高通量測序在醬香白酒微生態多樣性研究中的應用
2017-06-01 12:42:08郭敏黃永光邱樹毅胡建峰胡峰鐘方達
中國釀造 2017年5期
郭敏,黃永光*,邱樹毅,胡建峰,胡峰,鐘方達
(1.貴州大學釀酒與食品工程學院,貴州貴陽550025;2.貴州大學貴州省發酵工程與生物制藥重點實驗室,貴州貴陽550025;3.貴州茅臺酒廠(集團)習酒有限責任公司,貴州習水564622)
高通量測序在醬香白酒微生態多樣性研究中的應用
郭敏1,2,黃永光1,2*,邱樹毅1,2,胡建峰3,胡峰3,鐘方達3
(1.貴州大學釀酒與食品工程學院,貴州貴陽550025;2.貴州大學貴州省發酵工程與生物制藥重點實驗室,貴州貴陽550025;3.貴州茅臺酒廠(集團)習酒有限責任公司,貴州習水564622)
通過提取醬香型白酒生產發酵過程3輪次、7輪次酒醅微生物基因組DNA,采用高通量測序技術對酒醅中的微生態多樣性進行測序分析。結果表明,醬香型白酒發酵過程3輪次酒醅(L3)中乳桿菌屬(Lactobacillus)和芽孢桿菌屬(Bacillus)是原核微生物中絕對優勢菌,而7輪次酒醅(L7)中鹽單胞菌屬(Halomonas)是原核微生物中最優勢菌屬;同時在L3樣品中嗜熱真菌屬(Thermomyces)和嗜熱子囊菌屬(Thermoascus)是發酵過程真核微生物中的絕對優勢菌,而在L7樣品中隱球菌屬(Cryptococcus)則是真核微生物中的最優勢菌屬。對比兩輪次酒醅中微生物分析結果,二者的微生態多樣性存在較大的差異。分析認為可能正是因為兩輪次酒醅中微生物在發酵過程中處于動態變化的差異性,導致兩個輪次生產出的酒在產量和質量風味上自然存在較大的差異。
醬香型白酒;酒醅;高通量測序方法;微生物多樣性
GUO Min1,2,HUANG Yongguang1,2*,QIU Shuyi1,2,HU Jianfeng3,HU Feng3,ZHONG Fangda3
(1.School of Liquor and Food Engineering,Guizhou University,Guiyang 550025,China;2.Key Laboratory of Fermentation Engineering and Biological Pharmacy of Guizhou Province,Guizhou University,Guiyang 550025,China; 3.Guizhou Moutai Brewery(Group)Xijiu Co.,Ltd.,Xishui 564622,China)
醬香型白酒是中國最具有代表性、獨特性的蒸餾酒,以其獨特的釀造工藝形成獨特的風格體系;以醬香突出、酒體醇厚、空杯留香等風格而聞名[1]。醬香型白酒生產過程發酵酒醅在堆積、窖池發酵過程富集、培養了大量微生物,微生物在酒醅中生長繁殖并產生大量代謝物和風味化合物。在醬香型白酒生產中,第3、4、5輪次生產酒醬香風格典型突出,酒體較為醇厚、豐滿,產量和質量效益優勢明顯;而第7輪次酒的生產屬于醬香型白酒釀造工藝的最后一輪,發酵底物的發酵特性處于最差狀態,微生物的群落結構也明顯與3、4、5輪次存在明顯差異,所生產基酒的風格、質量、出酒率也都相差較大。發酵酒醅中的微生物結構直接決定著酒的產質量與酒體風味,釀酒微生物主要包括細菌、真菌等,真菌包括霉菌、酵母菌等。細菌在發酵過程主要功能作用是分泌風味型酶、代謝產生風味物質、調節發酵過程微生態結構等。霉菌主要分泌糖化酶、蛋白酶等,將淀粉轉化為可發酵性的糖、分解蛋白質,提高淀粉、蛋白質的利用率。酵母菌主要包括了酒精酵母、產酯酵母等,酒精酵母主要起代謝產酒精的作用,產酯酵母具有產酯能力,能夠增加酒體的酯含量,賦予酒舒適的香氣。因此,研究醬香型白酒發酵過程的微生態多樣性及其結構對詮釋醬香型白酒獨特風味的形成具有重要的意義。
