松材線蟲CYP450基因與松樹蒎烯類物質代謝的相關性*
2017-07-18 12:10:14理永霞劉振宇賈秀貞張星耀
林業科學 2017年6期
關鍵詞:檢測
王 璇 理永霞 劉振宇 呂 全 賈秀貞 張星耀
(1.中國林業科學研究院林業新技術研究所 北京100091; 2.中國林業科學研究院森林生態環境與保護研究所國家林業局森林保護學重點實驗室 北京 100091; 3.山東農業大學 泰安 271018)
?
松材線蟲CYP450基因與松樹蒎烯類物質代謝的相關性*
王 璇1理永霞2劉振宇3呂 全2賈秀貞2張星耀2
(1.中國林業科學研究院林業新技術研究所 北京100091; 2.中國林業科學研究院森林生態環境與保護研究所國家林業局森林保護學重點實驗室 北京 100091; 3.山東農業大學 泰安 271018)
【目的】 通過比較松材線蟲CYP450基因和寄主萜烯類物質代謝的時間特異性,研究松材線蟲解毒相關CYP450基因與寄主蒎烯類物質代謝之間的相關性,為該病致病機制的深入研究提供基礎。【方法】 松材線蟲接種5年生馬尾松后,通過實時定量PCR研究松材線蟲CYP450基因和寄主萜烯合成酶基因表達模式,同時通過氣相色譜法檢測寄主松樹主要萜烯類物質代謝規律。【結果】 松材線蟲接種5年生馬尾松后,松樹蒎烯合成酶基因分別在第6天和第21天上調表達,而寄主松樹體內α-蒎烯和β-蒎烯也具有2個峰值,分別在第9天和第27天時含量最高。松材線蟲CYP-33C9基因在第12天和第15天大量表達,CYP-33C4基因在接種后第21天表達量最高。【結論】 松材線蟲的入侵引起松樹蒎烯合成酶基因上調表達,生物合成大量蒎烯類物質,松材線蟲CYP450基因響應松樹蒎烯類物質的大量積累而上調表達。因此,根據松材線蟲CYP450基因與松樹蒎烯類物質代謝在時間上的相關性,推測CYP450基因可能參與了松材線蟲蒎烯類物質代謝過程,可能是松材線蟲致病相關基因之一。
松材線蟲; 蒎烯代謝; 細胞色素P450; 致病機制
松材線蟲病,即松樹萎蔫病(pine wilt disease,PWD),是一種以松材線蟲(Bursaphelenchusxylophilus)(Nickleetal., 1981)為病原,墨天牛屬昆蟲(Monochamusspp.)(Mamiya, 1983)為主要媒介,綜合人為參與、寄主松樹、相關伴生菌和環境因素互作的復雜病害系統(張星耀等, 2003)。該病傳播蔓延迅速,防治難度極大,是我國目前最為嚴重的森林災害。
松材線蟲侵入寄主松樹后取食松樹薄壁細胞,從而誘導薄壁細胞生物合成大量的蒎烯類物質(Kuroda, 1989),研究發現松材線蟲和海岸松(Pinuspinaster)共培養后,其揮發物主要是α-蒎烯和β-蒎烯(Wangetal., 2010),該類揮發性的蒎烯類物質在負壓條件下汽化形成氣泡(Ikedaetal., 1992),在管胞內形成空洞阻斷水柱的形成,同時其疏水性防止水分重新回到管胞中,從而引起薄壁細胞的空洞逐漸擴大,最終導致水分運輸受阻,松樹因缺水而死亡(Fariaetal., 2015)。因此,揮發性蒎烯類物質是寄主空洞化病理學過程中的特征性物質。
