豬鏈球菌2型轉(zhuǎn)錄調(diào)控因子Rex原核表達(dá)及體外結(jié)合活性分析
2017-09-03 10:17:28祝昊丹何孔旺范紅結(jié)
中國(guó)人獸共患病學(xué)報(bào) 2017年8期
關(guān)鍵詞:分析
王 勇,祝昊丹,何孔旺,范紅結(jié)
豬鏈球菌2型轉(zhuǎn)錄調(diào)控因子Rex原核表達(dá)及體外結(jié)合活性分析
王 勇1,2,3,祝昊丹1,3,何孔旺1,3,范紅結(jié)2
目的克隆豬鏈球菌2型轉(zhuǎn)錄調(diào)控因子Rex的編碼基因進(jìn)行原核表達(dá)和純化,對(duì)其進(jìn)行生物信息學(xué)分析和體外結(jié)合活性測(cè)定。方法PCR擴(kuò)增豬鏈球菌2型強(qiáng)毒株SS2-1的Rex基因編碼區(qū),將其克隆入pET28a質(zhì)粒中,再將pET28a-Rex重組質(zhì)粒轉(zhuǎn)入大腸桿菌表達(dá)菌BL21(DE3)中,重組菌株經(jīng)IPTG誘導(dǎo)后能表達(dá)出豬鏈球菌Rex蛋白;通過(guò)體外凝膠遷移實(shí)驗(yàn)( EMSA)對(duì)Rex蛋白與DNA的結(jié)合活性進(jìn)行分析。結(jié)果成功地原核表達(dá)并純化了Rex重組蛋白。Rex蛋白與自身啟動(dòng)子Prex特異性結(jié)合,高濃度的NADH抑制兩者的結(jié)合活性,而NAD+對(duì)結(jié)合沒(méi)有影響。結(jié)論通過(guò)體外結(jié)合試驗(yàn)發(fā)現(xiàn)Rex 是通過(guò)響應(yīng)NADH/NAD+平衡來(lái)調(diào)控自身基因的表達(dá)。
豬鏈球菌;Rex;調(diào)控因子
豬鏈球菌(Streptococcussuis)是一種重要的人獸共患病原菌,可導(dǎo)致腦膜炎,關(guān)節(jié)炎,敗血癥等疾病。該病原菌造成仔豬大量死亡,給全球養(yǎng)豬業(yè)造成巨大的經(jīng)濟(jì)損失[1]。通過(guò)莢膜分型,有35個(gè)血清型,其中2型在患病豬和患病人群中分離率是最高的[2-3]。在亞洲地區(qū),豬鏈球菌病是一種重要的公共衛(wèi)生疾病,主要引起成人腦膜炎[4]。1998年和2005年在我國(guó)江蘇和四川暴發(fā)了由豬鏈球菌2型引起的豬鏈球菌感染疫情,給公共衛(wèi)生安全造成了極大的危害,受到國(guó)內(nèi)外廣泛關(guān)注[2,5]。
