環(huán)割對(duì)杉木和馬尾松人工林土壤微生物群落結(jié)構(gòu)的影響*
2017-07-18 12:10:14賀同鑫孫建飛李艷鵬俞有志胡寶清王清奎
林業(yè)科學(xué) 2017年6期
關(guān)鍵詞:影響
賀同鑫 孫建飛 李艷鵬 俞有志 胡寶清 王清奎
(1.廣西師范學(xué)院 北部灣環(huán)境演變與資源利用教育部重點(diǎn)實(shí)驗(yàn)室 廣西地表過(guò)程與智能模擬重點(diǎn)實(shí)驗(yàn)室 南寧 530001;
?
環(huán)割對(duì)杉木和馬尾松人工林土壤微生物群落結(jié)構(gòu)的影響*
賀同鑫1,2孫建飛1李艷鵬2俞有志2胡寶清1王清奎2
(1.廣西師范學(xué)院 北部灣環(huán)境演變與資源利用教育部重點(diǎn)實(shí)驗(yàn)室 廣西地表過(guò)程與智能模擬重點(diǎn)實(shí)驗(yàn)室 南寧 530001;
2.中國(guó)科學(xué)院會(huì)同森林生態(tài)實(shí)驗(yàn)站 會(huì)同 418307)
杉木; 馬尾松; 人工林; 磷脂脂肪酸; 環(huán)割; 微生物群落結(jié)構(gòu)
土壤微生物通過(guò)調(diào)控陸地生態(tài)系統(tǒng)元素轉(zhuǎn)化和物質(zhì)循環(huán)過(guò)程,對(duì)土壤肥力的形成和保育產(chǎn)生重要影響,進(jìn)而影響土壤健康和生態(tài)服務(wù)功能。土壤微生物通過(guò)種群的消長(zhǎng)與植物的養(yǎng)分吸收形成互補(bǔ),維持和調(diào)節(jié)著生態(tài)系統(tǒng)的元素生物地球化學(xué)過(guò)程(方麗娜等, 2011)。土壤微生物能夠調(diào)控土壤碳庫(kù),反之,土壤碳有效性也能影響土壤微生物群落結(jié)構(gòu)(Brantetal., 2006; Tóthetal., 2007)。植物最新固定的光合產(chǎn)物約有30%~60%被輸送到根系,其中約20%會(huì)以根系分泌物的形式進(jìn)入到土壤中(Muchaetal., 2005; Yarwoodetal., 2009)。根系及根系分泌物是土壤微生物可利用碳的重要來(lái)源(Wangetal., 2013),光合產(chǎn)物供應(yīng)的變化可能會(huì)通過(guò)改變根系分泌物進(jìn)而影響土壤微生物群落結(jié)構(gòu)。因此了解光合產(chǎn)物供應(yīng)對(duì)土壤微生物的影響有助于更好地探索碳輸入的改變對(duì)土壤碳循環(huán)的影響。
