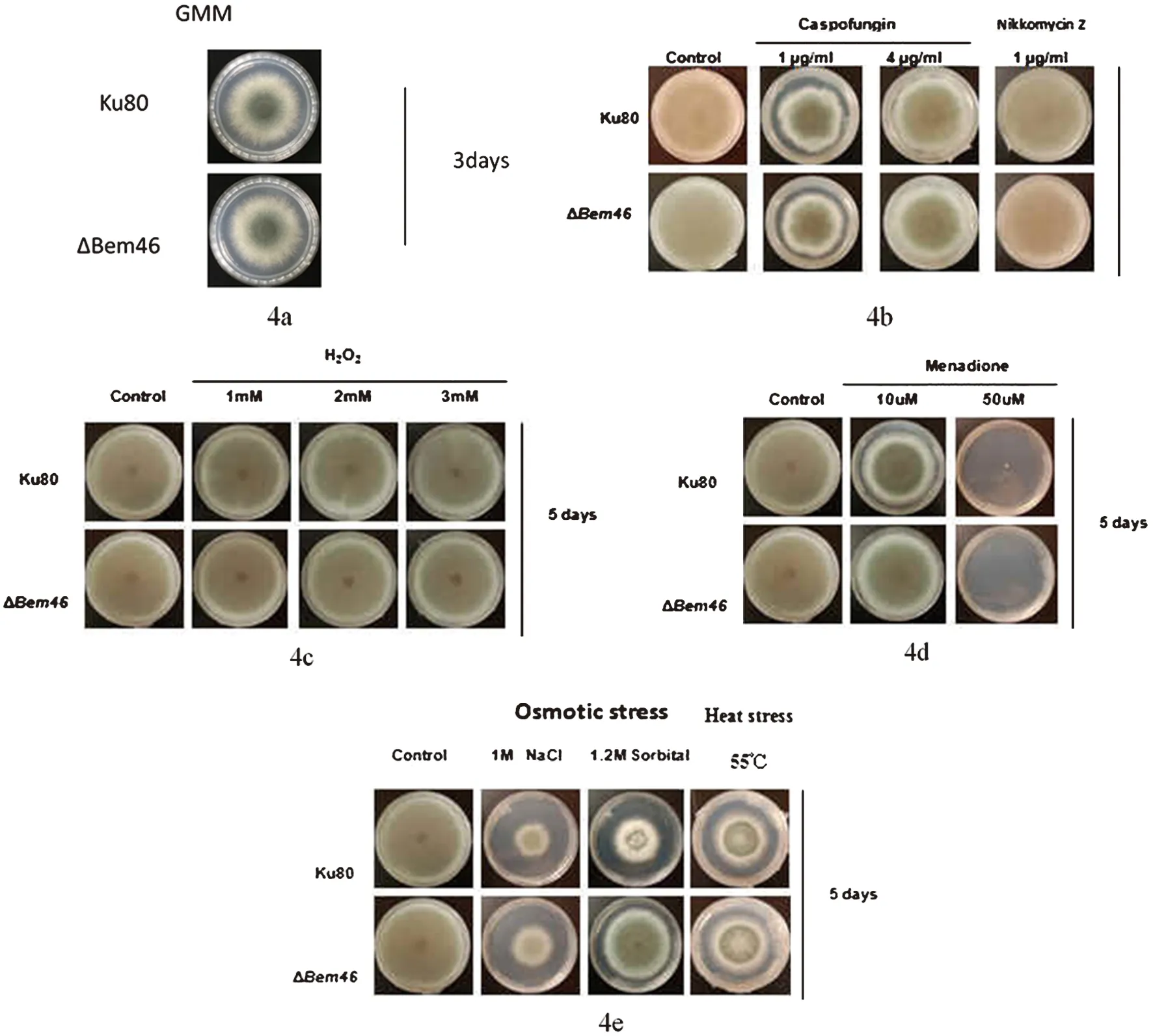
圖4 Ku80與ΔBem46在不同培養基上的生長狀況比較Fig.4 Comparison on the growth diameter of the Ku80 and the ΔBem46 on different medium
實驗結果顯示在GMM培養基上,Ku80與ΔBem46生長狀況無明顯差異;在對抑制真菌細胞壁藥物敏感試驗中可見煙曲霉對照菌株與Bem46基因敲除株在含有卡泊芬凈和尼可霉素這2種藥物的平皿上生長無明顯差異;對氧化壓力敏感性試驗中,對照菌株與ΔBem46在含有不同濃度過氧化氫的培養基、及含有不同濃度甲酰胺的培養基上二者的生長狀況無明顯差異;對滲透壓敏感試驗和溫度敏感試驗中,55 ℃條件下,二者生長狀況無明顯差異,以氯化鈉介導的滲透壓變化也沒有對二者的生長狀況產生影響,但在以山梨醇介導的滲透壓變化使得二者的生長狀況產生了差異,即在濃度為1.2 mol/L的山梨醇培養基上,煙曲霉Bem46基因缺陷株明顯較對照菌株Ku80生長要快。以上菌落直徑結果均經SPSS22.0軟件分析。
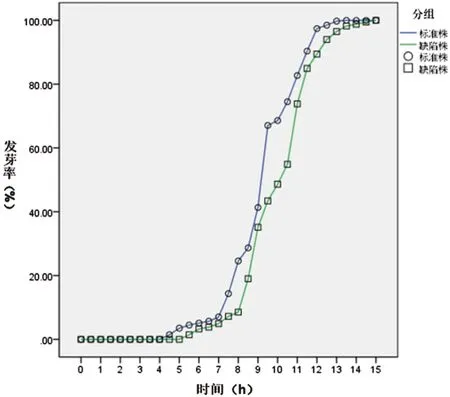
圖5 Ku80與ΔBem46發芽率百分率比較Fig.5 Comparison on the germination rate of the Ku80 and the ΔBem46
2.4.2 觀察煙曲霉Bem46基因在孢子發芽中的作用見圖5,經SPSS22.0軟件分析,由圖可見,在13 h時,對照株Ku80已經完全發芽,而缺陷株ΔBem46在15 h時才實現完全發芽,缺陷株ΔBem46出芽明顯滯后于對照株Ku80(F=3536.191,P<0.05)。15 h后二者均全部發芽。
3 討 論
煙曲霉是主要的致病絲狀真菌,尤其在中性粒細胞減少的患者中感染煙曲霉可以發展成為侵襲性曲霉病,如果造成播散型感染就可能有較差的預后[5]。盡管在診斷技術和抗真菌藥物的研究上取得了很大的進步,但是侵襲性曲霉病的發病和致死率仍然很高,也就顯得對煙曲霉生理生化性質以及其同煙曲霉的致病性,抗真菌藥物敏感性相關性的一些細節研究更有必要[6]。