松樹響應松材線蟲入侵產生的次生代謝產物是松樹防御的重要手段,高濃度的蒎烯類化合物是松樹主要次生代謝產物,也是松材線蟲在松樹體內成功定殖必須面對的主要逆境之一。談家金等(2009)測定了松樹主要蒎烯類物質對松材線蟲的影響,結果表明β -蒎烯、β-水芹烯等抑制松材線蟲繁殖且β -水芹烯和莰烯具有殺松材線蟲活性。Niu(2012)發現α-蒎烯和β-蒎烯等在低濃度條件下抑制松材線蟲的繁殖,高濃度下可以促進松材線蟲的繁殖。因此,在寄生過程中,松材線蟲必須面對寄主次生代謝產物脅迫,對松樹次生代謝產物的降解就顯得至關重要。
細胞色素P450(cytochrome P450,CYP450)代謝途徑是生物體內有毒物質降解的重要代謝途徑(Arltetal., 2008; Zhaoetal., 2007),也是寄生性線蟲外源物質代謝中的重要代謝通路(Laingetal., 2015)。因此本研究通過松材線蟲接種5年生馬尾松(Pinusmassoniana)基礎上,檢測松樹主要蒎烯類物質的積累與松材線蟲CYP450基因表達模式之間的關系,從而探究松材線蟲細胞色素CP450基因在松材線蟲致病過程中的作用機制。
1 材料與方法
1.1 松材線蟲接種
1.1.1 松材線蟲處理 供試松材線蟲蟲株來自于本實驗所保存的Nxy61蟲株,該蟲株分離于2012年10月浙江寧波當年發病的馬尾松枝條,通過貝爾曼漏斗法分離后用灰葡萄孢(Botrytiscinerea)玉米培養基長期保存。試驗接種線蟲為混合蟲態松材線蟲(包括蟲卵、2~4齡幼蟲以及成蟲),在灰葡萄孢PDA培養基上25 ℃黑暗培養5天后,貝爾曼漏斗法收集分離線蟲,無菌水沖洗3次后,再用0.5%硫酸鏈霉素溶液處理2 min后,無菌水沖洗3次,調整線蟲懸浮液濃度20頭·μL-1后備用。
1.1.2 松材線蟲接種及取樣 本試驗接種所用松樹來自于中國林業科學研究院科研溫室所培養的5年生馬尾松,參照Otoguro等(1988)的方法,在馬尾松側枝距離主干1 cm處,剪斷側枝,將1.5 mL離心管的底部剪斷,用封口膜固定于切口處,往離心管注入1 mL NXY61線蟲液(10 000頭),以1 mL無菌水(CK)作為對照。分別在接種后1個月內,每隔3天取1次樣,共10次。每天3個重復。截取接種點以下1~5 cm部位枝條,用以檢測樹體內蒎烯含量; 截取距離接種處5 cm處枝條用以檢測松樹蒎烯合成酶apin基因表達模式研究; 收集接種點及接種松樹體內松材線蟲,用以松材線蟲CYP450基因表達模式研究。
1.2 松樹蒎烯合成酶apin基因表達模式研究
1.2.1 松樹RNA提取及反轉錄第1鏈合成 截取距離接種處5 cm處約1 cm長松枝,迅速剪碎裝于1.5 mL離心管內,液氮速凍后保存于-80 ℃冰箱,根據多酚植物RNA提取試劑盒(RNAprep pure Tissue Kit,TIANGEN, BeiJing)操作步驟提取松樹總RNA,通過電泳檢測其完整性,再用cDNA第1鏈合成試劑盒(FastQuant RT Kit,TIANGEN, BeiJing)反轉錄合成cDNA模板,其中總RNA的量統一為1 000 ng。