高通量測序技術(high-throughput sequencing)可以滿足對多個樣品中的微生物進行深入分析,其主要特點是通量高、速度快、準確度高、實時檢測、數據信息量大等,能在很短時間內獲取大量的數據[2-3]。目前,國內應用高通量測序技術分析研究濃香型、清香型白酒發酵酒醅中細菌或真菌微生態多樣性已有報道[4-7],且高通量測序技術也被用于研究不同發酵食品中的微生物生態,包括醋[8]和牛奶[9],而在醬香型白酒釀造領域還很少涉入。
本研究以醬香型白酒釀造的第3輪次和第7輪次的釀造酒醅為樣本,探索利用高通量測序分析酒醅中的細菌16SrDNA的V4變異區序列和真菌內的轉錄間隔區(internal transcribed spacer,ITS)序列,研究醬香型白酒釀造過程酒醅中細菌和真菌的生態結構。旨在從微生態多樣性的角度解析導致兩個輪次產酒在產量、質量和風格上存在較大差異的原因,建立高通量測序技術應用于醬香型白酒制酒微生態多樣性及其結構的研究分析方法,更加準確、完整的解析醬香型白酒釀造過程酒醅中微生物的群落結構及其主導功能,為今后全面研究醬香型白酒釀造微生態多樣性及其相關研究提供可行性的方法和奠定基礎。
1 材料與方法
1.1 材料與試劑
樣品取自茅臺集團XJ公司醬香型酒制酒車間2016年第3輪次、第7輪次生產酒醅,取后置于-20℃冰柜保存,備用。
DNA提取試劑盒:美國MP Biomedicals公司;引物由深圳市恒創基因科技有限公司合成。
1.2 儀器與設備
SpeedCycler PCR儀,德國Analytikjena公司;GS junior高通量測序儀,美國Roche公司。
1.3 方法
1.3.1 樣品處理方法
稱取10 g樣品,用15 mL滅菌后的0.1 mol/L磷酸鹽緩沖液(phosphate buffer solution,PBS)懸浮,加入適量玻璃珠,漩渦振蕩5 min,300 r/min離心5 min,取上清,沉淀用PBS緩沖液重復洗滌3次,離心后收集上清,將所有收集上清混勻。全部上清于9 000 r/min離心3 min,棄去上清,收集細胞沉淀。再用5 mL PBS洗3次,每次于9 000 r/min離心3 min,收集沉淀。
1.3.2 微生物DNA提取
采用試劑盒方法提取DNA,提取過程參照試劑盒說明書進行。
1.3.3 微生物擴增和焦磷酸測序
使用16S rDNA通用引物515F(5'-GTGCCAGCMGCC GCGG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')對細菌DNA的V4高變區進行擴增;使用真菌引物ITS1F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS2R(5'-GC TGCGTTCTTCATCGATGC-3')對真菌ITS1F-ITS2R區域進行擴增。
1.3.4 PCR反應體系
TransStartFastpfuDNA聚合酶20μL反應體系:Forward引物(5 μmol/L)0.8 μL;Reverse引物(5 μmol/L)0.8 μL;5×FastPfu Buffer 4 μL;2.5 mmol/L脫氧核糖核苷三磷酸(deoxy-ribonucleoside triphosphate,dNTPs)2 μL;FastPfu Polymerase0.4μL;TemplateDNA10ng;補ddH2O至20μL。