細(xì)菌在感染過(guò)程中,需要感知復(fù)雜多變的宿主體內(nèi)環(huán)境,調(diào)節(jié)各種相關(guān)基因的表達(dá),以達(dá)到在宿主體內(nèi)生存、繁殖和致病的目的。轉(zhuǎn)錄調(diào)節(jié)因子是有機(jī)體應(yīng)對(duì)動(dòng)態(tài)環(huán)境的一個(gè)重要機(jī)制。Rex是一種廣譜轉(zhuǎn)錄調(diào)節(jié)因子,在細(xì)菌的生存過(guò)程中起重要作用。首先在天藍(lán)色鏈霉菌中發(fā)現(xiàn)轉(zhuǎn)錄調(diào)節(jié)因子Rex響應(yīng)胞內(nèi)NADH/NAD+平衡[6]。之后在糞腸球菌(Enterococcusfaecalis)、金黃色葡球菌(Staphylococcusaureus)、變形鏈球菌(Streptococcusmutans)、枯草芽孢桿菌(Bacillussubtilis)等的轉(zhuǎn)錄調(diào)控均有大量報(bào)道。豬鏈球菌中Rex的功能尚不清楚。
本研究對(duì)SS2強(qiáng)毒株SS2-1的轉(zhuǎn)錄調(diào)控因子Rex進(jìn)行生物信息學(xué)分析,利用大腸桿菌原核表達(dá)系統(tǒng)對(duì)Rex蛋白進(jìn)行體外表達(dá)和純化,研究Rex蛋白與體外靶基因的結(jié)合活性,為后續(xù)深入研究Rex的功能奠定基礎(chǔ)。
1 材料與方法
1.1 菌株、質(zhì)粒和引物 所用的菌株、質(zhì)粒及引物分別見(jiàn)表1和表2,引物由上海英濰捷基公司合成。

表1 菌株及質(zhì)粒Tab.1 Bacterial strains and plasmids used in this study

表2 PCR擴(kuò)增引物Tab.2 Primers used in this study
1. 2 主要試劑 PCR Mix和DNA Marker購(gòu)于Vazyme公司,pMD19-T載體和DNA限制性內(nèi)切酶購(gòu)于TaKaRa公司;質(zhì)粒提取試劑盒和膠回收試劑盒購(gòu)于Axygen;THB和LB培養(yǎng)基購(gòu)于美國(guó)BD公司;NAD+和NADH購(gòu)于 Sigma公司;EMSA/Gel-shift結(jié)合緩沖液和上樣緩沖液購(gòu)于碧云天生物公司。
1.3Rex基因克隆和生物信息學(xué)分析 以SS2-1基因?yàn)槟0澹O(shè)計(jì)引物Rex-F/Rex-R,利用Prime STAR HS DNA聚合酶進(jìn)行PCR擴(kuò)增。PCR擴(kuò)增程序?yàn)椋?4 ℃ 2 min,98 ℃ 15 s,55 ℃ 30s,72 ℃ 1 min,30 個(gè)循環(huán),72 ℃ 5 min。電泳檢測(cè)。回收目的片段。將目的片段回收產(chǎn)物與pMD19-T載體進(jìn)行連接,轉(zhuǎn)化到感受態(tài)細(xì)胞DH5α,挑取陽(yáng)性克隆測(cè)序。使用EditSeq翻譯成氨基酸序列,用Blastp和ClustalW等生物信息學(xué)工具對(duì)Rex氨基酸序列進(jìn)行分析,并用ESPript 3.0 對(duì)ClustalW對(duì)比結(jié)果進(jìn)行二級(jí)結(jié)構(gòu)分析。