1.2.2 Real-time PCR檢測松樹蒎烯合成酶apin基因表達 本試驗所用儀器為ABI 7500 Real-time PCR 擴增儀。根據松樹蒎烯合成酶apin基因序列設計檢測引物,以松樹actin基因為內參,根據SYBR PreExTaqTM(TaKaRa code: DRR0820A)操作步驟進行熒光定量檢測。
擴增特異性分析: 設置5個梯度,分別將松樹cDNA模板稀釋為1,5,10,15,20倍,執行RT-PCR反應,通過擴增曲線Ct值對起始模板的定量分析,確定最適宜模板應進行10倍稀釋。根據融解曲線和PCR產物瓊脂糖凝膠電泳檢測,確認目的基因和內參基因的特異擴增引物如表1所示。
RT-PCR: 以接種后不同時間點的松材線蟲cDNA樣品為模板,并將模板稀釋10倍后,進行松樹apin基因Real-time PCR相對定量分析,反應體系為20 μL,每個樣品設4次重復,用 2-ΔΔCt法分析數據,確定基因相對表達量。
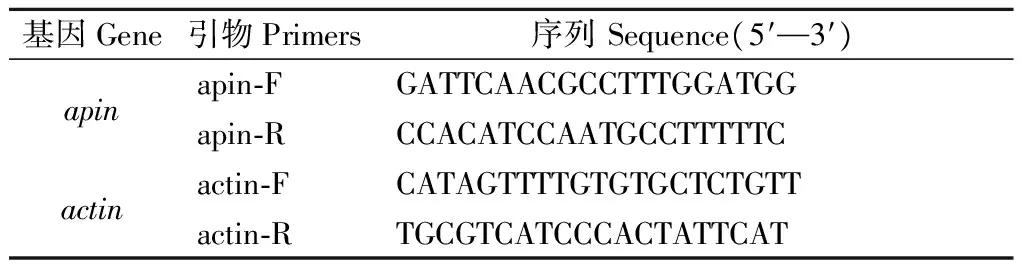
表1 apin基因Real-time PCR引物序列Tab.1 Real-time PCR primer sequences of apin gene
1.3 松樹蒎烯類物質含量檢測
1.3.1 α-蒎烯的提取方法 稱取1 g接種點1~5 cm處枝條,迅速剪成微小碎塊,放入5 mL離心管中,加入4 mL正己烷,蓋好管蓋,密閉浸泡24 h。然后將溶液振蕩搖勻,用0.45 μm濾膜過濾,裝入氣相瓶中待測。
1.3.2 氣相色譜法檢測α-蒎烯含量 通過氣相色譜儀(Agilent(GC-FID,7890A))檢測,手性柱(Cycoldex-B(30 m×0.25 mm× 0.25 μm))上樣。檢測條件為: 50 ℃保持3 min,以4 ℃·min-1的速度升至200 ℃,保持10 min; 載氣: 氮氣; 流速: 1.5 mL·min-1; 進樣溫度: 250 ℃; 進樣量0.5 μL; 無分流進樣; 保留時間: 3.68 min。將(-)-α-蒎烯和β-標準品按梯度稀釋,稀釋濃度為20,40,60,80,100 μL·L-1進氣相色譜儀檢測,制備標準曲線,根據樣品峰面積對樣品進行定量分析。
1.4 松材線蟲CYP450基因表達模式檢測
1.4.1 松材線蟲RNA提取及反轉錄第1鏈合成 收集接種松樹體內松材線蟲,液氮速凍后保存于-80 ℃冰箱,根據動物總RNA提取試劑盒(RNAprep pure Tissue Kit,TIANGEN, BeiJing)操作步驟提取線蟲總RNA,cDNA第1鏈合成同1.2.1。
1.4.2 Real-time PCR檢測松材線蟲CYP450基因表達 以不同接種時間點的松材線蟲cDNA為模板,松材線蟲β-actin基因為內參,根據松材線蟲CYP-33C9和CYP-33C4基因序列設計特異性引物,進行熒光定量檢測,操作步驟同1.2.2。最后選定引物序列見表2。
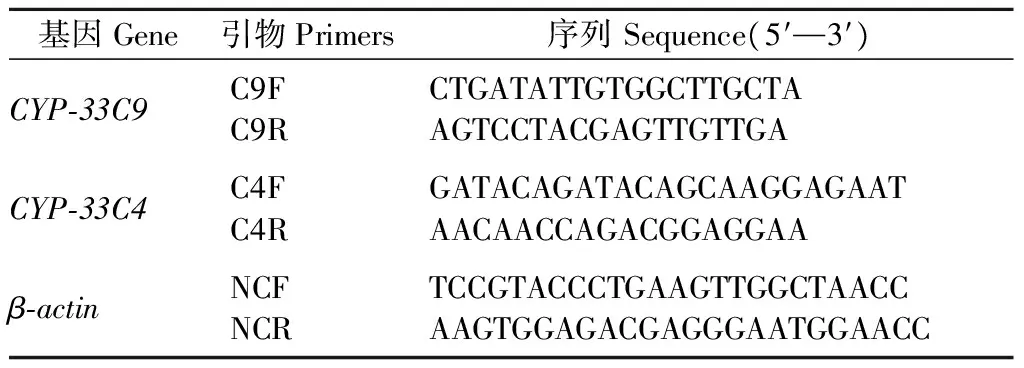