聚合酶鏈反應(polymerase chain reaction,PCR)擴增程序:細菌為95℃預變性5 min;95℃變性30 s;55℃復性30 s;72℃延伸45 s;共27個循環。最后72℃延伸10 min。真菌為95℃預變性5 min;95℃變性30 s;55℃復性30 s;72℃延伸45 s;共29個循環。最后72℃延伸10 min。
1.3.5 高通量測序與數據分析
采用高通量測序儀進行測序。應用QIIME軟件總體分析測序數據后再利用Greengene數據庫進行物種注釋,進一步確定序列對應微生物的分類學地位。
2 結果與分析
2.1 樣品微生物測序指標分析
對酒醅中微生物的DNA提取并PCR產物純化及定量后上機測序分析,其Shannon、Chao1指數等分析指標見表1。

表1Shannon等指標分析結果Table 1 Analysis results of Shannon and other indexes
由表1可知,在所測定2個樣本的細菌中共獲得69651條16SrDNA基因序列(其中L3為32 846,L7為36 805),真菌中共獲得34 497條ITS基因序列(其中L3為634,L7為12 459)。為了獲得樣品中微生物多樣性信息,根據97%相似性將序列聚類成為操作分類單元(operational taxonomic unit,OUT),OUT數一定程度上說明了樣品多樣性的程度。細菌的總OUT數為657(其中L3為634,L7為23),真菌的總OUT數為127(其中L3為54,L7為73),說明樣品中微生物構成中,原核微生物的復雜程度高于真核微生物。Alpha值可用來估計樣品中細菌群落物種的豐度和多樣性,包括Chao1、Shannon等分析指數,在相似度97%以上的分類水平上,利用應用軟件計算樣品的豐度和多樣性指數。Chao1指數用來衡量群落豐度的指數,隨著值的增大,微生物群落的豐度也相應較高。本研究中,樣品的Shannon曲線值分別是0.12和4.27,Chao 1指數值分別是23和669.68,細菌的測序深度為30 000,因此只需對30 000條序列進行測定分析就可以覆蓋酒醅中所有細菌。本次研究樣品中細菌多樣性分析了36 805和32 846條序列;在真菌中,樣品的Shannon曲線值分別為3.9和1.47,Chao 1指數值分別為73和64,真菌的測序深度為10000,因此也只需對10000條序列進行測定分析就可以覆蓋酒醅中所有真菌,而本次研究真菌多樣性分析了12 459和22 038條序列,說明樣品的物種多樣性和豐富度都較高。
由表1還可知,2個樣品中細菌的Chao1指數大小順序為L3>L7,說明第3輪次堆積酒醅中細菌的群落豐度比第7輪次高;比較2個樣品真菌的Chao1指數,結果表明Chao1指數大小為L7>L3,說明第7輪次堆積酒醅中真菌的群落豐度比第3輪次堆積酒醅高,這與生產實際符合。Shannon指數的大小與Chao 1指數的變化一致。測序結果表明醬香型酒醅樣品中的生物物種多樣性和豐度都比較高,所覆蓋的信息量足以真實性的表征酒醅中微生物群落結構的多樣性。
2.2 細菌多樣性分析
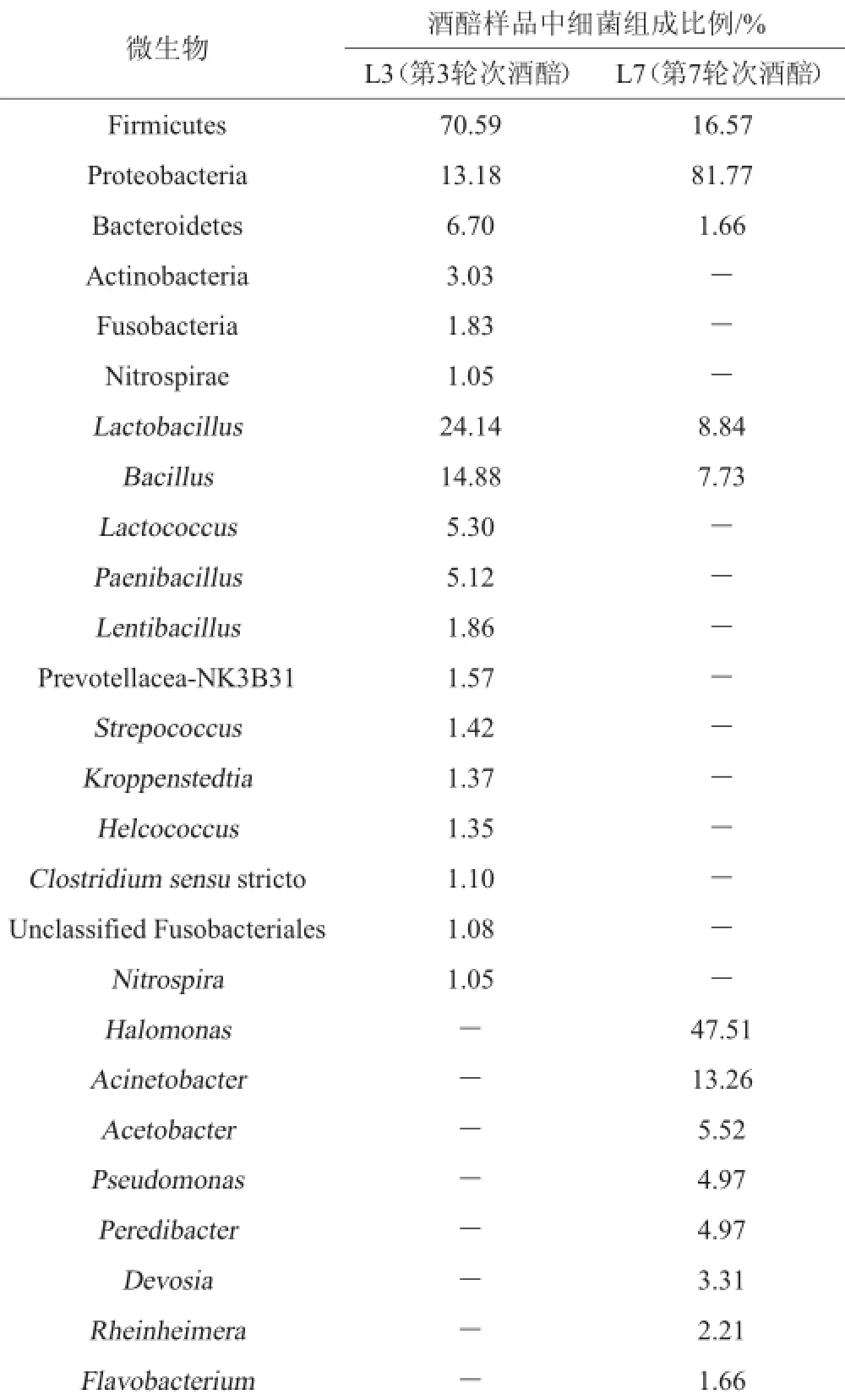
表2 樣品中細菌序列的百分含量Table 2 Percentage contents of bacterial sequences in samples