自H?gberg等(2001)利用環(huán)割法研究改變光合產(chǎn)物供應(yīng)對(duì)土壤呼吸的影響之后,因其對(duì)土壤-根系-生物系統(tǒng)的擾動(dòng)較少,因而被越來(lái)越多的學(xué)者采用。前人研究表明環(huán)割通過(guò)阻斷光合產(chǎn)物供應(yīng),改變土壤理化性質(zhì)(尤其是活性碳庫(kù)和氮庫(kù)),從而對(duì)土壤微生物群落結(jié)構(gòu)產(chǎn)生重要影響(H?gbergetal., 2007; 2010; Kaiseretal., 2010; Chenetal., 2012; Keeletal., 2012)。然而不同生態(tài)系統(tǒng)中環(huán)割對(duì)土壤微生物的影響有很大的差別,如在歐洲赤松(Pinussylvestris)和歐洲云杉(Piceaabies)林中環(huán)割導(dǎo)致微生物生物量顯著下降30%~40% (P< 0.05)(H?gbergetal., 2001; Subkeetal., 2004)。而在尾葉桉(Eucalyptusurophylla)和厚莢相思(Acaciacrassicarpa)2個(gè)人工林中,環(huán)割對(duì)土壤微生物生物量并沒(méi)有產(chǎn)生顯著影響,同時(shí)其研究表明環(huán)割降低了真菌含量,但增加了細(xì)菌含量,因而改變了土壤微生物群落結(jié)構(gòu),然而環(huán)割對(duì)2種人工林的影響程度具有較大的差異(Chenetal., 2012)。這表明土壤微生物群落結(jié)構(gòu)對(duì)環(huán)割的響應(yīng)可能存在樹(shù)種差異,但是這方面的研究還較少,限制了深入理解光合產(chǎn)物供應(yīng)的改變對(duì)土壤微生物群落結(jié)構(gòu)的影響。
目前,利用環(huán)割的方法探討光合產(chǎn)物供應(yīng)對(duì)土壤碳庫(kù)、土壤呼吸及樹(shù)干呼吸影響的研究較多(H?gbergetal., 2001; Chenetal., 2010; Maieretal., 2010; Trahanetal., 2015),而對(duì)土壤微生物群落影響的研究則相對(duì)較少,尤其是樹(shù)種差異性更是鮮有報(bào)道。為了研究環(huán)割對(duì)土壤微生物群落結(jié)構(gòu)的影響是否具有樹(shù)種差異性,且其時(shí)間效應(yīng)又是怎樣的?本研究在湖南中國(guó)科學(xué)院會(huì)同森林生態(tài)實(shí)驗(yàn)站選擇了亞熱帶地區(qū)常見(jiàn)的杉木(Cunninghamialanceolata)和馬尾松(Pinusmassoniana)2種人工純林,在林中分別設(shè)置6個(gè)6 m × 6 m樣方,并進(jìn)行環(huán)割試驗(yàn)。環(huán)割1個(gè)月和1年后利用磷脂脂肪酸的方法測(cè)定土壤微生物群落結(jié)構(gòu),探討環(huán)割對(duì)不同樹(shù)種土壤微生物的影響,環(huán)割1個(gè)月后測(cè)定土壤基本理化性質(zhì),探討環(huán)割后土壤碳氮有效性的變化及其對(duì)土壤微生物的影響,以期揭示環(huán)割對(duì)土壤微生物群落結(jié)構(gòu)的影響。