表2 CYP450基因 Real-time PCR引物序列Tab.2 Real-time PCR primer sequences of CYP450 gene
2 結果與分析
2.1 松樹蒎烯合成酶apin基因表達模式
如圖1所示,松樹蒎烯合成酶apin基因在松材線蟲接種5年生馬尾松后1個月內表達量有2個高峰,分別在第6天和第21天大量表達且其表達量遠遠高于無菌水接種的對照處理。對照組表達量較低且波動幅度較小。松材線蟲的入侵誘導了松樹蒎烯合成酶的大量表達,為后期蒎烯類物質的大量生物合成提供物質基礎。
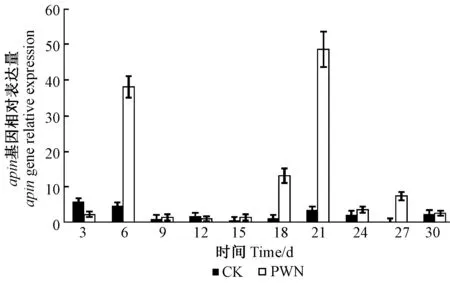
圖1 松樹apin基因相對表達量Fig.1 The apin gene relative expression
2.2 松材線蟲接種馬尾松后主要蒎烯類含量檢測
2.2.1 松材線蟲接種馬尾松后α-蒎烯含量檢測 松材線蟲接種5年生馬尾松后,松樹α-蒎含量有2個峰值,分別在9和27天時大量積累,在18和30天時含量最低。不同時期松材線蟲接種后松樹α-蒎烯含量均高于對照組含量。接種無菌水的對照組α-蒎烯含量21天時最高,12天時最低,整體變化幅度較小,趨于平行,而松材線蟲接種后松樹α-蒎烯含量變化明顯(圖2)。
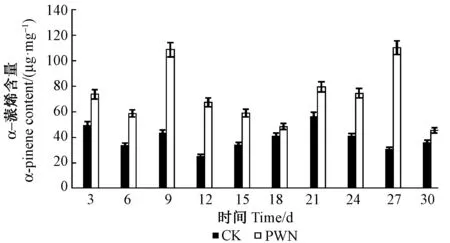
圖2 α-蒎烯含量Fig.2 The content of α-pinene
2.2.2 松材線蟲接種馬尾松后β-蒎烯含量檢測 松材線蟲接種馬尾松后松樹β-蒎烯含量也在9和27天時大量積累,呈現2個峰值,在18天時含量最低,21天時含量有所增加,27天時含量最高。對照組3天時β-蒎烯含量最高,9天時含量增加但小于3天的含量,隨后趨于不變,30天時有小幅度的增加(圖3)。
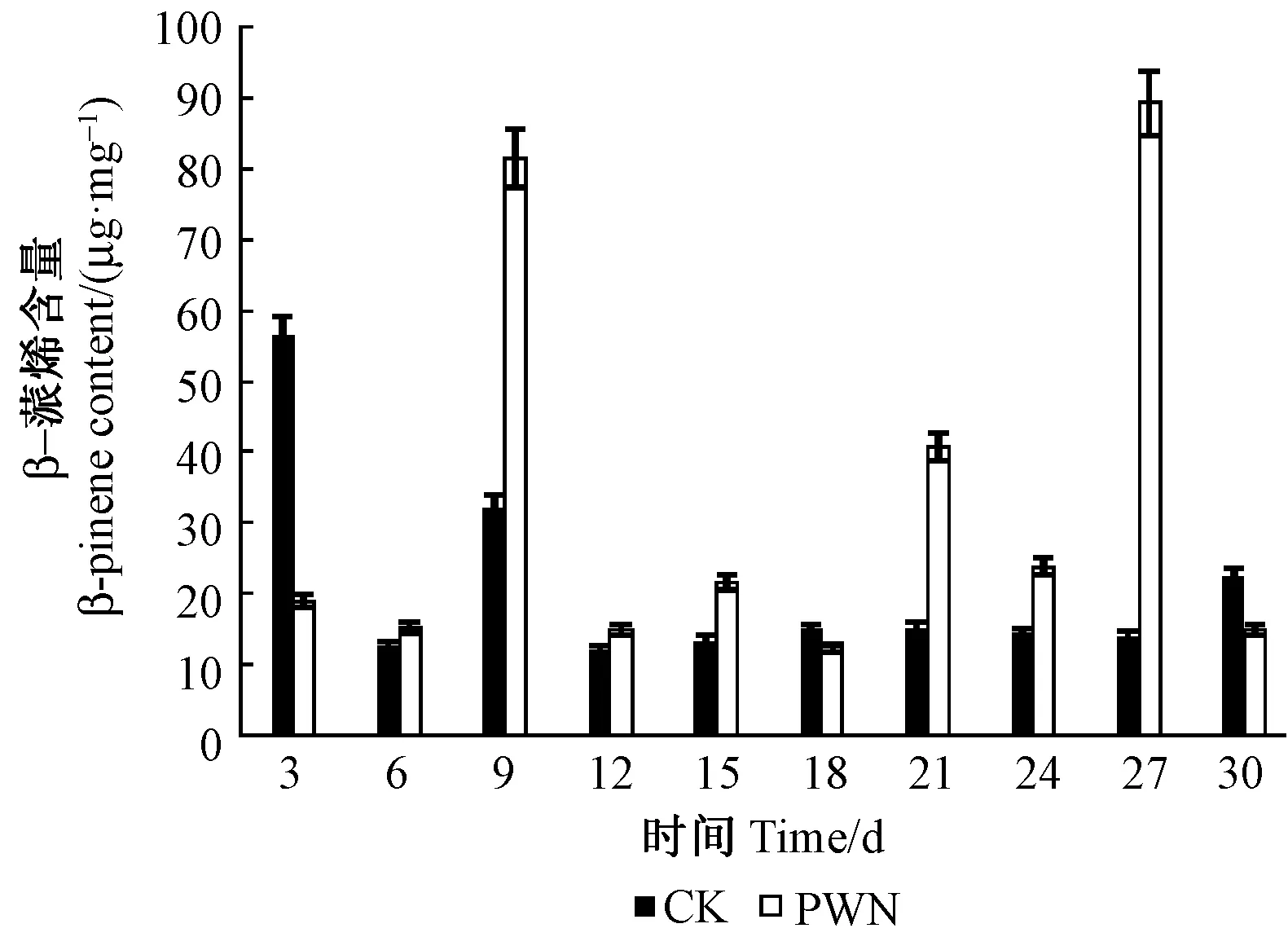
圖3 β-蒎烯含量Fig.3 The content of β-pinene
2.3 松材線蟲CYP450基因表達模式
2.3.1 松材線蟲CYP-33C9基因表達模式 松材線蟲CYP-33C9基因在松材線蟲接種后9天之前與對照組表達量相近,12和15天時超量表達,15天時最高,18天后幾乎不表達(圖4)。松樹α-蒎烯及β-蒎烯含量檢測結果發現松樹主要蒎烯類物質在9天是含量大幅增加,而CYP-33C9基因在12和15天時突然大量表達,因此,松材線蟲CYP-33C9基因響應松樹α-蒎烯及β-蒎烯含量的第1次大量積累,可能參與其蒎烯類物質的代謝過程,以便于松材線蟲的松樹體內的成功定殖和大量繁殖。
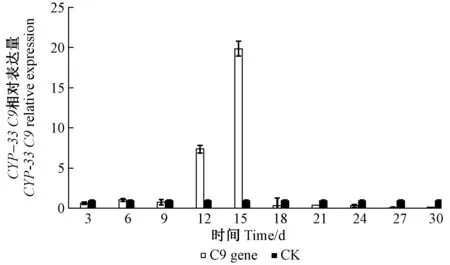
圖4 CPY-33C9相對表達量Fig.4 The CPY-33C9 relative expression
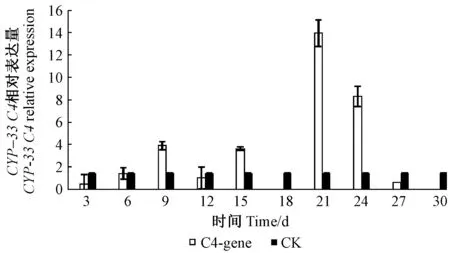
圖5 CPY-33C4相對表達量Fig.5 The CPY-33C4 relative expression