樣品中細菌菌群測序數據解析結果見表2。由表2可知,在L3樣品中厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)、梭桿菌門(Fusobacteria)、硝化螺旋菌門(Nitrospirae)為優勢菌門。在屬分類水平上,優勢菌屬分別為乳桿菌屬(Lactobacillus)24.14%、芽孢桿菌屬(Bacillus)14.88%、乳球菌屬(Lactococcus)5.30%、類芽孢桿菌屬(Paenibacillus)5.12%、Lentibacillus屬1.86%、普雷沃氏菌科-NK3B31屬(Prevotellaceae-NK3B31)1.57%、鏈球菌屬(Strepococcus)1.42%、科羅彭斯特菌屬(Kroppenstedtia)1.37%、創傷球菌屬(Helcococcus)1.35%、狹義的梭菌屬(Clostridium sensu strict)1.10%、未分類梭桿菌科(UnclassifiedFusobacteriales)1.08%、硝化螺旋菌屬(Nitrospira)1.05%。在L3樣品中特有的優勢細菌屬有:乳球菌屬(Lactococcus)、類芽孢桿菌屬(Paenibacillus)、Lentibacillus、普雷沃氏菌科-NK3B31屬(Prevotellacea-NK3B31)、鏈球菌屬(Strepococcus)、科羅彭斯特菌屬(Kroppenstedtia)、創傷球菌屬(Helcococcus)、狹義的梭菌屬(Clostridium sensu stricto)、未分類梭桿菌科(Unclassified fusobacteriales)、硝化螺旋菌屬(Nitrospira)。
由表2還可知,在L7樣品中,以厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)、擬桿菌門(Bacteroidetes)為主要優勢門。在屬的分類水平上,鹽單胞菌屬(Halomonas)47.51%是最主要的優勢屬,此外,占總序列的13.26%、8.84%、7.73%、5.52%、4.97%、4.97%、3.31%、2.21%、1.66%的序列分別屬于不動桿菌屬(Acinetobacter)、乳桿菌屬(Lactobacillus)、芽孢桿菌屬(Bacillus)、醋酸桿菌屬(Acetobacter)、假單胞菌屬(Pseudomonas)、吞菌弧菌屬(Peredibacter)、戴沃斯菌屬(Devosia)、南極適冷菌屬(Rheinheimera)、黃桿菌屬(Flavobacterium),還包括金黃桿菌屬(Chryseobacterium)、土壤桿菌屬(Agrobacterium)、纖維弧菌屬(Cellvibrio)。在L7樣品中特有的優勢細菌屬有鹽單胞菌屬(Halomonas)、不動桿菌屬(Acinetobacter)、醋酸桿菌屬(Acetobacter)、假單胞菌屬(Pseudomonas)、吞菌弧菌屬(Peredibacter)、戴沃斯菌屬(Devosia)、倫黑墨氏菌屬(Rheinheimera)、黃桿菌屬(Flavobacterium)。其中,鹽單胞菌屬(Halomonas)為最優勢菌屬(占總序列的47.51%),該菌屬在釀酒領域研究中的報道較少,唐婧[10]利用宏基因組學方法對茅臺酒酒曲中的細菌進行多樣性分析過程檢測到鹽單胞菌,該菌為耐鹽細菌,部分鹽單胞菌可應用于處理高鹽染料廢水[11]。不動桿菌屬(Acinetobacter)為條件致病菌,其耐藥性較強,對氨芐青霉素、氯霉素等抗菌藥物具有耐藥作用[12],該類菌主要為石油烴的降解者,對石油烴具有乳化和降解作用,可以降低石油烴生物毒性[13]。假單胞菌屬(Pseudomonas)于條件致病菌[14],該菌類大多可對蛋白質和脂肪進行分解;假單胞菌屬與某些高級醇類具有較好的相關性,說明該屬的細菌能生成高級醇。