1 研究區(qū)概況
研究區(qū)位于湖南省西南部的中國(guó)科學(xué)院會(huì)同森林生態(tài)實(shí)驗(yàn)站(110°08′E,27°09′N(xiāo))。該地區(qū)為低山丘陵地貌類(lèi)型,海拔200~500 m,屬于典型的亞熱帶濕潤(rùn)氣候,年均氣溫16.5 ℃,年降水量1 200~1 400 mm,土壤為山地紅黃壤,pH值為4~6。地帶性植被為以絲栗栲(Castanopsisfargesii)、青岡(Cyclobalanopsisglauca)和刨花潤(rùn)楠(Machiluspauhoi)等為建群種的天然常綠闊葉林; 主要人工林類(lèi)型為杉木、毛竹(Phyllostachysedulis)和馬尾松人工純林。
2 研究方法
2.1 試驗(yàn)設(shè)置、環(huán)割方法和植被管理
2012年6月,分別在26年生杉木及23年生馬尾松2個(gè)人工純林內(nèi)開(kāi)展試驗(yàn),2個(gè)人工林的密度均為1 200株·hm-2。在每個(gè)林中選取6個(gè)6 m × 6 m樣方,每個(gè)樣方至少包含5棵樹(shù)。樣地坡度約為23°,坡向?yàn)槟掀拢挛粸橹衅隆M寥篮?0~80 cm,土壤密度為1.2 g·cm-3,土壤孔隙度約為35%。土壤質(zhì)地較黏重,為中黏壤至重黏壤,土體中石礫含量約15%。杉木和馬尾松平均樹(shù)高分別為16.3和15.3 m,平均胸徑分別為18.5和17.2 cm。杉木林和馬尾松林的枯落物層厚度分別約為5和2 cm,郁閉度分別為0.7和0.8。
每種林分6個(gè)樣方中隨機(jī)選3個(gè)為對(duì)照(CK),3個(gè)進(jìn)行環(huán)割處理。環(huán)割處理是指在樹(shù)木胸徑處去除10 cm寬的樹(shù)皮及韌皮部。環(huán)割處理后杉木和馬尾松在環(huán)割部位下方萌芽,因此每個(gè)月去掉萌芽。馬尾松環(huán)割后3個(gè)月開(kāi)始掉葉,掉在地上的葉子盡快移除樣地外以避免在樣地內(nèi)積累。杉木的凋落物比較特殊,葉同枝一起掉落,且一般即使葉子枯萎也不會(huì)立馬掉落,因此杉木林中的凋落物層沒(méi)有太大變化。
杉木林中林下植被主要有杜莖山(Maesajaponica)、薔薇葉懸鉤子(Rubusrosifolius)、蕨(Pteridiumaquilinum)、中華金星蕨(Parathelypterischinensis)和邊緣鱗蓋蕨(Microlepiamarginata),在試驗(yàn)期間每月都在環(huán)割和對(duì)照小區(qū)用砍刀清除林下植被,為避免對(duì)土壤干擾,只去掉地上部分,并移除。馬尾松林中的林下植被很少,沒(méi)有進(jìn)行處理。
