轉舞毒蛾LdCYP6AN15v1基因果蠅品系對氯蟲苯甲酰胺脅迫響應*
2017-07-18 12:10:13黨英僑殷晶晶陳傳佳孫麗麗曹傳旺
林業科學 2017年6期
黨英僑 殷晶晶 陳傳佳 孫麗麗 劉 鵬 曹傳旺
(東北林業大學林學院 哈爾濱 150040)
?
轉舞毒蛾LdCYP6AN15v1基因果蠅品系對氯蟲苯甲酰胺脅迫響應*
黨英僑 殷晶晶 陳傳佳 孫麗麗 劉 鵬 曹傳旺
(東北林業大學林學院 哈爾濱 150040)
【目的】 舞毒蛾是林業重要害蟲,細胞色素P450是昆蟲體內廣泛分布的參與外源化合物代謝關鍵酶系,探討P450家族基因CYP6AN15v1對殺蟲劑代謝解毒功能,為舞毒蛾有效治理提供依據。【方法】 通過RT-PCR法獲得LdCYP6AN15v1基因cDNA全長,采用傳統酶切連接的方法構建轉CYP6AN15v1基因果蠅載體,通過轉基因技術獲得表達LdCYP6AN15v1果蠅品系(命名為attP40>CYP6AN15v1)。采用分光光度計法研究低劑量氯蟲苯甲酰胺(7.17 mg·L-1)處理對轉基因和非轉基因果蠅品系細胞色素P450活性的影響,并采用qRT-PCR法測定其對CYP6AN15v1基因表達的影響。【結果】 從舞毒蛾無參照轉錄本文庫中克隆獲得CYP6AN15v1全長基因,編碼512個氨基酸,蛋白分子質量為59.02 kDa; 系統進化樹分析表明CYP6AN15v1與甜菜夜蛾和棉鈴蟲關系較近。以DNA和cDNA為模板,attP40>CYP6AN15v1果蠅品系均檢測到1 539 bp目的基因,表明LdCYP6AN15v1基因成功整合到果蠅基因組。與非轉基因attP40果蠅品系相比,轉基因attP40>CYP6AN15v1果蠅品系對氯蟲苯甲酰胺的敏感性顯著降低,致死中濃度LC50為非轉基因果蠅的2.92倍; 低劑量(7.17 mg·L-1)氯蟲苯甲酰胺脅迫下,舞毒蛾細胞色素P450酶活性和CYP6AN15v1基因的誘導作用呈現時間效應,attP40>CYP6AN15v1果蠅品系P450活性為非轉基因果蠅的1.09~1.93倍,主要表現為誘導效應; attP40>CYP6AN15v1果蠅品系的CYP6AN15v1基因mRNA表達量呈誘導激活,其表達量為非轉基因的44.54~137.80倍。【結論】 利用轉基因技術成功構建了轉LdCYP6AN15v1果蠅品系attP40>CYP6AN15v1; 氯蟲苯甲酰胺可能通過誘導LdCYP6AN15v1基因mRNA的上調表達而增強黑腹果蠅P450酶活性,從而參與對氯蟲苯甲酰胺的解毒作用。關鍵詞: 舞毒蛾; 轉基因果蠅; 氯蟲苯甲酰胺;CYP6AN15v1; 誘導表達
Responses of TransformantDrosophilaExpressingLdCYP6AN15v1 Gene to
舞毒蛾(Lymantriadispar)是分布很廣的世界性害蟲,國外報道可危害300多種植物(Leonard, 1981),國內報道可危害500多種植物,分布于東北、華北、西北、華東、華中、華南、西南等地區(侯雅芹等, 2009; 許娜等, 2009)。目前,使用廣泛、殺蟲效果好、作用迅速的方法是利用氯氰菊酯、高效氯氰菊酯、溴氰菊酯、氧化樂果等化學殺蟲劑進行防治,但這些殺蟲劑易產生“3R”問題(侯雅芹等, 2009; 倪鳴等, 2009),易對環境造成嚴重的危害。以魚尼丁受體為靶標的雙酰胺類殺蟲劑的出現,改變了傳統農藥高毒性、高殘留、環境不友好的局面(劉熠等, 2009)。
雙酰胺類殺蟲劑與魚尼丁受體結合后,會刺激胞內Ca2+從肌肉細胞的肌質網釋放和消耗,使昆蟲肌肉細胞的收縮功能癱瘓(Cordovaetal., 2006),而魚丁尼受體則是調節Ca2+釋放的選擇性通道。氯蟲苯甲酰胺是美國杜邦公司2008年推出的一種新型鄰甲酰氨基苯甲酰胺類殺蟲劑,用于各種作物對鱗翅目(董波等, 2009; 陳偉國等, 2010)、鞘翅目(Bassietal., 2008; 2009; Koppenh?feretal., 2008; Kuharetal., 2008)、雙翅目(Teixeiraetal., 2009)、等翅目(Yeohetal., 2007; Spomeretal., 2009)、半翅目(Dhawanetal., 2009; Lahmetal., 2009)等害蟲的防治,而對非靶標節肢動物具有良好的選擇性,且對哺乳動物、鳥類、魚等的毒性低,此外也有研究表明氯蟲苯甲酰胺對一些鱗翅目的害蟲有顯著殺卵作用(Ioriattietal., 2009; Lahmetal., 2009),在亞致死濃度處理下,會影響雌性蘋果蠹蛾(Cydiapomonella)的交配比例(Knightetal., 2007)。