1.4 表達(dá)質(zhì)粒的構(gòu)建與鑒定 將含有 pET28a質(zhì)粒的宿主菌DH5α單菌落接種至LB液體(含卡那霉素50 μg/mL)中,37 ℃過(guò)夜培養(yǎng)。按質(zhì)粒提取試劑盒說(shuō)明書抽提質(zhì)粒DNA,以BamH I、XhoI進(jìn)行雙酶切,電泳回收線性化的質(zhì)粒DNA。將質(zhì)粒雙酶切回收產(chǎn)物與雙酶切的PCR產(chǎn)物進(jìn)行連接,轉(zhuǎn)化到感受態(tài)細(xì)胞DH5α,經(jīng)PCR和雙酶切鑒定后,將重組表達(dá)質(zhì)粒pET28a-Rex轉(zhuǎn)化進(jìn)表達(dá)宿主BL21(DE3),挑取陽(yáng)性克隆測(cè)序。
1.5 重組蛋白質(zhì)的誘導(dǎo)表達(dá)與純化 將含重組質(zhì)粒pET28a-REX的大腸桿菌BL21 (DE3) 接種LB液體,37 ℃搖振培養(yǎng)3~4 h,至 OD600值為0.4~0.6時(shí),加入IPTG至終濃度1 mmol/L,繼續(xù)劇烈搖振培養(yǎng)3~4 h。離心去上清,于沉淀中加 SDS-PAGE上樣緩沖液混勻后煮沸10 min,進(jìn)行SDS-PAGE。將重組菌擴(kuò)大培養(yǎng)并經(jīng) IPTG 誘導(dǎo)表達(dá),離心收集菌體,PBS 重懸后冰浴超聲破碎菌體,離心后的上清過(guò)濾,用GE Healthcare公司的His-Tag親和層析柱純化重組蛋白質(zhì)。
1.6 凝膠阻滯分析(Electrophoresis mobility shift assay,EMSA) Rex與啟動(dòng)子DNA體外結(jié)合活性。
1.6.1 Rex與啟動(dòng)子DNA體外結(jié)合 EMSA實(shí)驗(yàn):PCR擴(kuò)增Rex基因啟動(dòng)子區(qū)并對(duì)產(chǎn)物純化回收。反應(yīng)體系(10 μL):蛋白 1 μL,DNA 1 μL,5×Binding buffer 2 μL,H2O26 μL;30 ℃水浴20 min。在反應(yīng)樣品中1 μL 10×Loading buffer,將樣品加至6%變性聚丙烯酰胺凝膠中進(jìn)行電泳。將電泳完畢取下的凝膠放入0.5 μg/mL EB溶液中染色15 min后置于凝膠成像系統(tǒng)中觀察。