2.2 土壤樣品采集及室內(nèi)處理

2.3 土壤理化指標(biāo)測(cè)定
2.4 土壤微生物群落結(jié)構(gòu)分析
土壤微生物群落結(jié)構(gòu)的測(cè)定采用磷脂脂肪酸(phospholipid fatty acid, PLFA)分析方法,采用改進(jìn)后的簡(jiǎn)單提取法(Frosteg?rdetal., 1996; Bossioetal., 1998),主要分為脂類(lèi)提取、分離及甲基化3個(gè)步驟。1)按照1∶4的比例加入提取液(檸檬酸緩沖液、氯仿和甲醇按0.8∶1∶2體積比混合)于黑暗中充分震蕩3 h,然后離心取上清液(2 500 r·min-1離心10 min)。重復(fù)此過(guò)程,將2次上清液合并。向上清液中加入氯仿及緩沖液,混勻,靜止過(guò)夜。吸取氯仿層,N2吹干。2)依次采用氯仿、丙酮和甲醇分離出中性脂、糖脂和磷脂,收集甲醇相,N2吹干。3)甲基化: 將磷脂溶于1∶1的甲醇和甲苯溶液中,加入0.2 mol·L-1的氫氧化鉀甲醇溶液進(jìn)行皂化,最后用正己烷萃取,收集正己烷相,即為磷脂脂肪酸甲酯。在測(cè)定前2~3天,加入19∶0的內(nèi)標(biāo)以定量。采用氣相色譜質(zhì)譜儀(安捷倫GC-MS,6 890 N-5 973 N)測(cè)定每個(gè)樣品中的磷脂脂肪酸甲酯。PLFA含量用 nmol·g-1表示。從土壤中提取的PLFA的量可表征土壤微生物的群落結(jié)構(gòu)。PLFA的群落分類(lèi)詳見(jiàn)賀同鑫等(2015),PLFA的命名方式中的i、a、cy和Me,分別表示異、反異、環(huán)丙基和甲基分枝脂肪酸; ω、c和t分別表示脂肪端、順式空間構(gòu)造和反式空間構(gòu)造。
2.5 數(shù)據(jù)處理
所有數(shù)據(jù)統(tǒng)計(jì)分析基于SPSS 17.0軟件進(jìn)行。采用獨(dú)立樣本t檢驗(yàn)分析土壤微生物群落各組分(細(xì)菌、真菌和放線菌)和土壤基本理化性質(zhì)的差異。相關(guān)分析采用Pearson檢驗(yàn)法。在統(tǒng)計(jì)檢驗(yàn)過(guò)程中,顯著水平為P< 0.05,極顯著水平為P< 0.01。
3 結(jié)果與分析
3.1 樹(shù)木生長(zhǎng)變化
杉木具有很強(qiáng)的萌發(fā)能力,環(huán)割1個(gè)月后,其枝葉沒(méi)有顯著變化; 環(huán)割1年后仍具萌芽能力,大部分枝葉仍是綠色的,因此表明杉木仍有大部分根系存活。馬尾松萌芽能力很弱,環(huán)割3個(gè)月內(nèi),樹(shù)木80%的枝葉已經(jīng)死亡; 環(huán)割1年后枝葉全部凋落,樹(shù)木已經(jīng)全部死亡。
3.2 土壤性質(zhì)變化
3.3 土壤微生物群落結(jié)構(gòu)