細胞色素P450(cytochrome P450)是參與殺蟲劑代謝重要解毒酶系之一,在殺蟲劑的氧化、還原等化學反應及殺蟲劑抗性的形成中發揮重要作用(Zhouetal., 2010),其中昆蟲已有超過1 000個細胞色素P450基因被確定(Nelson, 2009),主要分布在CYP4、CYP6、CYP9、CYP28、CYP321和CYP12家族(Feyereisen, 2005),常參與殺蟲劑代謝和抗性相關的P450基因分布于CYP4、CYP6、CYP9、CYP12亞家族中(Feyereisen, 2006; Lietal., 2007)。大量研究表明, 細胞色素P450過表達是昆蟲產生抗藥性的重要機制之一(Liuetal., 1998; Lietal., 2006)。Bautista等(2007)研究發現氯菊酯小菜蛾(Plutellaxylostella)抗性品系中CYP6BG1和CYP6BG2的轉錄水平較高。已有研究表明CYP321E1參與了小菜蛾對氯蟲苯甲酰胺的抗性形成(Huetal., 2014)。筆者實驗室前期研究發現舞毒蛾CYP6AN15v1基因在殺蟲劑溴氰菊酯、甲萘威、氧化樂果(15 mg·L-1)脅迫下主要以下調表達為主(Sunetal., 2014); 并采用RNA干擾(RNAi)技術通過體外合成CYP6AN15v1的dsRNA顯微注射至舞毒蛾3齡幼蟲體內,發現dsRNA可以在特定時間內高效特異的沉默舞毒蛾體內CYP6AN15v1基因的mRNA表達,且舞毒蛾幼蟲對有機磷類殺蟲劑的敏感性顯著提高(曹傳旺等, 2015)。然而,RNAi不能穩定遺傳,只能達到瞬時干擾基因表達的目的,轉基因昆蟲技術是在昆蟲基因組中插入具有特定的基因,分析這些基因功能。黑腹果蠅(Drosophilamelanogaster)作為重要的模式生物,其所具有的轉基因技術優勢,促進了生物學諸多領域的研究進展(周秀娟等, 2008)。目前,已有昆蟲學者將與殺蟲劑作用相關基因轉入黑腹果蠅中,以驗證其參與殺蟲劑代謝(孫麗麗等,2016; 薛緒亭等, 2016)。Riveron等(2013; 2014)借助于GAL4/UAS系統將按蚊(Anophelesfunestus)CYP6P9a、CYP6P9b和CYP6M7基因分別表達于果蠅體內,證實其參與氯菊酯和溴氰菊酯的抗性形成。本文在分析舞毒蛾轉錄本文庫的基礎上,克隆分析了CYP6AN15v1基因的cDNA全長,并通過轉基因技術研究CYP6AN15v1在舞毒蛾對氯蟲苯甲酰胺類殺蟲劑的脅迫響應,旨在深入探索舞毒蛾CYP6AN15v1參與魚尼丁受體抑制劑類殺蟲劑抗藥性形成機制以及為在生產實踐中進行抗藥性監測和治理提供理論依據。
1 材料與方法
1.1 供試昆蟲
供試舞毒蛾卵塊和人工飼料均購自于中國林業科學研究院森林生態環境與保護研究所,放置于溫度(25±1)℃、光照14 L∶10D、相對濕度75%的條件下人工飼養。
黑腹果蠅來源于中國科學院上海生命科學研究院生物化學與細胞生物學研究所果蠅資源與技術平臺,置于(25±1)℃、相對濕度60%、光照14 L∶10D人工養蟲室內飼養。所用人工飼料主要成分為蔗糖10 g,玉米粉10 g,酵母粉0.5 g,瓊脂1 g,100 mL蒸餾水,丙酸0.7 mL,每4天將蟲子換入新飼料瓶中,保持瓶內空氣流通。
黑腹果蠅品系: attP40果蠅品系(即25C)基因型為y[1] M {vas-int.Dm}ZH-2A w[*], P{CaryP}attP40; 雜交果蠅品系為ywR13 s=yw, sp/Cyo, MKRS/TM2。
1.2 主要試劑
考馬斯亮藍G-250、牛血清白蛋白(BSA)、苯甲基磺酰氟(PMSF)、二硫蘇糖醇(DTT)、乙二胺四乙酸(EDTA)、丙基硫氧嘧啶(PTU)、還原性輔酶Ⅱ(NADPH)和二甲基亞砜(DMSO)均購自于SIGMA公司; 磷酸二氫鈉、磷酸氫二鈉、濃鹽酸、氯仿、乙醇、磷酸、氫氧化鈉、丙酮均為分析純級,購自天津永大試劑有限公司。氯蟲苯甲酰胺原藥(96%,國家生物農藥工程中心);XhoI和XbaI限制性內切酶、T4 DNA連接酶、DNase I均購自于Promega公司; RNA提取試劑盒購自Qiagen公司; pMD18-T載體和PrimeScriptTMRT reagent Kit cDNA合成試劑盒購自TaKaRa公司,SYBR Green Real-time PCR Master mix購自Toyobo公司,西班牙瓊脂糖購自Gene公司; pUAST-attB載體為美國奧本大學劉喃喃教授贈予; 大腸桿菌(Escheriacoli)DH5α為本實驗室保存。
1.3LdCYP6AN15v1基因克隆與分析
根據舞毒蛾幼蟲cDNA文庫獲得的舞毒蛾CYP6AN15v1基因序列(命名為LdCYP6AN15v1)。設計全長cDNA引物進行PCR,切膠回收后連接于pMD18-T 載體上,轉化至大腸桿菌DH5α,在含氨芐抗性的平板上挑取重組克隆,獲得的陽性菌株,進一步進行測序分析,最終確定所獲得的LdCYP6AN15v1基因cDNA序列,并將此重組載體記為CYP6AN15v1/pMD18-T。
應用ORF founder(http://www.ncbi.nlm.nih.gov/gorf.html)程序確定其開放讀碼框。用NCBI的Conserved Domains程序(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)預測蛋白的保守區,并檢查蛋白質序列的結構域及功能基序; 利用Blast 程序(http://www.ncbi.nlm.nih.gov/BLAST/)進行序列同源性搜索,選擇與其相似程度高的其他昆蟲GPCR蛋白氨基酸序列,用多序列聯配程序Clustalx(1.83)進行多序列比對。應用Clustalx(1.83)和MEGA(5.1),采用鄰接法(Neighbor-Joining, N-J)構建系統發育樹,