1.6.2 NADH和NAD+對(duì)Rex與特異性DNA片段結(jié)合活性的影響 按方法1.6.1研究NADH和NAD+對(duì)Rex與特異性DNA片段結(jié)合活性的影響,結(jié)合反應(yīng)進(jìn)行20 min后,在結(jié)合反應(yīng)樣品中分別添加不同濃度(0、5、10和50 mmol/L)的NADH和NAD+繼續(xù)反應(yīng)15 min,染色觀察。
2 結(jié) 果
2.1Rex生物信息學(xué)分析 SS2-1的Rex蛋白的開放閱讀框ORF由639個(gè)核苷酸組成,編碼213個(gè)氨基酸,預(yù)測(cè)分子量為24 kDa。通過(guò)blastp 比對(duì)后,結(jié)果發(fā)現(xiàn)SS2-1 Rex蛋白與金黃色葡球菌、糞腸球菌、馬鏈球菌獸疫亞種、無(wú)乳鏈球菌、化膿鏈球菌、變形鏈球菌和肺炎鏈球菌中的Rex家族蛋白的氨基酸序列分別具有44.7%、51.0%、71.9%、72.4%、73.8%、76.7%和78.7%的一致性。 以無(wú)乳鏈球菌的Rex蛋白晶體結(jié)構(gòu)Protein data bank(PDB)為模型,使用ClustalW軟件和ESPript 3.0對(duì)其同源蛋白進(jìn)行多重序列比對(duì)和二級(jí)結(jié)構(gòu)分析。N-端是特異的DNA結(jié)合位點(diǎn),以及1個(gè)可結(jié)合DNA靶序列的螺旋-轉(zhuǎn)角-螺旋(Helix-turn-helix)基序,而該部分序列在Rex蛋白家族成員中非常保守(圖1)。結(jié)合NAD(H)的C-端有一個(gè)Rossmann折疊,這個(gè)折疊由典型的2個(gè)β-α-β-α-β結(jié)構(gòu)單元系列組成,通過(guò)α-螺旋,它們一同形成了一個(gè)平行的β-折疊,這是在依賴NAD(H)脫氫酶(如乳酸脫氫酶)中的典型結(jié)構(gòu)特征。
2.2 重組表達(dá)質(zhì)粒鑒定Rex基因PCR擴(kuò)增后,經(jīng)雙酶切、純化,定向克隆到表達(dá)載體 pET28a中,轉(zhuǎn)化到大腸桿菌 DH5α。挑取陽(yáng)性克隆提取質(zhì)粒,圖2為重組表達(dá)質(zhì)粒經(jīng)BamH I和XhoⅠ雙酶切鑒定結(jié)果,這表明目的片段已成功插入到表達(dá)質(zhì)粒中。
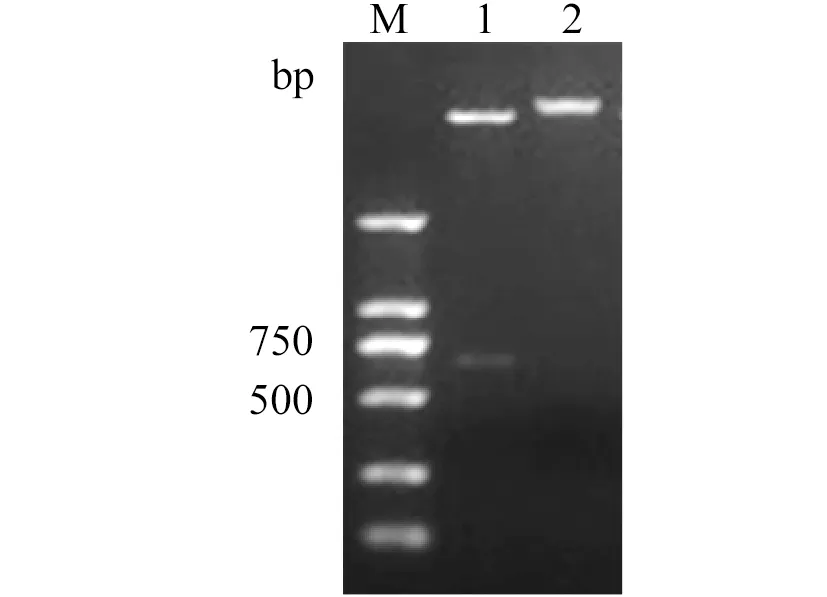
1: BamH I/Xho I double restriction enzymes