杉木和馬尾松2種人工純林的土壤微生物生物量表現(xiàn)為細(xì)菌>真菌>放線菌含量(圖1b,c,h),表明在亞熱帶人工林中土壤微生物是以細(xì)菌為主體的群落結(jié)構(gòu)。環(huán)割處理1個(gè)月后和1年后杉木和馬尾松土壤總微生物生物量(總磷脂脂肪酸含量)、細(xì)菌、真菌和放線菌含量總體呈現(xiàn)降低趨勢(shì),而且其群落結(jié)構(gòu)均發(fā)生了顯著變化(圖 1)。環(huán)割處理1個(gè)月后,杉木林土壤微生物生物量和群落結(jié)構(gòu)變化比馬尾松林大; 但1年后,馬尾松比杉木變化大(圖1a,d,i)。
杉木林中,環(huán)割1個(gè)月后總磷脂脂肪酸、細(xì)菌、真菌含量、真菌∶細(xì)菌和革蘭氏陰性細(xì)菌含量分別下降了10.3%,10.9%, 20.0%,10.1%和13.4%(P< 0.05); 環(huán)割1年后,細(xì)菌和革蘭氏陰性細(xì)菌含量分別下降了20.3%和22.1%(P< 0.05)。馬尾松林中,環(huán)割1個(gè)月后真菌和革蘭氏陽(yáng)性細(xì)菌含量分別下降21.9%和14.5%(P< 0.05); 環(huán)割1年后,總磷脂脂肪酸、細(xì)菌和放線菌含量下降了17.8%,15.9%和27.4%(P< 0.05)。此外,杉木林中環(huán)割處理1個(gè)月后磷脂脂肪酸cy17∶0/16∶1ω7c顯著升高,而在馬尾松林中環(huán)割處理1年后該比值顯著升高(P< 0.05,圖1i)。
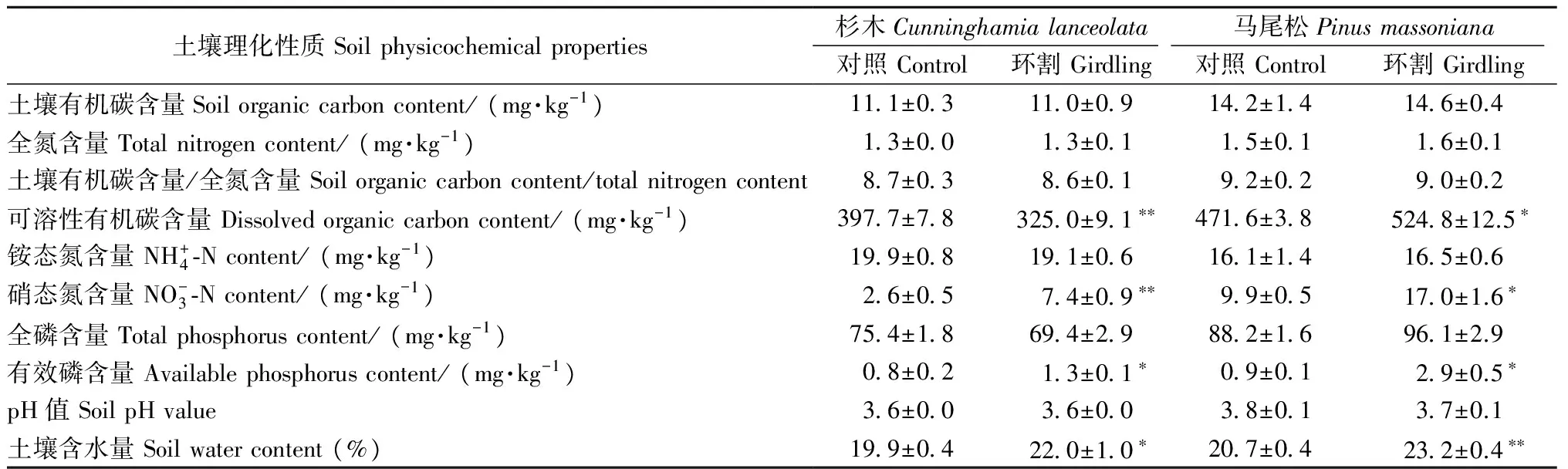
表1 環(huán)割1個(gè)月后土壤理化性質(zhì)變化①Tab. 1 Variation of soil physicochemical properties after one month of girdling
① *:P< 0.05;**:P< 0.01.下同The same below.
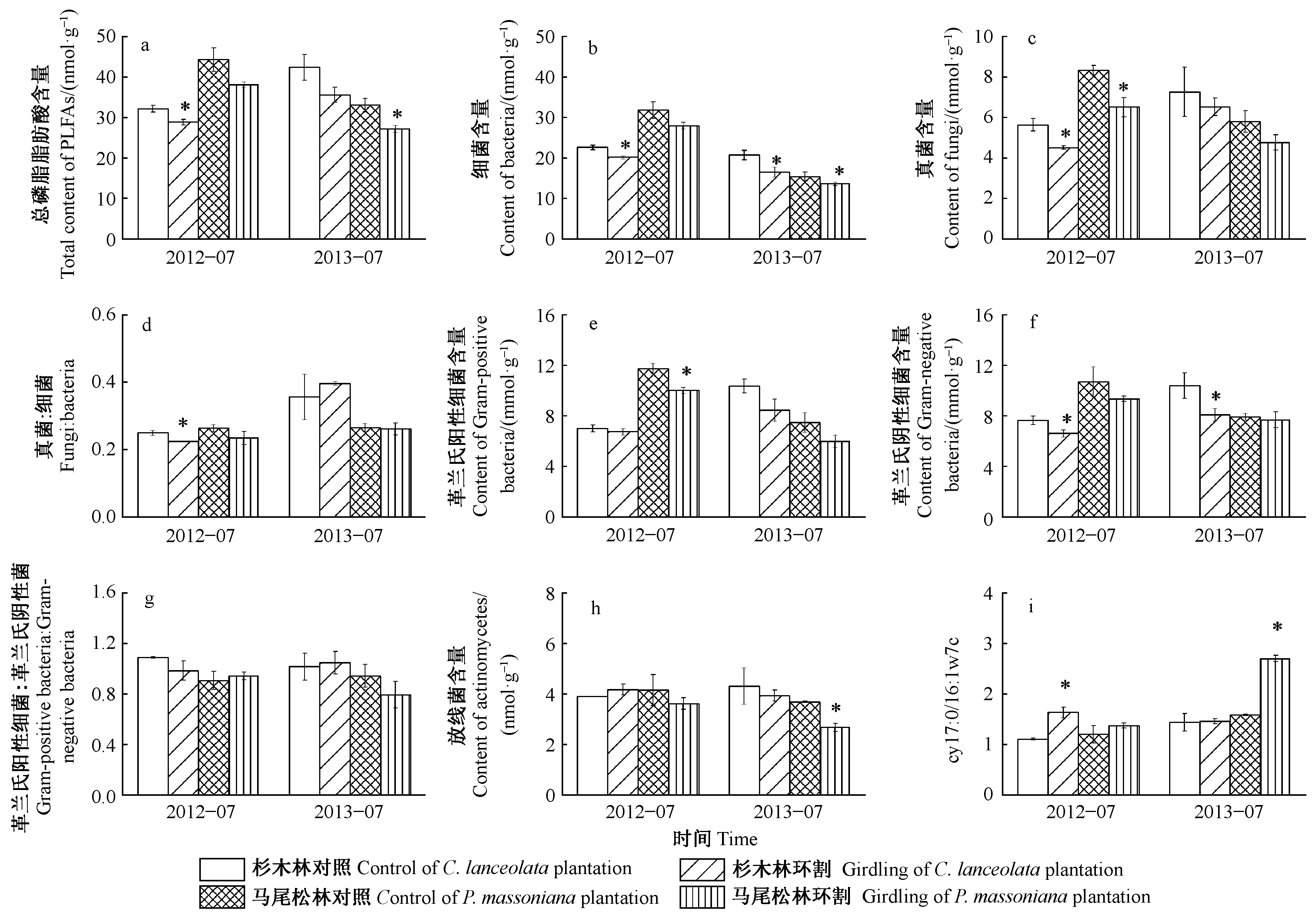
圖1 環(huán)割處理1個(gè)月后和1年后土壤微生物群落結(jié)構(gòu)變化Fig.1 Variation of soil microbial community composition after one month and one year of girdling
3.4 土壤微生物和土壤性質(zhì)的相關(guān)性分析

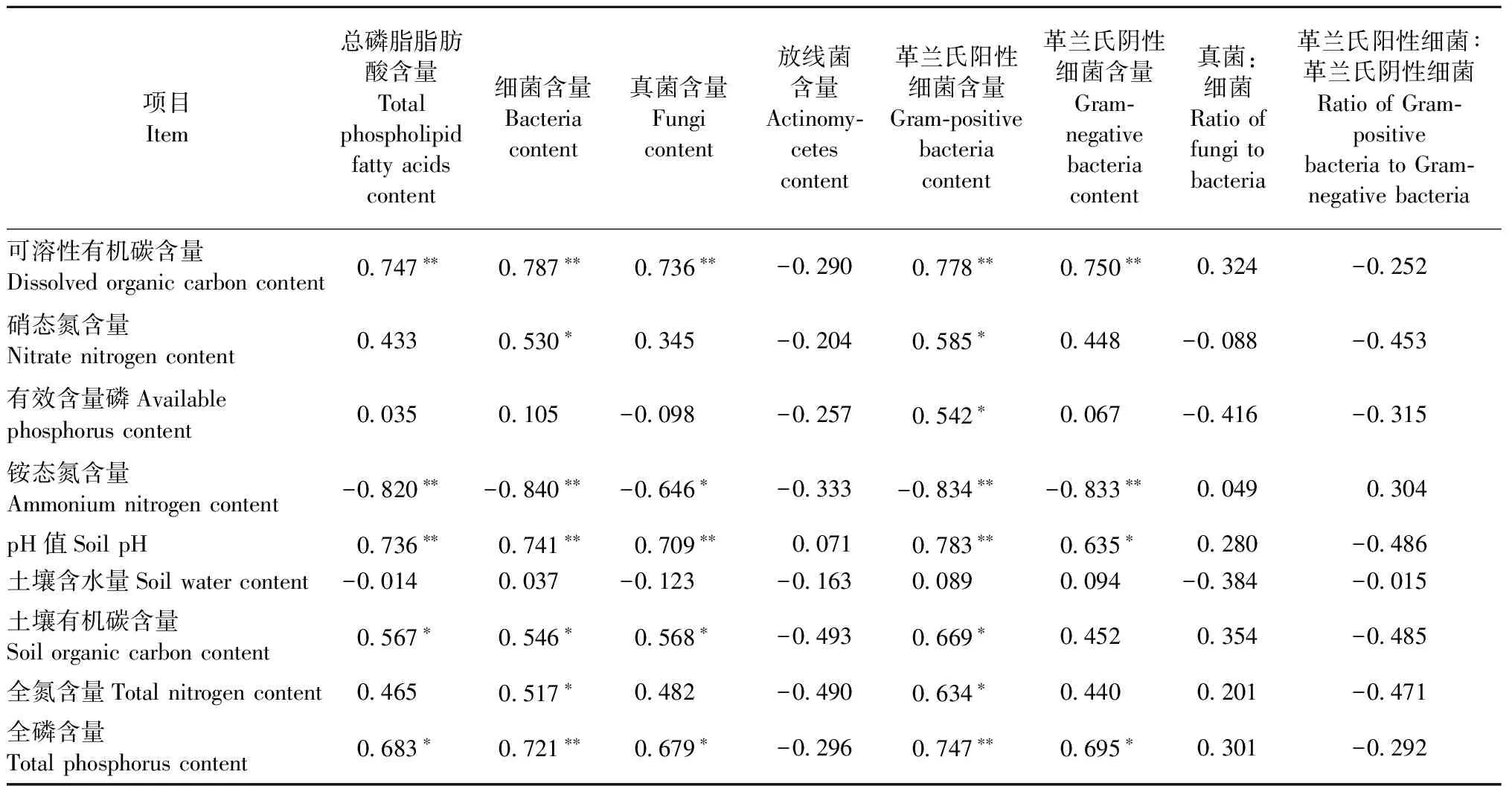
表2 土壤微生物群落結(jié)構(gòu)與土壤性質(zhì)相關(guān)性①Tab. 2 Correlations between soil microbial community composition and soil properties
①**:P<0.01; *:P<0.05.