1.4 果蠅表達載體構建及轉LdCYP6AN15v1果蠅品系的建立
分別用限制性內切酶XhoI和XbaI酶切CYP6AN15v1/pMD18-T質粒和 pUAST-attB載體質粒,酶切產物進行1.5%瓊脂糖凝膠電泳檢測后,分別回收LdCYP6AN15v1基因片段和pUAST-attB載體,通過T4 DNA連接酶,16 ℃連接16 h,轉化至大腸桿菌DH5α,在含氨芐抗性的平板上挑取重組克隆,經質粒PCR和雙酶切消化后的陽性克隆,送至北京六合華大生物技術科技有限公司測序驗證,成功構建的重組質粒命名為pUAST-attB-CYP6AN15v1。
轉基因果蠅定點注射委托中國科學院上海生命科學研究院生物化學與細胞生物學研究所果蠅資源與技術平臺完成,具體步驟如下: 將重組質粒經Qiagen質粒提取試劑盒去內毒素抽提后,注射至帶attp背景的親本果蠅胚胎中,定點微量注射入2#染色體; y[1] M {vas-int.Dm}ZH-2A w[*], P{CaryP}attP40] G0代果蠅胚胎中,羽化后的G0代果蠅雄蠅與yw, sp/Cyo, MKRS/TM2處女蠅單雜交(或者羽化后的G0代果蠅處女蠅與yw, sp/Cyo, MKRS/TM2雄單雜交); 約2周后,檢查這些果蠅的G1代,從中挑選眼睛為紅色或橘黃色的果蠅,可初步認定為轉基因果蠅。挑取帶balancer紅眼(或橘黃色)雄果蠅,再與yw, sp/Cyo, MKRS/TM2處女蠅雜交; 挑選帶Cyo的轉基因紅眼果蠅自交,雜合或純合保種,查紅眼獲得穩定的轉基因果蠅品系,命名為attP40>CYP6AN15v1,目的基因均以單拷貝插入到果蠅的2#染色體上。
1.5 轉LdCYP6AN15v1果蠅品系分子驗證
采用Omega 試劑盒提取attP40>CYP6AN15v1、attP40(25C)果蠅品系DNA,以DNA為模板,用全長特異性引物進行RT-PCR擴增(引物見表1)。采用RNeasy Mini動物組織總RNA提取試劑盒(Qiagen)提取attP40>CYP6AN15v1、attP40(25C)果蠅品系總RNA,利用反轉錄試劑盒PrimeScriptTMRT reagent Kit(TaKaRa)合成cDNA,以cDNA為模板用全長特異性引物進行RT-PCR擴增(引物見表1)。反應體系如下: 10×LA Buffer 1 μL,正向和反向引物各0.5 μL,dNTP 0.2 μL,cDNA(或DNA)1.0 μL,LATaq酶 0.2 μL,ddH2O補足至10 μL。PCR擴增程序為: 94 ℃預變性 3 min; 94 ℃變性 30 s,60 ℃退火 30 s,72 ℃ 延伸2 min,35個循環; 72 ℃ 延伸10 min。1.0%瓊脂糖凝膠電泳檢測目的基因擴增產物。

表1 本文中所用的引物Tab. 1 Primer sequences used in this experiment
1.6 轉LdCYP6AN15v1果蠅致毒處理
參考王曉軍等(2006)和薛緒亭等(2016)方法采用藥膜法生物測定,取200 μL不同濃度的氯蟲苯甲酰胺藥液在閃爍瓶中形成均勻藥膜,挑取30頭不同品系果蠅放入瓶中,以G0代果蠅品系為對照,每個處理3次重復,于24 h統計死亡率,采用POLO軟件,計算致死中濃度LC50。
取200 μL氯蟲苯甲酰胺(LC20=7.17 mg·L-1)藥液于閃爍瓶中,水平轉動形成均勻藥膜,加入少許的人工飼料(約1 cm3)。分別取180頭大小、顏色一致的對照和轉基因果蠅成蟲置于處理的閃爍瓶中,以丙酮處理為對照組,每個處理30頭,6個重復,分別于6,12,24,48和72 h挑取20頭活潑的果蠅放于離心管中,液氮速凍后,置于-80 ℃冰箱內,分別用于實時熒光RT-PCR和細胞色素P450酶活性測定。1.7 細胞色素P450活性測定
1.7.1 酶液制備 從超低溫冰箱內依次取出不同時間點處理的果蠅品系,以冷凍狀態輕敲裝有果蠅成蟲的離心管,隨機挑選10頭快速倒至5 mL玻璃勻漿器內,加入4.0 mL 0.1 mol·L-1pH 7.5的磷酸緩沖液(含0.1 mmol·L-1DTT、1 mmol·L-1EDTA、1 mmol·L-1PTU、1 mmol·L-1PMSF),冰上研磨充分,12 000 g離心15 min,上清液即為原酶液。蛋白質含量測定參照Bradford(1976)的考馬斯亮藍G-250法。
1.7.2 細胞色素P450活性測定 將1 mL原酶液、800 μL 0.1 mol·L-1pH 7.5磷酸緩沖液、100 μL 4.8 mmol·L-1NADPH,100 μL 20 mmol·L-1對硝基苯甲醚底物放入10 mL離心管中充分振蕩混勻,30 ℃水浴溫育30 min,加入40 μL濃鹽酸終止反應,再加入5 mL氯仿,劇烈振蕩20 min,靜置3~5 min,4 200 g,4 ℃離心15 min,吸取下層清液3 mL并轉移至10 mL離心管中,加入3 mL 0.5 mol·L-1氫氧化鈉溶液,劇烈震蕩15 min,靜置5 min,吸取上層萃取液為待測液。空白對照組采用3 mL氫氧化鈉溶液,每個處理3次重復,于412 nm測定生成對硝基苯酚OD值,再根據對硝基苯酚標準曲線求出對硝基苯酚含量。以3 mL 0.5 mol·L-1氫氧化鈉溶液中加入不同濃度對硝基苯酚,劇烈震蕩15 min,靜置5 min,吸取上層萃取液測定412 nm下OD值,根據OD值和對硝基苯酚濃度制作標準曲線。根據反應時間、對硝基苯酚的生成量和蛋白質含量計算細胞色素P450比活力(nmol·min-1mg-1)。