目前對Bem46蛋白了解有限,只對模式絲狀真菌N.crassa有所研究,在N.crassa中該基因在子囊孢子發芽是必須的,且該基因定位到內質網膜[2],在N.crassa該基因編碼的蛋白在細胞特異性種類極性的形成是必不可少的,Bem46基因定位在內質網周,在靠近細胞膜處形成斑點狀[7-8]。鑒于N.crassa同致病真菌煙曲霉基因同源性較高,是非常好的絲狀真菌的模式菌,我們推測該基因可能在煙曲霉中有類似作用。因而我們構建了該基因缺陷株。通過原生質體法我們成功獲得煙曲霉Bem46基因缺陷株ΔBem46,并且觀察了其對煙曲霉一般生長性狀的影響。在基礎培養基上,我們發現該基因的缺失對于煙曲霉的生長沒有明顯的影響。煙曲霉細胞壁的合成是其生長過程中重要的一環,如果合成出現異常,則可能為我們提供新的藥物靶點。因此,我們選擇了臨床常用的2種影響細胞壁合成的藥物卡泊芬凈和尼克霉素Z。卡泊芬凈主要抑制β-1,3葡聚糖合成酶,阻礙葡聚糖的形成,而尼克霉素Z的作用機制抑制細胞壁中幾丁質的合成,這2種藥物從不同的方面來驗證該基因是否在煙曲霉細胞壁形成過程中發揮作用,由我們的結果可見Bem46這個基因在含有此2種物質的培養基上生長速度同對照菌株并沒有明顯差異,因此我們推測該基因可能并不參與煙曲霉細胞壁的合成。以往的研究表明氧化壓力、滲透壓力以及溫度對于煙曲霉的致病都有重要的意義[9],因此我們進一步研究了該基因在氧化壓力、滲透壓力和溫度敏感性中的作用。本研究顯示該基因對于過氧化氫和甲酰胺產生的氧化壓力和對照菌株沒有明顯的差異,也就是對這兩種物質產生的氧化壓力不敏感,可能不參與其產生的氧化壓力的傳導。在滲透壓力的敏感性中,由氯化鈉產生的滲透壓力并未對二者生長產生不同的影響,但同時發現基因缺陷株ΔBem46對山梨醇產生的滲透壓力呈現陽性反應,即相比野生對照菌株而言其生長明顯加快,推測也許Bem46參與了由山梨醇介導的滲透壓力傳導通路,在山梨醇存在的條件下,Bem46基因有抑制生長的作用。在比較二者孢子發芽的實驗中發現,缺陷株實現完全發芽的時間明顯慢與對照株實現完全發芽的時間,即Bem46基因的缺失導致了其發芽滯后,而15 h后二者均全部發芽,表明Bem46基因的缺失并不會阻止煙曲霉孢子發芽。所以我們推測,Bem46基因極有可能有促進孢子出芽的作用,參與孢子的出芽生長過程,但其并不是煙曲霉孢子出芽的決定性基因。