2.3.2 松材線蟲CYP-33C4基因表達模式 松材線蟲CYP-33C4基因在接種后21天時表達量最高,18和30天時沒有檢測到表達量; 在9天之前,其表達量逐漸增加,12天時有所降低,15天表達量與9天表達量相近,但在第18天時表達量很低,以至于沒有檢測到,21天時表達量急劇增至最高,隨后降低(圖5)。松樹蒎烯類物質是在9天時大量積累,而CYP-33C4基因在12~21天之間表達量呈現上升趨勢。因此,該基因可能響應蒎烯類物質的積累而大量表達,可能參與了松材線蟲蒎烯類物質代謝。
3 討論
Myers(1986)首次提出松材線蟲病空洞化理論,該理論認為,松樹萎蔫主要是由于管胞中出現的空洞影響了松樹水分輸導造成的。松樹感染了松材線蟲后,樹體內單蒎烯和倍半蒎烯的含量急劇增加,這類物質易汽化、表面張力低,滲入管胞后,在管胞中形成空洞,致使水分輸導受阻,從而引起松樹萎蔫(楊寶君, 2002)。Kuroda(1995)通過聲波發射(acoustic emission,AE)技術研究發現,松材線蟲接種后1~2個星期AE信號突然增加,并且在晚上和白天均有信號。接種3~4個星期后,AE信號比正常低,癥狀可見,老葉變黃,尖端萎蔫。這一時期1/2~2/3的枝條由于空洞化而功能紊亂,形成層和射線部分壞死,松樹接近死亡。因此,空洞化的形成是松材線蟲病典型的病理學特征。
松樹蒎烯特別是單萜類物質的積累是松樹響應松材線蟲入侵的主要防御反應,高濃度松樹單萜揮發物對松材線蟲具有生理毒性,被認為是松樹防御反應的重要化合物(Seyboldetal., 2006)。松材線蟲侵染感病松樹海岸松時,蒎烯次級代謝(包括殺線蟲活性物質)基因大量表達,松樹產生大量蒎烯類物質等次級代謝產物,抵御松材線蟲的入侵(Santosetal., 2012)。松材線蟲入侵后,寄主釋放的揮發性物質主要是α-蒎烯、β-蒎烯和長葉烯三種蒎烯類物質,其比例為1∶0.1∶0.1,α-蒎烯和β-蒎烯是主要的單萜類物質(Zhaoetal., 2007)。松材線蟲接種黑松(Pinusthunbergii)后,單萜和倍半萜含量顯著增加: α-蒎烯含量是健康松樹的2~4倍,β-蒎烯和其他幾種單萜是2~3倍(Kuroda, 1989)。α-蒎烯是松樹響應松材線蟲入侵的主要蒎烯類次生代謝產物,Takeuchi等(2006)發現自然感病的松樹和人工接種松材線蟲的松樹體內蒎烯類物質如α-蒎烯釋放量大量積累。本研究發現松材線蟲接種后,松樹蒎烯合成酶(apin)基因大量表達,蒎烯類物質大量積累,且都具有2個峰值。松樹apin基因大量表達的時間早于蒎烯類物質大量積累的時間,說明松材線蟲接種后,激發松樹蒎烯合成酶大量表達,大量合成蒎烯類物質。松材線蟲入侵后,分泌纖維素酶等細胞壁相關蛋白(Maetal., 2011),類毒液過敏原蛋白(Lietal., 2016)等激發寄主防御反應,從而引起蒎烯類次級代謝產物大量積累。因此,松材線蟲接種后馬尾松蒎烯類物質第1次大量增加是松樹響應松材線蟲的入侵而進行的主動防御反應; 而蒎烯類物質的第2次大量積累可能是因為松材線蟲大量繁殖后,通過其他致病基因干擾松樹蒎烯類物質代謝,從而使松樹蒎烯類物質代謝紊亂的結果。