1.8 實時熒光定量RT-PCR


圖1 舞毒蛾LdCYP6AN15v1基因的cDNA及由此推導的氨基酸序列Fig.1 The cDNA and deduced amino acid sequence of LdCYP6AN15v1 in L. dispar
采用RNeasy Mini動物組織總RNA提取試劑盒提取對照(丙酮)、氯蟲苯甲酰胺(LC20=7.17 mg·L-1)處理的果蠅成蟲總RNA,采用PrimeScriptTMRT reagent Kit cDNA合成試劑盒合成cDNA,將此cDNA模板稀釋10倍,作為實時熒光定量RT-PCR模板。實時熒光定量RT-PCR使用試劑盒SYBR Green Real-time PCR Master mix,內參為RpL32、ATP基因(Willisetal., 2010),引物序列見表1。實時熒光定量RT-PCR反應體系為: 10 μL 2×SYBR premix ExTaq酶、正向和反向引物(10 μmol·L-1)各1,2 μL稀釋后cDNA模板,加去離子水補至20 μL。實時熒光定量PCR反應條件為: 94 ℃預變性30 s,94 ℃變性12 s,60 ℃退火30 s,72 ℃延伸40 s,81 ℃讀板1 s,45個循環,每個樣品重復3次,用2-△△Ct方法進行基因的相對定量分析(Pfaffletal., 2002)。1.9 數據統計分析
運用SPSS17.0軟件,采用Duncan方法對氯蟲苯甲酰胺脅迫下不同處理時間P450酶活性和LdCYP6AN15v1基因轉錄水平進行顯著性差異分析(P< 0.05)。
2 結果與分析
2.1LdCYP6AN15v1基因分析
獲得的舞毒蛾LdCYP6AN15v1基因的閱讀框(ORF)長1 539 bp,編碼512個氨基酸(圖1)。用SignalP4.1軟件對LdCYP6AN15v1氨基酸序列前70個氨基酸殘基進行信號肽預測。預測結果如圖2 所示,在信號肽切割位點位于第17~18個氨基酸之間即VYF-TR間,表明這種蛋白是分泌型蛋白。采用NCBI的CDD軟件分析LdCYP6AN15v1蛋白的保守結構域表明該蛋白屬于P450家族。

圖2 舞毒蛾LdCYP6AN15v1基因的保守結構域Fig.2 The cDNA and deduced amino acid sequence of LdCYP6AN15v1 in L. dispar
通過BLASTP多序列比對,選擇與舞毒蛾LdCYP6AN15v1蛋白相似程度極高的16種昆蟲P450蛋白進行多序列比對,構建的16種昆蟲P450基因系統進化樹表明P450蛋白聚為2大類,其中舞毒蛾CYP6AN15v1蛋白與甜菜夜蛾(Spodopteralittoralis)和棉鈴蟲(Helicoverpaarmigera)親緣關系近而聚為一類(圖3)。
2.2 轉LdCYP6AN15v1基因果蠅載體構建
將LdCYP6AN15v1基因連接至pUAST-attB載體,獲得重組轉基因質粒pUAST-attB-CYP6AN15v1,重組質粒分別經XhoI和XbaI雙酶切檢測,可見1 539 bp目的基因條帶。將重組質粒進行測序,其結果與目的基因序列完全一致,沒有序列發生缺失或突變,表明pUAST-attB-CYP6AN15v1載體構建成功。
2.3 轉LdCYP6AN15v1基因果蠅驗證
圖4顯示表達LdCYP6AN15v1基因果蠅純合品系表現型。提取表征為紅眼的轉基因果蠅總RNA,反轉錄合成cDNA,以cDNA為模板,引物見表1,PCR檢測CYP6AN15v1基因,結果顯示轉基因果蠅品系中CYP6AN15v1基因得以表達。以DNA為模板進行PCR擴增時,attP40>CYP6AN15v1果蠅品系的PCR產物片段大小為1 539 bp(圖5),非轉基因attP40果蠅品系未見目的條帶,這表明舞毒蛾的CYP6AN15v1基因成功整合到黑腹果蠅的基因組中。
2.4 氯蟲苯甲酰胺對轉LdCYP6AN15v1基因果蠅毒力
與非轉基因attP40果蠅品系相比,表達LdCYP6AN15v1果蠅品系attP40>CYP6AN15v1對氯蟲苯甲酰胺的敏感性增加,LC50值高于親本,為對照親本的2.92倍(表2)。
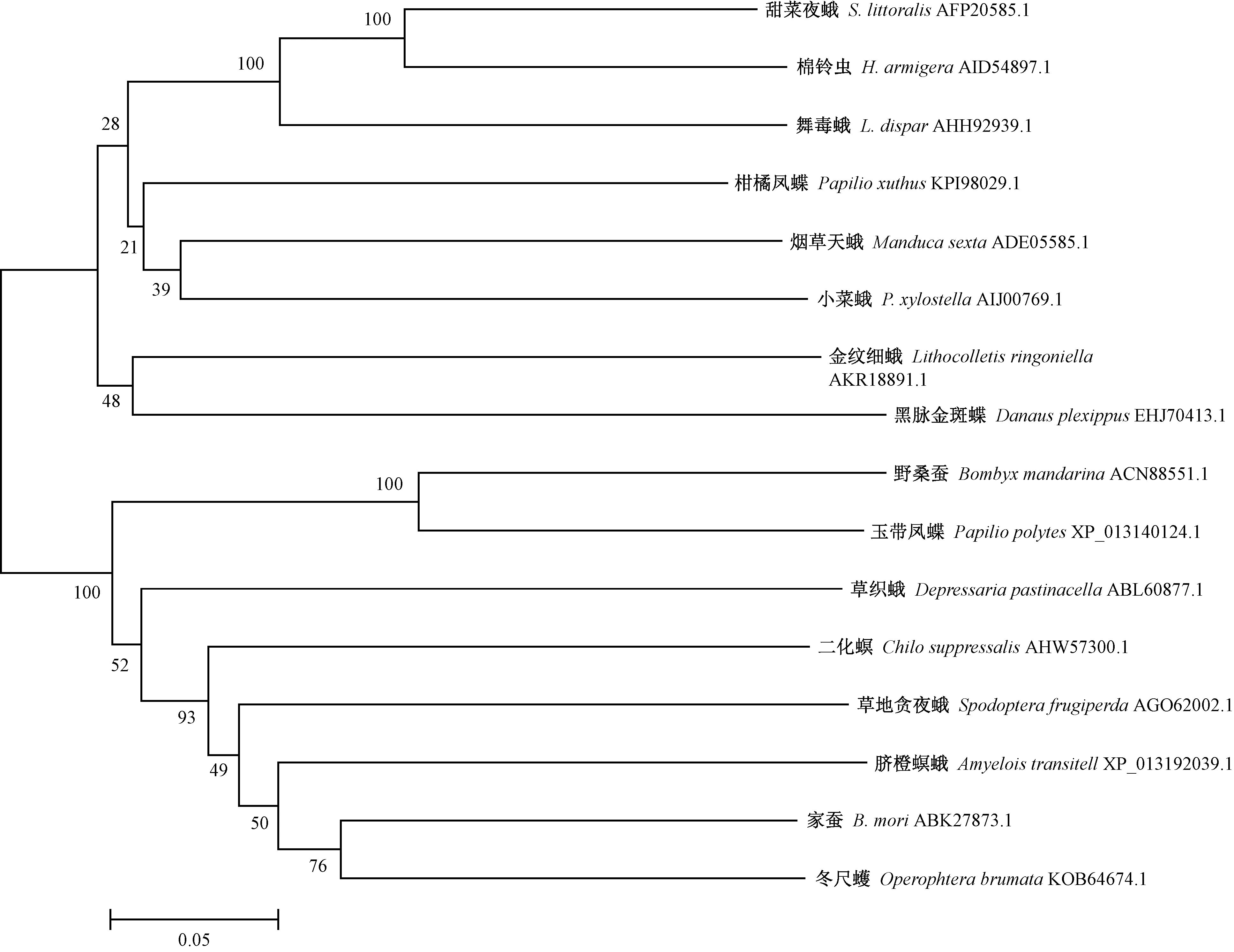
圖3 16種昆蟲P450蛋白系統進化樹Fig.3 Phylogenetic analysis of P450 proteins from 16 insects

圖4 轉LdCYP6AN15v1基因和非轉基因黑腹果蠅品系表現型Fig.4 Phenotype of transformant and untransformant D.melanogaster A: 轉基因果蠅attP40>CYP6AN15v1 Transformant D.melanogaster attP40>CYP6AN15v1; B: 非轉基因果蠅attP40 Untransformant D.melanogaster attP40.