2.3 SDS-PAGE分析和Western blotting鑒定 SDS-PAGE分析結(jié)果表明,含重組表達(dá)質(zhì)粒pET28a-Rex的E.coliBL21經(jīng)IPTG誘導(dǎo)后在27 kDa左右處有1條明顯的蛋白條帶,分子量大小與預(yù)期一致(圖3-A)。利用His親和層析柱對(duì)表達(dá)產(chǎn)物進(jìn)行純化,將純化蛋白用12% SDS-PAGE方法進(jìn)行蛋白電泳,轉(zhuǎn)印后,以His-Tag單克隆抗體為一抗進(jìn)行Western blotting鑒定分析(圖3-B)。
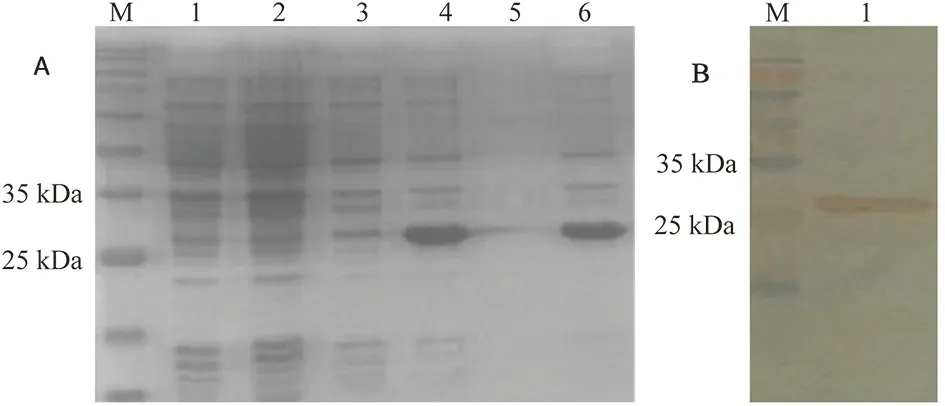
A: SDS-PAGE analysis of the purification.1:Non-induced pET28a/BL21, 2:Induced pET28a/BL21,3:Non-induced pET28a-Rex/BL21,4:Induced pET28a-Rex/BL21,5:Induced pET28a-Rex/BL21 Insoluble fraction, 6:Induced pET28a-Rex/BL21 Soluble fraction. B:Western blot with monoclonal antibody against His-tag. 1: rSsRex圖3 SDS-PAGE 分析和Western blotting鑒定Fig.3 Recombinant protein by SDS-PAGE and Western blotting
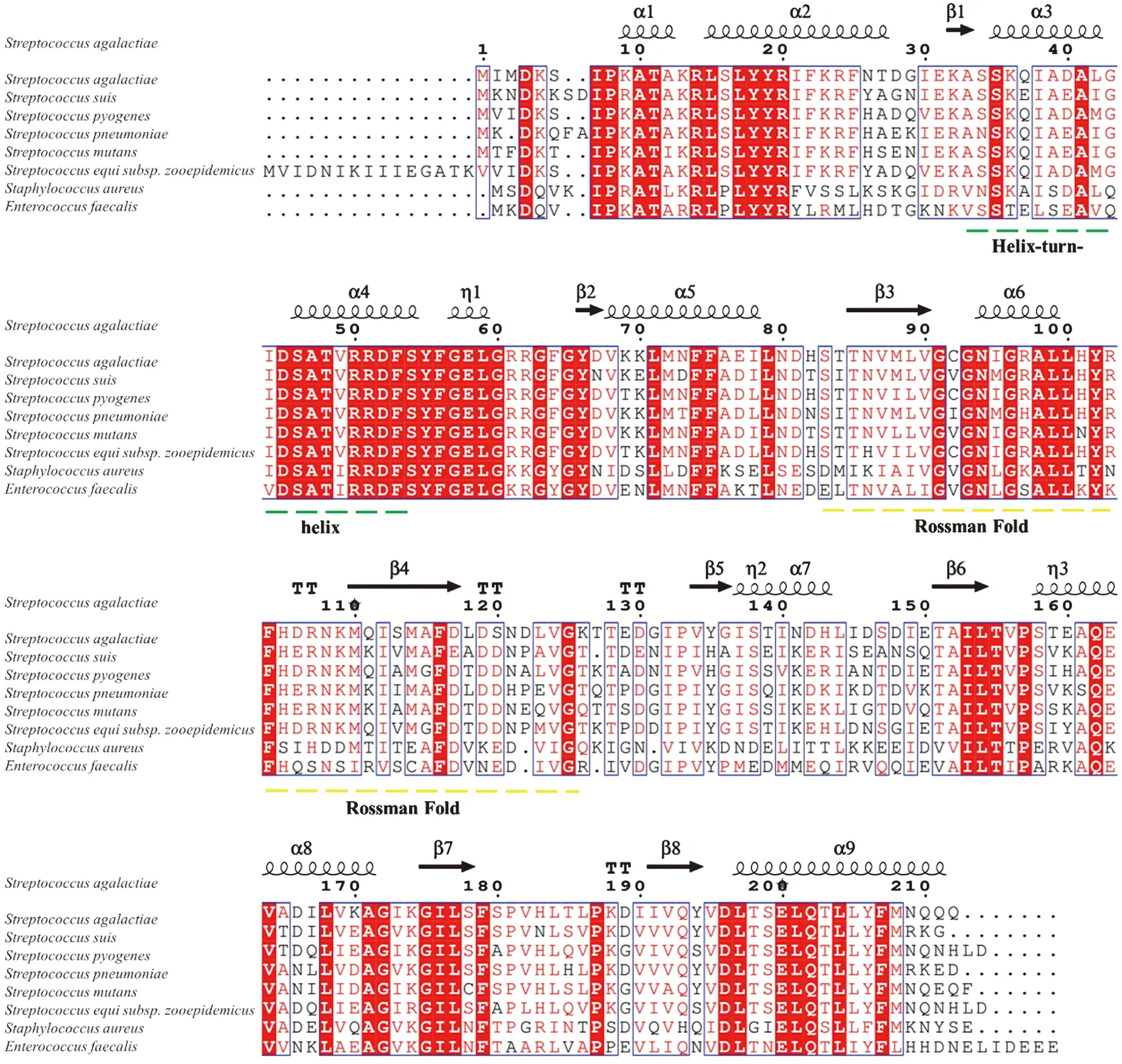
圖1 Rex同源蛋白的多重序列比對(duì)和結(jié)構(gòu)分析Fig.1 Structure-based multiple sequence alignment of Rex ortholog protein
2.4 凝膠阻滯分析(Electrophoresis mobility shift assay,EMSA) Rex與啟動(dòng)子DNA體外結(jié)合活性
2.4.1 Rex與啟動(dòng)子DNA體外結(jié)合 EMSA實(shí)驗(yàn)結(jié)果證實(shí)Rex蛋白能與其自身啟動(dòng)子序列(Prex)特異性的結(jié)合,而非特異性DNA與Rex蛋白不發(fā)生作用(圖4 )。 隨著Rex蛋白濃度的增加 (從0 ng遞增到500ng),結(jié)合的DNA越多,游離的DNA片段越少(圖5 ),初步說(shuō)明Rex蛋白能調(diào)控自身的轉(zhuǎn)錄,且隨著Rex蛋白增多,結(jié)合的Prex也越多。
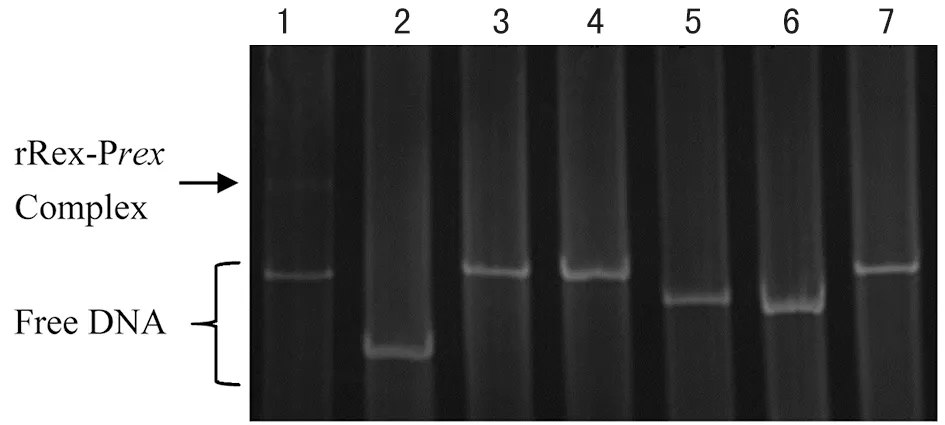