松材線蟲入侵后,分泌一系列代謝相關蛋白通過結合、氧化還原等作用降低寄主次級代謝產物對松材線蟲的危害。松材線蟲對寄主松樹次級代謝產物的代謝利用預計有3個不同階段: 1) 給次級代謝產物加上功能團,使其更適合作為下游反應的底物; 2) 分解代謝過程; 3) 排出體外。而CYP450可能是第1階段最重要的酶(Kikuchi, 2011)。松材線蟲接種黑松后,CYP450等致病相關的基因高量表達(Urlacheretal., 2012)。Hirao等(2012)通過抑制消減雜交技術分析不同抗性黑松接種不同致病性線蟲的轉錄組發現,抗性黑松接種松材線蟲后CYP450蛋白和細胞壁相關水解酶類蛋白等轉錄組水平迅速減少,CYP450表達下調可能與松材線蟲入侵后樹脂道內產生蒎烯類化合物有關。
CYP450家族的酶在次級代謝產物生物轉化過程中有重要作用。Yan等(2012)通過松材線蟲和擬松材線蟲比較轉錄組學分析結果顯示CYP450是外源物質分解利用的主要代謝途徑。本研究發現松材線蟲CYP-33C9和CYP-33C4基因響應松樹蒎烯類物質的增加而表達量增加,推測CYP450基因可能參與了松材線蟲蒎烯類物質代謝過程。但隨著松材線蟲的入侵,松樹體內蒎烯類物質再次積累,還存在其他致病基因的作用,有助于松材線蟲躲避松樹次級代謝產物對松材線蟲的抑制作用。Lindblom等(2006)認為線蟲體內的外源物質代謝相關基因主要包括CYP450、短鏈脫氫酶(SDR)、谷胱甘肽轉移酶(GST)和糖醛酸脫氫酶(UGT)等。Kikuchi(2011)通過KEGG分析,松材線蟲的細胞色素P450的基因拷貝數明顯大于秀麗隱桿線蟲(C.elegans),是進行外源物質和有毒物質代謝最主要的路徑之一,可能是CYP450基因的下游基因。
通過松材線蟲與松樹互作早期特異表達基因研究中發現有7種CYP450基因顯著變化,4種基因上調表達,3種基因下調表達,其中CYP-33C4和CYP-33C9變化最明顯(Qiuetal., 2013)。Xu等(2015)通過RACE技術克隆了松材線蟲CYP-33C9、CYP-33C4、CYP-33D3基因,RNAi沉默這些基因后,松材線蟲活性、遷移率、繁殖率、致病性和對阿維菌素等殺蟲劑耐受性降低。本研究發現接種松材線蟲后CYP-33C4基因表達量均低于對照組,這可能是因為CYP-33C4基因與CYP-33C9基因存在同工酶的關系,CYP-33C9基因大量表達可能抑制CYP-33C4基因的表達。也有可能是因為取樣時間間距太大,CYP-33C4表達量最高點不在各個取樣時間點之內。松材線蟲CYP-33C9基因響應松樹α-蒎烯和β-蒎烯大量積累而超量表達,說明該基因有可能參與了松材線蟲對松樹蒎烯類物質降解過程。
4 結論
本研究發現松材線蟲接種后,松樹主要揮發性物質α-蒎烯和β-蒎烯的大量積累與松材線蟲CYP-33C4和CYP-33C9基因的大量表達在時間上存在緊密聯系,因此推測CYP450基因可能參與了松材線蟲蒎烯類物質代謝過程,可能是松材線蟲致病過程中的相關基因之一。此結果為進一步深入研究CYP450在松材線蟲致病機制中的作用提供了重要的參考依據。
談家金, 郝德君, 潘玉雯, 等. 2009. 幾種松樹揮發物對松材線蟲行為的影響. 東北林業大學學報, 37(12): 58-59.