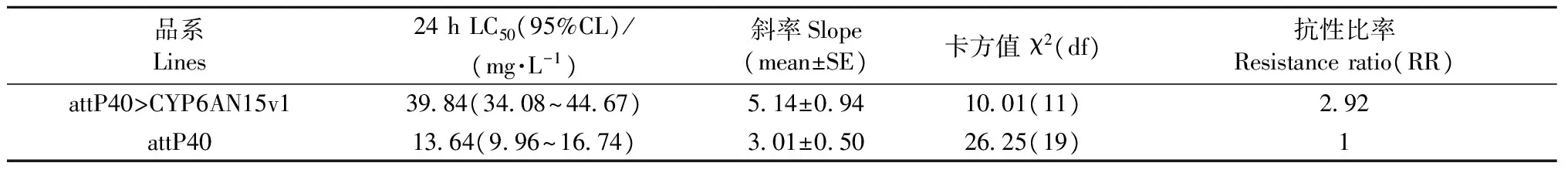
品系Lines24hLC50(95%CL)/(mg·L-1)斜率Slope(mean±SE)卡方值χ2(df)抗性比率Resistanceratio(RR)attP40>CYP6AN15v13984(3408~4467)514±0941001(11)292attP401364(996~1674)301±0502625(19)1
① χ2(11,0.05)=19.68; χ2(19,0.05)=30.14。
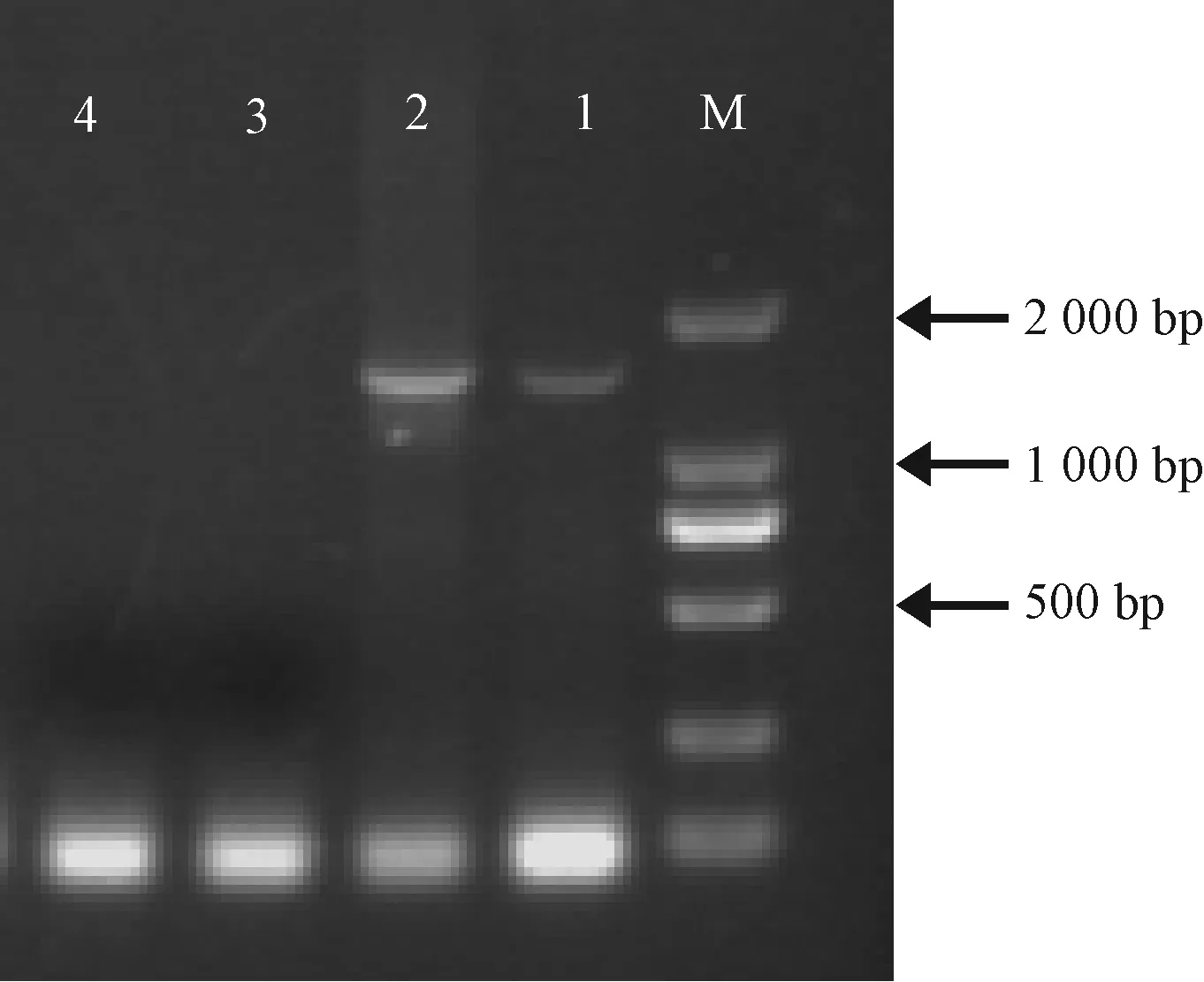
圖5 轉基因果蠅品系LdCYP6AN15v1 基因PCR檢測Fig.5 PCR analysis of LdCYP6AN15v1 in transformant D.melanogaster 1: attP40>CYP6AN15v1果蠅品系DNA為模板DNA of attP40>CYP6AN15v1 D.melanogaster line as template; 2: attP40>CYP6AN15v1果蠅品系cDNA為模板cDNA of attP40>CYP6AN15v1 D.melanogaster line as template; 3: attP40果蠅品系DNA為模板 DNA of attP40 D.melanogaster line as template; 4: attP40果蠅品系cDNA為模板 cDNA of attP40 D.melanogaster line as template.
2.5 氯蟲苯甲酰胺對轉LdCYP6AN15v1果蠅P450活性影響
氯蟲苯甲酰胺(7.17 mg·L-1)脅迫下轉基因果蠅和非轉基因果蠅品系體內的P450活性結果顯示,除12 h外,其余各時間點轉基因果蠅品系P450比活力均高于對照果蠅品系。處理初期0 ~ 12 h, attP40>CYP6AN15v1果蠅品系的總酶活性與對照組attP40的相比,均無顯著性差異(P> 0.05)(圖6),隨脅迫時間的延長(24 ~ 72 h),attP40>CYP6AN15v1果蠅品系的P450活性顯著高于對照組attP40品系(P< 0.05); 脅迫72 h,轉基因果蠅品系的P450活性顯著高于對照組,活性增加93.08%,為未處理轉基因果蠅品系的2.55倍。

圖6 氯蟲苯甲酰胺對黑腹果蠅品系P450活性影響Fig.6 Effects of chlorantraniliprole on cytochrome P450 activities in D.melanogaster *表示同一時間轉基因和對照組果蠅品系P450活性差異顯著(P<0.05),下同。 * indicates the P450 activities between transgenic- and control-D.melanogaster are significant at the same time at 0.05 level. The same below.