4 討論
4.1 土壤養(yǎng)分有效性的變化
4.2 土壤微生物群落結(jié)構(gòu)的變化
本研究結(jié)果表明環(huán)割處理后,土壤總微生物生物量、真菌和細(xì)菌含量均顯著降低,表明土壤微生物對(duì)地上光合產(chǎn)物具有依懶性,而且光合碳輸入的改變對(duì)真菌群落的影響要顯著高于細(xì)菌群落。環(huán)割處理1個(gè)月顯著降低了2種人工林中真菌含量,但是并沒(méi)有對(duì)馬尾松土壤細(xì)菌群落產(chǎn)生顯著影響; 而且盡管杉木林中環(huán)割導(dǎo)致細(xì)菌生物量下降了10.9%,但真菌下降的程度(20.3%)顯著高于細(xì)菌(圖1b,c)。這主要是由于真菌比其他微生物群落更依賴(lài)于光合產(chǎn)物供應(yīng)(Keith-Roachetal., 2002),而且前人的研究也表明地下光合碳輸入的降低導(dǎo)致依賴(lài)于根系分泌物的菌根真菌數(shù)量顯著下降(H?gbergetal., 2007; 2010; Kaiseretal., 2010; Chenetal., 2012)。此外,本研究結(jié)果表明環(huán)割1個(gè)月后,杉木土壤革蘭氏陰性細(xì)菌含量顯著下降(圖1f),也說(shuō)明光合產(chǎn)物供應(yīng)的重要性,因?yàn)楦锾m氏陰性細(xì)菌更傾向于利用根系分泌的活性碳(Treonisetal., 2004)。
本研究發(fā)現(xiàn)環(huán)割對(duì)土壤微生物群落的影響具有樹(shù)種差異性。在杉木林中環(huán)割1個(gè)月后,總磷脂脂肪酸、細(xì)菌、真菌、革蘭氏陰性細(xì)菌含量和真菌∶細(xì)菌都顯著下降,而在馬尾松林中,環(huán)割1個(gè)月僅降低了真菌和革蘭氏陽(yáng)性細(xì)菌的含量。這表明短期時(shí)間內(nèi)阻斷光合產(chǎn)物供應(yīng)后,杉木林土壤微生物的響應(yīng)大于馬尾松林,這歸結(jié)于樹(shù)種本身的生理特性。杉木具有很強(qiáng)的萌芽作用,大部分根系一直保持活性,杉木在環(huán)割1年后仍有大部分樹(shù)枝是活的也支持這一觀點(diǎn)。在同樣具有萌芽作用的尾葉桉人工林中,Chen等(2010)發(fā)現(xiàn)環(huán)割6個(gè)月后其根系生物量?jī)H下降了18%,這主要是由于尾葉桉根系中存儲(chǔ)的大量淀粉使得根系在環(huán)割后很長(zhǎng)的時(shí)間都不會(huì)死亡,并且有利于促進(jìn)尾葉桉樹(shù)木萌芽(Binkleyetal., 2006)。環(huán)割減弱了植物地下碳輸入,但并沒(méi)有造成植物根系的大量死亡,而且植物活根系中儲(chǔ)存的碳更多地用于維持根系生長(zhǎng)和促進(jìn)植物萌芽,因而減少向土壤中的碳輸入(表1),導(dǎo)致微生物生長(zhǎng)受到碳限制。前人的研究表明磷脂脂肪酸cy17∶0/16∶1ω7c可以用來(lái)指示基質(zhì)可利用性的限制作用(Moore-Kuceraetal., 2007; Fengetal., 2009),該比值的升高表明基質(zhì)有效性不足增加了微生物生長(zhǎng)壓力。杉木林中顯著升高的微生物磷脂脂肪酸cy17∶0/16∶1ω7c也說(shuō)明環(huán)割1個(gè)月后微生物受到可利用性碳的限制。而馬尾松萌芽能力較弱,環(huán)割后大部分根系死亡,盡管環(huán)割降低了植物光合碳供應(yīng),但是死亡的根系為微生物提供了分解底物,增加土壤可利用性碳(表1),從而在短時(shí)間內(nèi)降低了環(huán)割對(duì)微生物的影響。然而,環(huán)割1年對(duì)馬尾松土壤微生物的影響要高于杉木林。這主要是由于環(huán)割1年后,杉木的萌芽能力逐漸減弱,因而減少了對(duì)根系碳的消耗,而且死亡的根系量增加,兩者為土壤微生物提供較充足的碳(圖1i),有利于維持微生物的生長(zhǎng)。而在馬尾松林中,環(huán)割1年后,死亡根系中的可利用性碳已被微生物消耗,而且缺乏新的碳輸入導(dǎo)致微生物受到碳限制,從而抑制了微生物生長(zhǎng)(圖1i)。
環(huán)割對(duì)土壤微生物的樹(shù)種差異性影響具有時(shí)間效應(yīng),主要是由于2個(gè)樹(shù)種的萌芽能力不同,導(dǎo)致環(huán)割后短期和長(zhǎng)期時(shí)間段內(nèi)土壤碳輸入發(fā)生變化。本研究中土壤DOC含量與微生物生物量之間的顯著正相關(guān)關(guān)系也證明了土壤可利用性碳的變化是導(dǎo)致土壤微生物發(fā)生變化的主要原因(表2)。Chen等(2012)在尾葉桉和厚莢相思人工林的研究結(jié)果也表明,由于植物光合碳供應(yīng)和死根分解碳補(bǔ)給的變化,導(dǎo)致環(huán)割對(duì)土壤微生物的影響隨時(shí)間發(fā)生變化。
5 結(jié)論
通過(guò)在亞熱帶杉木和馬尾松2種人工林中為期1年的環(huán)割試驗(yàn)發(fā)現(xiàn): 1)環(huán)割降低了土壤微生物生物量,且改變了土壤微生物群落結(jié)構(gòu),這表明土壤微生物在很大程度上依賴(lài)于光合產(chǎn)物地下碳分配,而且真菌對(duì)植物光合碳的依賴(lài)高于細(xì)菌; 2)杉木和馬尾松林土壤微生物對(duì)環(huán)割的響應(yīng)具有較大的差異,表明環(huán)割對(duì)土壤微生物群落結(jié)構(gòu)的影響具有樹(shù)種差異性,而且這種差異性在時(shí)間尺度上受到植物光合碳供應(yīng)和死根分解碳補(bǔ)給的調(diào)控。本研究表明采用環(huán)割的方法改變光合產(chǎn)物碳供應(yīng)是一種有效的方法,但是由于其對(duì)土壤微生物的影響具有樹(shù)種差異性,因此在今后的研究中應(yīng)將樹(shù)種本身的特性及測(cè)定時(shí)間考慮在內(nèi)。此外,這一結(jié)果也在一定程度上解釋了目前研究中不同生態(tài)系統(tǒng)中結(jié)果差異較大的原因。但是由于本研究中重復(fù)數(shù)量偏少,而且由于對(duì)樹(shù)木的破壞性使得小區(qū)面積也偏小,且研究時(shí)間不夠長(zhǎng)使得對(duì)環(huán)割的時(shí)間效應(yīng)的研究不是很深入,因此在今后的研究中應(yīng)加強(qiáng)此方面的研究,從而更好地探索光合產(chǎn)物供應(yīng)變化對(duì)土壤微生物群落結(jié)構(gòu)的影響,以期為研究碳循環(huán)提供更準(zhǔn)確的基礎(chǔ)數(shù)據(jù)。