(Tan J J, Hao D J, Pan Y W,etal. 2009. Effects of several pine volatiles on behavior ofBursaphelenchusxylophilus. Journal of Northeast Forestry University, 37(12): 58-59.[in Chinese])
楊寶君. 2002. 松材線蟲病致病機理的研究進展.中國森林病蟲, 21(1): 27-31, 14.
(Yang B J. 2002. Advance in research of pathogenetic mechanism of pine wood nematode. Forest Pest and Disease, 21(1): 27-31, 14. [in Chinese])
張星耀, 駱有慶. 2003. 中國森林重大生物災害. 北京: 中國林業出版社, 1-29.
(Zhang X Y, Luo Y Q. 2003. Major Forest Diseases and Insect Pests in China. Beijing: China Forestry Publishing House, 1-29. [in Chinese])
Arlt V M, Stiborová M, Henderson C J,etal. 2008. Metabolic activation of benzo [a] pyreneinvitroby hepatic cytochrome P450 contrasts with detoxificationinvivo: experiments with hepatic cytochrome P450 reductase null mice. Carcinogenesis, 29(3): 656-665.
Faria J M S, Sena I, da Silva I V,etal. 2015.Invitroco-cultures ofPinuspinasterwithBursaphelenchusxylophilus: a biotechnological approach to study pine wilt disease. Planta, 241(6): 1325-1336.
Hirao T, Fukatsu E, Watanabe A. 2012. Characterization of resistance to pine wood nematode infection inPinusthunbergiiusing suppression subtractive hybridization. BMC Plant Biology, 12(1): 13.
Ikeda T, Ohtsu M. 1992. Detection of xylem cavitation infield-grown pine trees using the acoustic emission technique. Ecol Res, 7(3): 391- 395.
Kikuchi T, Cotton J A, Dalzell J J,etal. 2011. Genomic insights into the origin of parasitism in the emerging plant pathogenBursaphelenchusxylophilus. PLoS Pathogens, 7(9): e1002219
Kuroda K. 1989. Terpenoids causingtracheid-cavitation inPinusthunbergiiinfected by the pine wood nematode (Bursaphelenchusxylophilus). Ann Phytopathol Soc Jpn, 55(2): 170-178.
Kuroda K. 1995. Acoustic emission technique for the detection of abnormal cavitation in pine trees infected with pine wilt disease∥Yang B J. Proceedings of International Symposium on Pine Wilt Disease Caused by Pine Wood Nematode, 53-58.
Laing R, Bartley D J, Morrison A A,etal. 2015. The cytochrome P450 family in the parasitic nematodeHaemonchuscontortus. International Journal for Parasitology, 45(4): 243-251.
Li Y X, Wang Y, Liu Z Y,etal. 2016. Functional analysis of the venom allergen-like protein gene from pine wood nematodeBursaphelenchusxylophilususing a baculovirus expression system. Physiological and Molecular Plant Pathology, 93: 58-66.
Lindblom T H, Dodd A K. 2006. Xenobiotic detoxification in the nematodeCaenorhabditiselegans. J Exp Zool A Comp Exp Biol, 305(9): 720-730.
Ma H B, Lu Q, Liang J,etal. 2011. Functional analysis of the cellulose gene of the pine wood nematode,Bursaphelenchusxylophilus, using RNA interference. Genetics and Molecular Research: GMR, 10(3):1931-1941.
Mamiya Y. 1983. Pathology of the pine wilt disease caused byBursaphelenchusxylophilus. Annual Review of Phytopathology, 21(1): 201-220.
Myers R F. 1986. Cambium destruction in conifers caused by pine wood nematodes. Journal of Nematology, 18(3): 398.
Nickle W R, Golden A M, Mamiya Y,etal. 1981. On the taxonomy and morphology of the pine wood nematode,Bursaphelenchusxylophilus(Steiner &Buhrer 1934) Nickle 1970. Journal of Nematology, 13(3): 385.
Niu H T, Zhao L L, Lu M,etal. 2012.The ratio and concentration of two monoterpenes mediate fecundity of the pinewood nematode and growth of its associated fungi.PLoS One, 7(2): e31716.