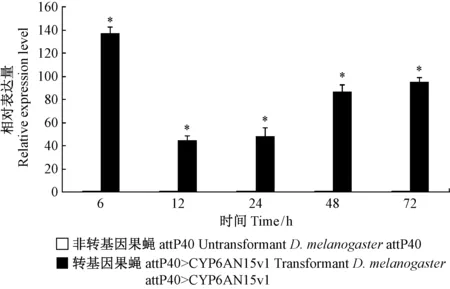
圖7 氯蟲苯甲酰胺對黑腹果蠅品系CYP6AN15v1表達量影響Fig.7 Effects of chlorantraniliprole on CYP6AN15v1 gene expression in transformant D.melanogaster
2.6 氯蟲苯甲酰胺對轉基因果蠅CYP6AN15v1表達量影響
采用qRT-PCR法測定了氯蟲苯甲酰胺(7.17 mg·L-1)脅迫下轉基因和非轉基因果蠅品系CYP6AN15v1的轉錄水平影響。氯蟲苯甲酰胺顯著誘導了attP40>CYP6AN15v1果蠅品系體內CYP6AN15v1的表達(P<0.05),相對表達量為對照組44.54~137.80倍; 其中,12 hCYP6AN15v1的誘導表達水平最低,為對照組的44.54倍(圖7)。
3 討論
昆蟲細胞色素P450兼具單加氧酶和氧化酶的功能,參與許多生物體內內源性化合物(如昆蟲激素、脂肪酸和甾醇等)的生物合成(Lietal., 2007),也具有對外源性化合物的催化代謝能力(Lvaylo, 2001; Zhaoetal., 2002; Lietal., 2004),在昆蟲的生長發育和對環境的適應性方面具有重要的作用(Feyereisen, 2012)。昆蟲產生抗藥性的主要機制是P450介導的殺蟲劑代謝解毒作用,P450增強殺蟲劑代謝解毒作用一是與酶量增加相關,二是由于酶結構變化導致解毒活性的增強。細胞色素P450表達量的增加主要有2種機制: 一是基因組層面上的基因拷貝數增加,二是基因轉錄水平的基因上調表達,其中第2種機制更為普遍(邱星輝, 2014)。已有研究證實P450參與了對有機磷類、擬除蟲菊酯類、新煙堿類、氨基甲酸酯類等化學合成殺蟲劑以及天然的植物防御化合物的代謝解毒(Guengerich, 2004)。Chiu等(2008)通過原核表達CYP6Z1蛋白具有代謝DDT的作用,證實該基因的上調表達參與了岡比亞按蚊(Anophelesgambiae)對DDT的抗性; Zhu等(2010)報道赤擬谷盜(Triboliumcastaneum)QTC279品系對溴氰菊酯的抗性是源于腦部特異性過量表達CYP6BQ9。
本文利用模式動物果蠅轉基因技術研究了舞毒蛾CYP6AN15v1基因對氯蟲苯甲酰胺誘導解毒功能,結果表明氯蟲苯甲酰胺脅迫下轉基因果蠅品系CYP6AN15v1基因表達和P450活性均顯著高于非轉基因果蠅品系,且轉CYP6AN15v1基因果蠅品系對氯蟲苯甲酰胺的敏感性降低,這可能是由于CYP6AN15v1基因誘導表達增加解毒能力導致的。曹傳旺等(2015)利用RNAi技術體外合成CYP6AN15v1 dsRNA微量注射入經饑餓處理12 h舞毒蛾3齡幼蟲體內,并將沉默后的幼蟲置于混有40 mg·L-1的氧化樂果飼料中,48 h時試驗組舞毒蛾幼蟲死亡率為60.00%,比對照組ddH2O、GFPdsRNA分別高出26.67%和40.00%,表現出對氧化樂果高敏感性,這表明CYP6AN15v1基因表達量減少降低了細胞色素P450對氧化樂果的解毒能力。
近些年來,已有研究報道了細胞色素P450家族基因參與氯蟲苯甲酰胺解毒代謝。邢靜等(2011)采用亞致死劑量(LC10和LC25)氯蟲苯甲酰胺連續處理小菜蛾5代后,發現細胞色素P450 O-脫乙基(ECOD)的比活力顯著高于對照,為對照組的1.65和1.56倍; Liu等(2015)篩選52代氯蟲苯甲酰胺抗性品系小菜蛾P450酶活性是敏感品系的4.26倍。氯蟲苯甲酰胺抗性品系和敏感品系小菜蛾轉錄組中,發現19個P450、3個GST解毒酶基因和魚丁尼受體基因均上調表達(Lin等, 2013)。歐善生等(2012)研究表明亞致死劑量氯蟲苯甲酰胺(LC10、LC25和LC50)處理棉鈴蟲3齡幼蟲時,P450-ECOD比活力為LC10> LC25> LC50。王學貴等(2015)研究發現以0.02 mg·kg-1氯蟲苯甲酰胺處理甜菜夜蛾,3種P450基因(CYP9A9、CYP4G37、CYP6B)和細胞色素P450還原酶基因HQ852049 mRNA上調表達以及P450酶活表達增加,其中中腸的P450基因CYP9A9、CYP4G37、CYP6B的相對表達量為對照的1.53 ~ 4.48倍,而處理組P450酶活性為對照組1.90 ~ 2.92倍。這些研究結果表明雙酰胺類氯蟲苯甲酰胺殺蟲劑作用靶標為魚尼丁受體外,眾多細胞色素P450家族基因參與了抗性形成和誘導代謝。
4 結論
利用轉基因技術成功構建了轉舞毒蛾P450基因CYP6AN15v1的果蠅品系attP40>CYP6AN15v1; 明確了氯蟲苯甲酰胺可能通過誘導CYP6AN15v1基因mRNA的上調表達而增加了黑腹果蠅細胞色素P450酶活性,轉舞毒蛾CYP6AN15v1果蠅對氯蟲苯甲酰胺敏感性降低可能源于P450活性的增加解毒能力增強導致的。有關舞毒蛾CYP6AN15v1表達P450蛋白對氯蟲苯甲酰胺解毒機制還有待于進一步研究。
曹傳旺, 高彩球, 孫麗麗, 等. 2015. 舞毒蛾CYP6AN15v1基因dsRNA及其在無公害防治中的應用. 申請號: 201510054278.1
(Cao C W, Gao C Q, Sun L L,etal. 2015. dsRNA ofCYP6AN15v1 inLymantriadisparand its application in friendly control. Application No. 201510054278.1.
陳偉國, 董瑞華, 孫海燕, 等. 2010. 農用殺蟲劑氯蟲苯甲酰胺對家蠶的毒性研究. 蠶業科學, 36(1):84-90.
(Chen W G, Dong R H, Sun H Y,etal. 2010. An investigation on toxicity of the agricultural pesticide chlorantraniliprole to the silkworm,Bombyxmori. Science of Sericulture, 36(1): 84-90. [in Chinese])
董 波, 孫國俊, 蔣林忠, 等. 2009. 茚蟲威15%乳油和氯蟲苯甲酰胺20%懸浮劑對稻縱卷葉螟的防效試驗. 農藥科學與管理, 30(3):31-32.
(Dong B, Sun G J,Jiang L Z,etal. 2009. Effect tests of indoxacarb and chlorantraniliprole onCnaphalocrocismedialisGuenee. Pesticide Science and Administration, 30(3): 31-32. [in Chinese])
侯雅芹, 南 楠, 李鎮宇. 2009. 舞毒蛾研究進展. 河北林果研究, 24(4):439-444.
(Hou Y Q, Nan N, Li Z Y. 2009.Research advances ofLymantriadispar. Hebei Journal of Forestry and Orchard Research, 24(4):439-444. [in Chinese])
劉 熠, 王國勝. 2009. 魚尼丁受體類新型殺蟲劑氯蟲酰胺的研究概述.化學工程師, 171(11):44-47.
(Liu Y, Wang G S. 2009. Summary of chlorantraniliprole—a new type of ryanodines receptor insecticide. Chemical Engineer, 171(11):44-47. [in Chinese])
倪 鳴, 柴汝松, 張 鵬, 等. 2009. 不同藥劑對舞毒蛾和春尺蠖的毒力測定. 東北林業大學學報, 37(5):121-122.
(Ni M, Chai R S, Zhang P,etal. 2009. Toxicity test of different kinds of insecticides toLymantriadisparandApocheimacinerarius. Journal of Northeast Forestry University, 37(5):121-122. [in Chinese])
歐善生, 梁 沛, 宋敦倫, 等. 2012. 氯蟲苯甲酰胺亞致死劑量對棉鈴蟲生長發育和解毒酶活性的影響. 植物保護, 38(4):1-8.
(Ou S S, Liang P, Song D L,etal. 2012. Effects of sublethal dosage of chlorantraniliprole on development and detoxifying enzymes activity ofHelicoverpaarmigera. Plant Protection, 38(4):1-8. [in Chinese])
邱星輝. 2014. 細胞色素 P450介導的昆蟲抗藥性的分子機制. 昆蟲學報, 57(4):477-482.