方麗娜,楊效東,杜 杰. 2011. 土地利用方式對(duì)西雙版納熱帶森林土壤微生物生物量碳的影響. 應(yīng)用生態(tài)學(xué)報(bào),22(4): 837-844.
(Fang L N,Yang X D,Du J. 2011. Effects of land use pattern on soil microbial biomass carbon in Xishuangbanna. Chinses Journal of Applied Ecology,22(4): 837-844.[in Chinese])
賀同鑫,李艷鵬,張方月,等. 2015. 林下植被剔除對(duì)杉木林土壤呼吸和微生物群落結(jié)構(gòu)的影響.植物生態(tài)學(xué)報(bào), 39(8): 797-806.
(He T X,Li Y P,Zhang F Y,etal. 2015. Effects of understory removal on soil respiration and microbial community composition structure in a Chinese fir plantation.Chinese Journal of Plant Ecology,39(8): 797-806. [in Chinese])
Binkley D,Stape J L,Takahashi E N,etal. 2006. Tree-girdling to separate root and heterotrophic respiration in twoEucalyptusstands in Brazil. Oecologia, 148(3): 447-454.
Bossio D A,Scow K M,Gunapala N,etal. 1998. Determinants of soil microbial communities: effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microbial Ecology,36 (1): 1-12.
Brant J B,Sulzman E W,Myrold D D. 2006. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation. Soil Biology and Biochemistry, 38(8): 2219-2232.
Carrillo Y,Dijkstra F A,Pendall E,etal. 2012. Controls over soil nitrogen pools in a semiarid grassland under elevated CO2and warming. Ecosystems, 15(5): 761-774.
Chen D M,Zhang Y,Lin Y B,etal. 2010. Changes in belowground carbon inAcaciacrassicarpaandEucalyptusurophyllaplantations after tree girdling. Plant and Soil, 326(1/2): 123-135.
Chen D M,Zhou L X,Wu J P,etal. 2012. Tree girdling affects the soil microbial community by modifying resource availability in two subtropical plantations. Applied Soil Ecology, 53(10): 108-115.
Dannenmann M,Simon J,Gasche R,etal. 2009. Tree girdling provides insight on the role of labile carbon in nitrogen partitioning between soil microorganisms and adult European beech. Soil Biology and Biochemistry, 41(8): 1622-1631.
Feng X,Simpson M J. 2009. Temperature and substrate controls on microbial phospholipid fatty acid composition during incubation of grassland soils contrasting in organic matter quality. Soil Biology and Biochemistry, 41(4): 804-812.
Frosteg?rd ?,B??th E. 1996. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biology and Fertility of Soils, 22(1): 59-65.
H?gberg M N,Briones M J,Keel S G,etal. 2010. Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest. New Phytologist, 187(2): 485-493.
H?gberg M N,H?gberg P,Myrold D D. 2007. Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three?.Oecologia, 150(4): 590-601.