1:Prex; 2,3,4,5,6,7:ive pieces of unspecific DNA; and Rex protein,respectively圖4 胞外Rex蛋白與特異性DNA的結(jié)合Fig.4 In vitro binding of protein Rex with specific DNA oligo

圖5 胞外Rex蛋白與Prex的結(jié)合活性Fig.5 In vitro binding activity between protein Rex and Prex
2.4.2 NADH和NAD+對(duì)Rex與特異性DNA片段結(jié)合活性的影響 競(jìng)爭(zhēng)試驗(yàn)結(jié)果表明NADH抑制Rex蛋白與自身啟動(dòng)子序列Prex的結(jié)合,且隨著濃度的增加抑制效果越強(qiáng)(圖6-A);NAD+則不影響兩者的結(jié)合(圖6-B),與在變形鏈球菌中報(bào)道的研究結(jié)果相一致[8]。

圖6 NADH和NAD+對(duì)Rex與特異性DNA片段結(jié)合活性的影響Fig.6 The effect of NADH and NAD+ on specific DNA oligo binding activity of Rex
3 討 論
細(xì)菌通過(guò)監(jiān)測(cè)代謝產(chǎn)物或特定化合物在細(xì)胞內(nèi)氧化還原狀態(tài)產(chǎn)生相應(yīng)的信號(hào),通過(guò)誘導(dǎo)調(diào)節(jié)蛋白或酶的構(gòu)象變化等方式最終作出生物反應(yīng)[11]。在天藍(lán)色鏈霉菌和枯草芽孢桿菌的研究中,發(fā)現(xiàn)了一種稱為Rex的新型轉(zhuǎn)錄調(diào)節(jié)因子,能直接響應(yīng)胞內(nèi)NADH/NAD+平衡變化。在枯草芽孢桿菌中,當(dāng)有氧呼吸過(guò)程中缺氧時(shí),NADH濃度增加,Rex抑制的基因轉(zhuǎn)錄被激活,這使具有更高氧氣親和力細(xì)胞色素bd和乳酸脫氫酶的形成增加,以保證氧的有效利用和多余的NADH的循環(huán)利用。在金黃色葡萄球菌,Rex調(diào)節(jié)厭氧發(fā)酵和NAD+再生途徑。在變形鏈球菌中Rex對(duì)生物被膜形成和氧化應(yīng)激起重要作用。
通過(guò)對(duì)強(qiáng)毒株SS2-1基因組中編碼Rex蛋白的氨基酸序列進(jìn)行序列比對(duì)分析,發(fā)現(xiàn)豬鏈球菌Rex蛋白與金黃色葡萄球菌、糞腸球菌、馬鏈球菌獸疫亞種、無(wú)乳鏈球菌、化膿鏈球菌、變形鏈球菌和肺炎鏈球菌中的Rex家族蛋白的氨基酸序列高度一致;結(jié)構(gòu)域分析發(fā)現(xiàn)Rex家族蛋白的N端都具有DNA結(jié)合蛋白中常見(jiàn)的螺旋-轉(zhuǎn)角-螺旋(Helix-turn-helix)基序,且非常保守(圖1),保證了Rex蛋白能夠結(jié)合特異性DNA序列從而調(diào)控靶基因的表達(dá),提示豬鏈球菌Rex作為Rex調(diào)控蛋白家族成員的一員,具有其共同的結(jié)構(gòu)特征和類似的調(diào)控機(jī)制。