(Qiu X H. 2014. Molecular mechanisms of insecticide resistance mediated by cytochrome P450s in insects. Acta Entomologica Sinica, 57(4):477-482. [in Chinese])
孫麗麗,劉 鵬,王志英,等.2016.轉LdOA1基因果蠅品系GSTs基因表達及對溴氰菊脂脅迫響應.北京林業大學學報,38(6):72-78.
(Sun L L, Liu P, Wang Z Y,etal. 2016. Glutathione S-transferase gene expression in transformantDrosophilaexpressing ocular albinism type 1 from gypsy moth and its response to deltamethrin stress.Journal of Beijing Forestry University,38(6):72-87.[in Chinese])
王曉軍, 張宗儉, 姜 輝, 等. 2006. NY/T1154.7-2006農藥室內生物測定試驗準則. 北京: 中國標準出版社.
(Wang X J, Zhang Z J, Jiang H,etal. 2006. NY/T1154.7-2006 Guidelines for indoor bioassay of pesticide. Beijing: China Standard Press.[in Chinese])
王學貴, 余慧靈, 梁 沛, 等. 2015. 氯蟲苯甲酰胺誘導甜菜夜蛾細胞色素P450 基因上調表達. 昆蟲學報, 58(3): 281-287.
(Wang X G, Yu H L, Liang P,etal. 2015. Chlorantraniliprole induces up-regulated expression of cytochrome P450 genes inSpodopteraexigua(Lepidoptera: Noctuidae). Acta Entomologica Sinica, 58(3): 281-287. [in Chinese])
邢 靜, 梁 沛, 高希武. 2011. 亞致死濃度氯蟲苯甲酰胺對小菜蛾藥劑的敏感度和解毒酶活性的影響. 農藥學學報, 13(5): 467-470.
(Xing J, Liang P, Gao X W,etal. 2011. Effects of sublethal concentrations of chlorantraniliprole on insecticide susceptibility and detoxifying enzyme activity inPlutellaxylostella. Chinese Journal of Pesticide Science, 13(5): 464-470. [in Chinese])
許 娜, 孫寶麗. 2009. 舞毒蛾的發生及防治. 現代農業科技, 17:172-173.
(Xu N,Sun B L. 2009. Occurrence and control ofLymantriadispar. Modern Agricultural Science and Technology, 17:172-173. [in Chinese])
薛緒亭,孫麗麗,劉 鵬,等.2016.表達LdCYP6B53果蠅品系建立及對殺蟲劑的敏感性.中國農學通報,32(19):97-101.
(Xue X T,Sun L L, Liu P,etal. 2016. Construction of transgenicDrosophilaexpressingLdCYP6B53 and its susceptibility to insecticides.Chinese Agricultural Science Bulletin,32(19):97-101.[in Chinese])
周秀娟,牛長纓,雷朝亮. 2008. 轉基因昆蟲的應用研究進展. 昆蟲知識,45(3): 368-373.
(Zhou X J, Niu C Y, Lei C L.2008. The application of transgenic insect technology. Chinese Bulletin of Entomology, 45(3):368-373.)
Bassi A, Molnar I, Zielinski D,etal. 2008. Chlorantraniliprole (Rynaxypyr?, Coragen?): a novel anthranilic diamide insecticide with outstanding control of Colorado Potato Beetle (Leptinotarsadecemlineata)∥Proceedings of the 17th Triennial Conference of the EAPR-International Potato Conference, 6-10, July 2008, Brasov, Romania.
Bassi A, Rison J L, Wiles J A. 2009. Chlorantraniliprole (DPX-E2Y45, Rynaxypyr?, Coragen?), a new diamide insecticide for control of codling moth (Cydiapomonella), Colorado potato beetle (Leptinotarsadecemlineata) and European grapevine moth (Lobesiabotrana). Zbornik predavanj in referatov 9 slovenskega posvetovanja o varstvu rastlin z mednarodno udele?bo.
Bautista M A M, Tanaka T, Miyata T. 2007. Identification of permethrin-inducible cytochrome P450s from the diamondback moth,Plutellaxylostella(L.) and the possibility of involvement in permethrin resistance. Pesticide Biochemistry and Physiology, 87(1): 85-93.
Chiu T L, Wen Z, Rupasinghe S G,etal. 2008. Comparative molecular modeling ofAnophelesgambiaeCYP6Z1, a mosquito P450 capable of metabolizing DDT. Proceeding of the National Academy of Sciences USA, 105(26): 8855-8860.
Cordova D, Benner E A, Sacher M D,etal. 2006. Anthranilic diamides: a new class of insecticides with a novel mode of action, ryanodine receptor activation. Pesticide Biochemistry and Physiology, 84: 196-214.
Dhawan A, Saini S, Singh K,etal. 2009. Persistence and residual toxicity of some insecticides againstPhenacoccussolenopsison cotton (Gossypiumspp.). Indian Journal of Agricultural Sciences, 79(3): 203-206.
Feyereisen R. 2005. Insect cytochrome P450 in comprehensive molecular insect science∥Gilbert L I, Iatrou K, Gill S S. Comprehensive Molecular Insect Science.Elsevier, Oxford, UK, 1-77.
Feyereisen R. 2006. Evolution of insect P450. Biochemical Society Transactions, 34(6): 1252-1255.
Feyereisen R, Jindra M. 2012. The silkworm coming of age-early. PLoS Genetics, 8(3):e1002591.
Guengerich F R, Krauser J A, Johnson W W. 2004.Rate-limiting steps in oxidations catalyzed by rabbit cytochrome P450 1A2. Biochemistry, 43(33): 10775-10788.
Hu Z,Lin Q, Chen H,etal. 2014. Identification of a novel cytochrome P450 gene,CYP321E1 from the diamondback moth,Plutellaxylostella(L.) and RNA interference to evaluate its role in chlorantraniliprole resistance. Bulletin of Entomological Research, 104 (6): 716.
Ioriatti C, Anfora G, Angeli G,etal. 2009. Effects of chlorantraniliprole on eggs and larvae ofLobesiabotrana(Denis & Schiffermüller) (Lepidoptera: Tortricidae). Pest Management Science, 65(6): 717-722.
Knight A L, Flexner L. 2007. Disruption of mating in codling moth (Lepidoptera: Tortricidae) by chlorantranilipole, an anthranilic diamide insecticide. Pest Management Science, 63(2): 180-189.
Koppenh?fer A M, Fuzy E M. 2008. Effect of the anthranilic diamide insecticide, chlorantraniliprole, onHeterorhabditisbacteriophora(Rhabditida: Heterorhabditidae) efficacy against white grubs (Coleoptera: Scarabaeidae). Biological Control, 45: 93-102.
Kuhar T, Doughty H, Hitchner E,etal. 2008. Evaluation of foliar insecticides for the control of Colorado potato beetle in potatoes in Virginia,2007. Arthropod Management, 17(1):140-145.
Lahm G, Cordova D, Barry J. 2009. New and selective ryanodine receptor activators for insect control. Bioorganic & Medicinal Chemistry, 17(12): 4127-4133.
Leonard D E. 1981. Bioecology of the gypsy moth∥Doane C C, McManus M L. The gypsy moth: Research toward integrated pest management. Washington, D.C.: U.S. Department of Agriculture, Forest Service, Science and Education Agency, Animal and Plant Health Inspection Service, 9-29.