總之,研究通過構建煙曲霉Bem46基因缺陷株,初步了解該基因對煙曲霉生長狀況的影響,提示可以從山梨醇介導的滲透壓傳導通路入手,研究Bem46基因對煙曲霉生長的抑制作用;Bem46基因缺陷使得發芽速度減慢提示可從參與孢子出芽過程的基因入手,研究其對出芽生長的傳導通路產生的影響;研究也將進一步進行相關的實驗,以期發現該基因更多更有意義的性狀。
[1] Abad A, Fernandez-Molina JV, Bikandi J, et al. What makesAspergillusfumigatusa successful pathogen? Genes and molecules involved in invasive aspergillosis[J]. Rev Iberoam Micol, 2010, 27(4): 155-182. DOI:10.1016/j.riam.2010.10.003
[2] Mercker M, Kollath-Leiss K, Allgaier S, et al. The BEM46-like protein appears to be essential for hyphal development upon ascospore germination inNeurosporacrassaand is targeted to the endoplasmic reticulum[J]. Curr Genet, 2009, 55(2): 151-161.DOI:10.1007/s00294-009-0232-3
[3] Fan H,Guo JY,Ma SL,et al.Construction and identification of eukaryotic expression plasmids of synthetic TAG gene ofPneumocystisp55 antigen[J].Chin J Zoonoses,2015,31(5):399-402.DOI: 10.3969/j.issn.1002-2694.2015.05.002(in Chinese)
樊華,國九英,馬素麗.等. 人工合成肺孢子菌p55抗原串聯基因真核表達載體的構建及表達[J].中國人獸共患病學報,2015,31(5):399-402.