利用大腸桿菌原核表達(dá)系統(tǒng)成功表達(dá)并純化了強(qiáng)毒株SS2-1的Rex蛋白。體外EMSA實(shí)驗(yàn)證實(shí)了Rex蛋白對(duì)自身啟動(dòng)子序列Prex具有直接的結(jié)合活性,說(shuō)明在豬鏈球菌中Rex蛋白可直接作用于自身基因啟動(dòng)子區(qū)從而直接調(diào)控自身的轉(zhuǎn)錄。此外,EMSA實(shí)驗(yàn)還表明NADH和NAD+都能結(jié)合Rex蛋白,但只有NADH能使Rex失去對(duì)Prex的親和力,而 NAD+對(duì)結(jié)合沒(méi)有影響,即NADH和NAD+競(jìng)爭(zhēng)結(jié)合Rex,提示在豬鏈球菌中Rex發(fā)揮著氧化還原調(diào)控作用。該結(jié)果為我們進(jìn)一步研究Rex的功能奠定了基礎(chǔ)。
[1] Goyette DG, Auger JP, Xu J, et al.Streptococcussuis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing [J]. Emerg Microbes Infect, 2014, 3(6): e45. DOI: 10.1038/emi.2014.45
[2] Feng Y, Zhang H, Wu Z, et al.Streptococcussuisinfection: an emerging/reemerging challenge of bacterial infectious diseases [J]. Virulence, 2014,5(4):477-497. DOI: 10.4161/viru.28595
[3] Liu HZ, Yuan XM. Research advances in clinical laboratory diagnosis ofStreptococcussuisinfection[J]. Chin J Zoonoses,2016,32(5):490-493. DOI: 10.3969/j.issn.1002-2694.2016.05.001
劉海珠,袁曉明.人豬鏈球菌感染的臨床實(shí)驗(yàn)室診斷研究進(jìn)展[J].中國(guó)人獸共患病學(xué)報(bào), 2016,32(5):490-493.