Li X C, Baudry J, Berenhaum M R,etal. 2004. Structural and functional divergence of insect CYP6B proteins: From specialist to generalist cytochrome P450. Proceeding of the National Academy of Sciences USA, 101(9):2939-2944.
Li X C, Schuler M A, Berenbaum M R. 2007. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annual Review of Entomology, 52: 231-253.
Li A, Yang Y, Wu S,etal. 2006. Investigation on the resistance mechanisms to fipronil in diamondback moth. Journal of Economic Entomology, 99(3): 914-919.
Lin Q, Jin F, Hu Z,etal. 2013. Transcriptome analysis of chlorantraniliprole resistance development in the diamondback mothPlutellaxylostella. PLoS One, 8(8):e72314.
Liu X, Wang H Y, Ning Y B,etal. 2015. Resistance selection and characterization of chlorantraniliprole resistance inPlutellaxylostella(Lepidoptera: Plutellidae). Journal of Economic Entomology, 108(4):1978-1985.
Liu N, Scott J G. 1998. Increased transcription ofCYP6D1 causes cytochrome P450-mediated insecticide resistance in housefly. Insect Molecular Biology, 28(8): 531-535.
Lvaylo S. 2001.Cytochrome P450s: coupling development and environment. Trends in Genetics, 17(11):629-632.
Nelson D R.2009.The cytochrome P450 homepage.Human Genomics,4(1):59-65.
Pfaffl M W, Horgan G W, Dempfle L. 2002. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Research, 30(9):e36.
Riveron J M, Ibrahim S S, Chanda E,etal. 2014. The highly polymorphicCYP6M7 cytochrome P450 gene partners with the directionally selectedCYP6P9aandCYP6P9bgenes to expand the pyrethroid resistance front in the malaria vectorAnophelesfunestusin Africa. BMC Genomics, 15: 817.
Riveron J M, Irving H, Ndula M,etal. 2013. Directionally selected cytochrome P450 alleles are driving the spread of pyrethroid resistance in the major malaria vectorAnophelesfunestus. Proceeding of the National Academy of Sciences USA, 110(1): 252-257.
Spomer N A, Kamble S T, Siegfried B D. 2009. Bioavailability of chlorantraniliprole and indoxacarb to eastern subterranean termites (Isoptera: Rhinotermitidae) in various soils. Journal of Economic Entomology, 102(5): 1922-1927.
Sun L L, Wang Z Y, Zou C S,etal. 2014. Transcription profiling of 12 Asian gypsy moth(Lymantriadispar)cytochrome P450 genes in response to insecticides. Archives of Insect Biochemistry and Physiology, 85(4):181-194.
Teixeira L A F, Gut L J, Wise J C,etal. 2009. Lethal and sublethal effects of chlorantraniliprole on three species ofRhagoletisfruit flies (Diptera: Tephritidae). Pest Management Science, 65(2): 137-143.
Willis D K, Wang J, Lindholm J,etal. 2010. Microarray analysis of juvenile hormone response inDrosophilamelanogasterS2 cells. Journal Insect Science, 10: 66.
Yeoh B, Lee C Y. 2007. Tunneling responses of the Asian subterranean termite,Coptotermesgestroiin termiticide-treated sand (Isoptera: Rhinotermitidae). Sociobiology, 50(2): 457-468.
Zhao Y, Hull A K, Gupta N R,etal. 2002. Trp-dependent auxin biosynthesis inArabidopsis: involvement of cytochrome P450sCYP79B2 andCYP79B3. Genes & Development, 16(23): 3100-3112.
Zhou X J, Ma C X, Li M,etal. 2010.CYP9A12 andCYP9A17 in the cotton bollwormHelicoverpaarmigera: sequence similarity, expression profile and xenobiotic response. Pest Management Science, 66(1):65-73.
Zhu F, Parthasarathy R, Bai H,etal. 2010. A brain-specific cytochrome P450 responsible for the majority of deltamethrin resistance in the QTC279 strain ofTriboliumcastaneum. Proceeding of the National Academy of Sciences USA, 107(19):8557-8562.
(責任編輯 朱乾坤)
Chlorantraniliprole Stress
Dang Yingqiao Yin Jingjing Chen Chuanjia Sun Lili Liu Peng Cao Chuanwang
(CollegeofForestry,NortheastForestryUniversityHarbin150040)
【Objective】Lymantriadisparis a major forest pest. Cytochrome P450 is ubiquitous key metabolic detoxification enzyme for xenobiotics in insects. This study onCYP6AN15v1 detoxifying pesticides aims to provide theoretical basis forL.disparcontrol. 【Method】 The full length cDNA ofLdCYP6AN15v1 was cloned by RT-PCR technology. The transformantDrosophilavector expressingCYP6AN15v1 gene was constructed with the method of traditional restriction endonuclease digestion and ligation. Homozygous transformantDrosophilalines withLdCYP6AN15v1 were successfully constructed by using transformant technology. The effects of low dosage of chlorantraniliprole on cytochrome P450 activity andCYP6AN15v1 expression levels in transformant and untransformantDrosophilawere examined using spectrophotometry and real-time RT-PCR technology, respectively. 【Result】 The full length cDNA ofCYP6AN15v1 (namelyLdCYP6AN15v1) was isolated fromL.dispartranscriptome. The open reading frame (ORF) ofLdCYP6AN15v1 was 1 539 bp encoding a protein of 512 amino acid residues with the molecular mass of 59.02 kDa. Phylogenetic analysis of CYP proteins showed CYP6AN15v1 ofL.disparclustered into a group withSpodopteraexiguaandHelicoverpaarmigera. The transformantDrosophilaattP40>CYP6AN15v1 was detected 1 539 bp of target gene using DNA and cDNA as template showing successful expression ofLdCYP6AN15v1 into transformantDrosophila. Compared to untransformant attP40Drosophila, the susceptibility of attP40>CYP6AN15v1Drosophilato chlorantraniliprole was significantly decreased by 2.92-fold of untransformantDrosophilafor LC50. Under 7.17 mg·L-1chlorantraniliprole stress, the cytochrome P450 activity andCYP6AN15v1 expression inL.disparwere dependent on time effects. The cytochrome P450 activity of attP40>CYP6AN15v1Drosophilawas from 1.09- to 1.93-fold of untransformantDrosophilawhile mRNA expression levels ofCYP6AN15v1 in attP40>CYP6AN15v1Drosophilawere 44.54- to 137.80-fold of untransformantDrosophilashowing induction effects. 【Conclusion】 The transformantDrosophilaline attP40>CYP6AN15v1 was successfully constructed by using transgenic technology. The results suggest that the up-regulated expression ofLdCYP6AN15v1 gene induced by chlorantraniliprole could enhance P450 activity inL.disparlarvae to detoxify the chlorantraniliprole.
Lymantriadispar; transformantDrosophila; chlorantraniliprole;CYP6AN15v1; induced expression
10.11707/j.1001-7488.201706011
2016-08-13;
2017-01-20。
國家自然科學基金項目(31570642); 黑龍江省自然科學基金項目(C201409); 東北林業大學本科生創新項目(201510225183)。
S763.306
A
1001-7488(2017)06-0094-11
*曹傳旺為通訊作者。