[4] Steinbach WJ, Cramer RA, Jr.Perfect BZ, et al. Calcineurin controls growth, morphology, and pathogenicity inAspergillusfumigatus[J]. Eukaryot Cell, 2006, 5(7): 1091-1103.DOI:10.1128/EC.00139-06
[5] Segal BH. Aspergillosis[J]. N Engl J Med, 2009, 360(18): 1870-1884.DOI:10.1056/NEJMra0808853
[6] Erjavec Z, Kluin-Nelemans H, Verweij PE. Trends in invasive fungal infections, with emphasis on invasive aspergillosis[J]. Clin Microbiol Infect, 2009, 15(7): 625-633.DOI:10.1111/j.1469-0691.2009.02929.x
[7] Martinez-Martinez S, Rodriguez A, Lopez-Maderuelo MD, et al. Blockade of NFAT activation by the second calcineurin binding site[J]. J Biol Chem, 2006, 281(10): 6227-6235.DOI:10.1074/jbc.M513885200
[8] Kumar A, Kollath-Leiss K, Kempken F. Characterization of bud emergence 46 (BEM46) protein: sequence, structural, phylogenetic and subcellular localization analyses[J]. Biochem Biophys Res Commun, 2013, 438(3): 526-532.DOI:10.1016/j.bbrc.2013.07.103
[9] Ma Y, Qiao J, Liu W, et al. The sho1 sensor regulates growth, morphology, and oxidant adaptation inAspergillusfumigatusbut is not essential for development of invasive pulmonary aspergillosis[J]. Infect Immun, 2008, 76(4): 1695-1701.DOI:10.1128/IAI.01507-07
ConstructionofBem46knockoutstrainandtheroleoftheBem46geneinAspergillusfumigatus
LI Wen, LI Jia-juan, FENG Wen-li, MA Yan
(DepartmentofDermatologyandVenereology,theSecondHospitalofShanxiMedicalUniversity,Taiyuan030001,China)
The aims of this study are to construct theBem46 knockout strain ofAspergillusfumigatus, investigate the effect of theBem46 in the growth and germination, and observe the sensitivity of the oxidative stress, the osmotic stress, the temperature and the response to caspofungin and nikkomycin. TheBem46 gene was identified by sequence alignment inA.fumigatusgenome database. TheBem46 knockout strain was construction by the protoplast. The growth was observed and compared between Ku80 and ΔBem46 on different culture medium, such as GMM, GMM containing hydrogen peroxide, formamide, sodium chloride and sorbitol. The growth diameter was measured under the different temperatures. The germination rate was observed and compared by microscope. Results showed that theBem46 gene, Afu7g04660, contained 1 116 bp bases pair and encoding 311 amino acids. Six transformants were obtained by gene cloning and protoplast method and only one was confirmed by PCR and Southernblot. The ΔBem46 grew significantly faster than the control strain on the medium containing sorbitol. And there were no visible difference between ΔBem46 and Ku80 on the other medium. The germination rate of the ΔBem46 was more retarded than the Ku80 in GMM liquid medium. In conclusion, theBem46 gene plays a role in osmotic stress inA.fumigatusand involved in osmotic pressure induced by sorbitol. And there is no visible effect in oxidative stress. TheBem46 gene has a positive effect on spore germination.
Bem46;Aspergillusfumigatus; germination rate; signal transduction
10.3969/j.issn.1002-2694.2017.08.006
國家青年科學基金項目資助(No.81101233),山西省青年科技研究基金(No.2010021036-2),山西省衛生計生委科研課題(No.2015043),山西省回國留學人員科研資助項目(No.2016-052)聯合資助
馬 彥: Email:mayan197522@163.com
山西醫科大學第二醫院皮膚科,太原 030001
Supported by the National Science Foundation for Young Scientists of China(No. 81101233), the Natural Science Foundation for Youth in Shanxi Province (No.2010021036-2), the Research Project of Health and Family Planning Commission of Shanxi Province (No.2015043), and the Research Project Supported by Shanxi Scholarship Council of China (No.2016-052) Corresponding author: Ma Yan, Email: mayan197522@163.com
R378
:A
:1002-2694(2017)08-0694-06
2016-11-14編輯:梁小潔
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