[4] Gong G, Yi XCM, Luo RB,et al. Isolation and identification of strain SS and antibiotic resistance in Tibet pig[J]. Chin J Zoonoses, 2016,32(11):987-990. DOI: 10.3969/j.issn.1002-2694.2016.011.008(in Chinese)
貢嘎,以西措姆,羅潤(rùn)波.藏豬鏈球菌的分離鑒定及耐藥性研究[J].中國(guó)人獸共患病學(xué)報(bào), 2016,32(11):987-990.
[5] Tang J, Wang C, Feng Y, et al. Streptococcal toxic shock syndrome caused byStreptococcussuisserotype 2 [J]. PLoS Med, 2006, 3: e151. DOI: 10. 1371/journal. pemed. 0030151
[6] Brekasis D, Paget M S. A novel sensor of NADH/NAD+redox poise inStreptomycescoelicolorA3(2) [J]. EMBO J, 2003,22(18):4856-4865. DOI: 10.1093/emboj/cdg453
[7] Wang E, Bauer MC, Rogstam A, et al. Structure and functional properties of theBacillussubtilistranscriptional repressor Rex[J]. Mol Microbiol,2008, 69:466-478. DOI:10.1111/j.1365-2958.2008.06295.x
ProkaryoticexpressionofRextranscriptionregulatorinStreptococcussuis2andinvitrobindingassay
WANG yong1,2,3,ZHU Hao-dan1,3,HE Kong-wang1,3,FAN Hong-jie2
(1.InstituteofVeterinaryResearch,JiangsuAcademyofAgriculturalSciences,KeyLaboratoryofEngineeringResearchofVeterinaryBio-products,MinistryofAgriculture,Nanjing210014,China; 2.CollegeofVeterinaryMedicine,NanjingAgriculturalUniversity,Nanjing210095,China; 3.KeyLabofFoodQualityandSafetyofJiangsuProvince-StateKeyLaboratoryBreedingBase,Nanjing210014,China)
We cloned and prokaryoticly expressed the gene encoding Redox regulator(Rex)ofStreptococcussuisserotype 2 and analyzed biological information andinvitrobinding activity. The encodingRexgene of SS2-1 strain was amplified by PCR with the designed primers, and then cloned into prokaryotic expression plasmid pET28a. The recombinant plasmid pET28a-Rex was transformed intoE.coliBL21. After induced expression by IPTG, the Rex protein was obtained. The binding activity of Rex protein and DNA was analyzed by gel mobility shift assay (EMSA)invitro. Purification of recombinant protein Rex was successfully expressed. The presence of NAD+did not have major effect on mobility shift, but addition of NADH almost abolished such a binding activity. Byinvitrobinding assay, Rex was found to regulate the expression of Prex in response to NADH/NAD+equilibrium.
Streptococcussuis; Rex; transcriptional regulator
10.3969/j.issn.1002-2694.2017.08.003
國(guó)家轉(zhuǎn)基因生物新品種培育重大專項(xiàng)子課題(No.2014ZX0800946B-002);國(guó)家公益性行業(yè)科研專項(xiàng)(No.201303041)
何孔旺,Email:kwh2003@263.net
1.江蘇省農(nóng)業(yè)科學(xué)院獸醫(yī)研究所·農(nóng)業(yè)部獸用生物制品工程技術(shù)重點(diǎn)實(shí)驗(yàn)室,南京 210014; 2.南京農(nóng)業(yè)大學(xué)動(dòng)物醫(yī)學(xué)院,南京 210095; 3.江蘇省食品質(zhì)量安全重點(diǎn)實(shí)驗(yàn)室-省部共建國(guó)家重點(diǎn)實(shí)驗(yàn)室培育基地,南京 210014
Supported by the National Genetically Modified Organisms Breeding Major Projects of China(No.2014ZX0800946B-002)and the Special Fund for Agro-scientific Research in the Public Interest(No.201303041) Corresponding author: He Kong-wang, Email: kwh2003@263.net
R373.1
:A
:1002-2694(2017)08-0680-05
2016-11-25編輯:張智芳
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