H?gberg P,Nordgren A,Buchmann,etal. 2001. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature, 411(6839): 789-792.
Kaiser C,Koranda M,Kitzler B,etal. 2010. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytologist, 187(3): 843-858.
Keel S G,Campbell C D,H?gberg M N,etal. 2012. Allocation of carbon to fine root compounds and their residence times in a boreal forest depend on root size class and season. New Phytologist, 194(4): 972-981.
Keith-Roach M J,Bryan N D,Bardgett R D,etal. 2002. Seasonal changes in the microbial community of a salt marsh, measured by phospholipid fatty acid analysis. Biogeochemistry, 60(1): 77-96.
Maier C A,Johnsen K H,Clinton B D,etal. 2010. Relationships between stem CO2efux, substrate supply, and growth in young loblolly pine trees. New Phytologist, 185(2): 502-513.
Moore-Kucera J,Dick R P. 2007. PLFA profiling of microbial community structure and seasonal shifts in soils of a douglas-fir chronosequence. Microb Ecology, 55(3): 500-511.
Mucha A P,Almeida C M R,Bordalo A A,etal. 2005. Exudation of organic acids by a marsh plant and implications on trace metal availability in the rhizosphere of estuarine sediments. Estuarine Coastal and Shelf Science, 65(1/2): 191-198.
Scott-Denton L E,Rosenstiel T N,Monson R K. 2006. Differential controls by climate and substrate over the heterotrophic and rhizospheric components of soil respiration. Global Change Biology, 12(2): 205-216.
Subke J A,Hahn V,Battipaglia G,etal. 2004. Feedback interactions between needle litter decomposition and rhizosphere activity. Oecologia, 139(4): 551-559.
Tóth J A,Lajtha K,Kotroczó Z,etal. 2007. The effect of climate change on soil organic matter decomposition. Acta Silvatica et Ligniaria Hungaric, (3): 75-85.
Trahan N A,Dynes E L,Pugh E,etal. 2015. Changes in soil biogeochemistry following disturbance by girdling and mountain pine beetles in subalpine forests. Oecologia, 177(4): 981-995.
Treonis A M,Ostle N J,Stott A W,etal. 2004. Identification of groups of metabolically-active rhizosphere microorganisms by stable isotope probing of PLFAs. Soil Biology and Biochemistry, 36(3): 533-537.
Wang Q K,He T X,Wang S L,etal. 2013. Carbon input manipulation affects soil respiration and microbial community composition in a subtropical coniferous forest. Agricultural and Forest Meteorology, 178-179(4): 152-160.
Yarwood S A,Myrold D D,H?gberg M N. 2009. Termination of belowground C allocation by trees alters soil fungal and bacterial communities in a boreal forest. FEMS Microbiol Ecology, 70(1): 151-162.
Zeller B,Liu J,Buchmann N,etal. 2008. Tree girdling increases soil N mineralisation in two spruce stands. Soil Biology and Biochemistry, 40(5): 1155-1166.
(責(zé)任編輯 于靜嫻)
Effects of Girdling on Soil Microbial Community Composition inCunninghamialanceolataandPinusmassonianaPlantations
He Tongxin1,2Sun Jianfei1Li Yanpeng2Yu Youzhi2Hu Baoqing1Wang Qingkui2
(1.KeyLaboratoryofEnvironmentChangeandResourcesUseinBeibuGulfofMinistryofEducationGuangxiKeyLaboratoryofEarthSurfaceProcessesandIntelligentSimulationGuangxiTeachersEducationUniversityNanning530001;2.HuitongExperimentalStationofForestEcology,ChineseAcademyofSciencesHuitong418307)
Cunninghamialanceolata;Pinusmassoniana; plantation; phospholipid fatty acid; girdling; microbial community composition
10.11707/j.1001-7488.20170609
2015-10-08;
2017-01-20。
中國(guó)科學(xué)院戰(zhàn)略性先導(dǎo)科技專(zhuān)項(xiàng) (XDB15010301); 國(guó)家自然科學(xué)基金項(xiàng)目(31570466); 國(guó)家重點(diǎn)研發(fā)計(jì)劃 (2016YFA0600801); 廣西科學(xué)研究與技術(shù)開(kāi)發(fā)計(jì)劃 (14125008-2-24)。
S714.3
A
1001-7488(2017)06-0077-08
*王清奎為通訊作者。
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(xiāo)(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00

- 林業(yè)科學(xué)的其它文章
- 松材線蟲(chóng)CYP450基因與松樹(shù)蒎烯類(lèi)物質(zhì)代謝的相關(guān)性*
- Relationship between Metabolites Variation and Paulownia Witches’ Broom
- 基于空間自相關(guān)的天然蒙古櫟闊葉混交林林木胸徑-樹(shù)高模型*
- 基于GWR的大興安嶺森林立地質(zhì)量遙感分析*
- 容器類(lèi)型和胚根短截對(duì)栓皮櫟容器苗苗木質(zhì)量及造林初期效果的影響*
- 基于無(wú)損檢測(cè)技術(shù)的濕地松生長(zhǎng)及材性性狀遺傳變異分析*