Otoguro K, Liu Z X, Fukuda K,etal. 1988. Screening for new nematocidal substances of microbial origin by a new method using the pine wood nematode. The Journal of Antibiotics, 41(4): 573-575.
Qiu X, Wu X, Huang L,etal. 2013. Specifically expressed genes of the nematodeBursaphelenchusxylophilusinvolved with early interactions with pine trees. PLoS One, 8(10): e78063.
Santos C S, Pinheiro M, Silva A I,etal. 2012. Searching for resistance genes toBursaphelenchusxylophilususing high throughput screening. BMC Genomics, 13(1): 599.
Seybold S J, Huber D P W, Lee J C,etal. 2006. Pine monoterpenes and pine bark beetles: a marriage of convenience for defense and chemical communication. Phytochemistry Reviews, 5(1): 143-178.
Takeuchi Y, Kanzaki N, Futai K. 2006. Volatile compounds in pine stands suffering from pine wilt disease: qualitative and quantitative evaluation. Nematology, 8(6): 869-880.
Urlacher V B, Girhard M. 2012. Cytochrome P450 monooxygenases: an update on perspectives for synthetic application. Trends in Biotechnology, 30(1): 26-36.
Wang Z, Wang C Y, Fang Z M,etal. 2010. Advances in research of pathogenic mechanism of pine wilt disease. Afr J Microbiol Res, 4(6): 437-442.
Xu X L, Wu X Q, Ye J R,etal. 2015. Molecular characterization and functional analysis of three pathogenesis-related cytochrome P450 genes fromBursaphelenchusxylophilus(Tylenchida: Aphelenchoidoidea). International Journal of Molecular Sciences, 16(3): 5216-5234.
Yan X, Cheng X Y, Wang Y S,etal. 2012. Comparative transcriptomics of two pathogenic pinewood nematodes yields insights into parasitic adaptation to life on pine hosts. Gene, 505(1):81-90.
Zhao L L, Wei W, Kang L,etal. 2007. Chemotaxis of the pinewood nematode,Bursaphelenchusxylophilus, to volatiles associated with host pine,Pinusmassoniana, and its vectorMonochamusalternatus. Journal of Chemical Ecology, 33(6): 1207-1216.
(責任編輯 朱乾坤)
Relationship between the Cytochrome P450 Gene of Pine WoodNematode and the Accumulation of Pine Pinene
Wang Xuan1Li Yongxia2Liu Zhenyu3Lü Quan2Jia Xiuzhen2Zhang Xingyao2
(1.ResearchInstituteofForestryNewTechnology,CAFBeijing100091; 2.KeyLaboratoryofForestProtectionofStateForestryAdministrationResearchInstituteofForestEcology,EnvironmentandProtection,CAFBeijing100091;3.ShandongAgriculturalUniversityTai’an271018)
By comparing the expression pattern of cytochrome P450 (CYP450) genes of Pine Wood Nematode (PWN) (Bursaphelenchusxylophilus)and the accumulation of pine terpene, the relationship between the CYP450 genes of PWN and the metabolism of pine pinene was analyzed to provide fundamental information for PWN pathogenic mechanisms study. 【Method】Five-yearsPinusmassonianawere inoculated with PWN. Real-time Quantitative PCR was conducted to detect expression patterns ofapingene of pine trees and CYP450 family genes of PWN. Terpene metabolism in infectedP.massonianawas detected by Gas Chromatography. 【Result】The expression of pinene synthase genes were dramatically induced at 6 dpi (days post inoculation) and 21 dpi. Both α-pinene and β-pinene were accumulated with two peaks at 9dpi and 27dpi, respectively. Correspondingly, the expression of PWNCYP-33C9 dramatically increased at 12dpi and 15dpi, and the expression ofCYP-33C4 were highest at 21dpi. 【Conclusion】The pine tree pinene synthase gene was highly induced by PWN inoculation which resulted in sharp accumulation of the α-pinene and β-pinene. In responding to the accumulation of pine terpene, the expression of PWN CYP450 genes dramatically increased. Therefore, according to the temporal correlation between the pinene metabolism and the PWN CYP450 genes expression we inferred that the PWN CYP450 genes might participate in the terpene metabolism and might be one of the pathogenic genes in the PWN pathogenic process.
pine wood nematode; terpene metabolism; cytochrome P450; pathological mechanisms
10.11707/j.1001-7488.20170612
2015-11-16;
2016-03-10。
浙江省省院合作林業科技項目(2014SY17); 林業公益性行業科研專項經費(201204501)。
S763.11
A
1001-7488(2017)06-0105-06
*理永霞為通訊作者。
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