假單胞菌屬在釀酒領域報道較少,侯小歌等[15]以河南宋河白酒發酵30 d的糟醅為實驗材料,通過富集培養及稀釋涂布法分離厭氧及兼性厭氧菌,共分離到5株己酸菌、6株丁酸菌、2株甲烷菌、6株乳酸菌。丁酸菌、己酸菌和乳酸菌革蘭氏染色均為陽性,甲烷菌為革蘭氏陰性菌。丁酸菌和己酸菌分別歸梭菌屬(Clostridium)和芽孢桿菌屬(Bacillus),甲烷菌為假單胞菌屬(Pseudomonas),乳酸菌為芽孢乳桿菌屬(Sporolactobacillus)。乳桿菌屬(Lactobacillus)和芽孢桿菌屬(Bacillus)是醬香型白酒發酵過程的優勢細菌屬,并且乳桿菌屬(Lactobacillus)和芽孢桿菌屬(Bacillus)在L3樣品中為絕對優勢細菌,比例分別為24.14%和14.88%,而在L7樣品中含量相對較少,分別為8.84%和7.73%,這與第3輪次酒和第7輪次酒的風味差異成正相關,這一群落結構也決定了第3輪次酒和第7輪次酒的本質差異。乳酸桿菌屬廣泛存在于酒醅中,該類細菌為酒類發酵過程中的生酸微生物,可以通過同型和異型乳酸兩種發酵方式產生大量乳酸,并能產生抗菌素等物質,對釀酒過程中致病菌的生長繁殖具有抑制作用,可以促進酵母、霉菌等有益微生物的生長,對釀酒過程的微生態環境具有保護、穩定調控作用[16];芽孢桿菌是醬香型白酒生產過程中的主要產醬香功能菌,具有較強的分泌蛋白酶、淀粉酶和纖維素酶等酶類的能力,可分解大分子物質形成雙乙酰、含氮化合物等芳香物質,對醬香型白酒風味物質的形成具有重要貢獻作用。多項研究證明芽孢桿菌是醬香風味成分形成的基礎和關鍵,并將其應用于制作強化大曲和堆積過程具有增強白酒醬香風味與提高醬香型白酒質量等作用[17-18]。當前對芽孢桿菌耐高溫、產酶、產香的功能性研究較多,不同的芽孢桿菌代謝分泌的酶活力不同且功能特性也不一樣[19]。
2.3 真菌多樣性分析
醬香型白酒釀造過程真菌類微生物主要有酵母和霉菌。酵母具有產酒和產酯香功能;霉菌主要分泌糖化酶、蛋白酶等,降解淀粉、蛋白質等物質,對產酒具有積極的促進作用。同時,在生長代謝過程還產生一些呈香呈味物質,對醬香酒的風味具有一定的貢獻。樣品中真菌序列的百分含量分析結果見表3。
由表3可知,在L3樣品中子囊菌門(Ascomycota)為優勢菌門,占99.84%。在屬分類水平上,優勢菌屬分別為嗜熱霉屬(Thermomyces)40.73%、嗜熱子囊菌屬(Thermoascus)25.98%、未分類的散囊菌目(Unclassfied-eurotiales)3.16%、Rasamsonia屬4.16%、曲霉屬(Aspergillus)2.59%、絲衣霉屬(Byssochlamys)2.35%、假絲酵母屬(Candida)0.55%、紅曲霉屬(Monascus)0.17%、毛孢子菌屬(Trichosporon)0.07%、小囊菌屬(Microascus)0.06%、Trichomonascus屬0.03%、青霉屬(Penicillium)0.02%、鏈格孢霉屬(Alternaria)0.02%。在L3樣品中特有的優勢真菌屬包括嗜熱霉屬(Thermomyces)、嗜熱子囊菌屬(Thermoascus)、絲衣霉屬(Byssochlamys)、紅曲霉屬(Monascus)等。其中,嗜熱霉屬(Thermomyces)和嗜熱子囊菌屬(Thermoascus)是醬香型白酒發酵過程絕對優勢的真菌微生物且都屬于嗜熱真菌類,它們是白酒發酵環境中重要酶類的來源,大多能產生具有高活力和熱穩定性的纖維素酶、蛋白酶等,對降解高分子多糖和蛋白質具有促進作用,有利于釀酒原料被微生物利用,起到促進微生物繁殖生長以及產酒生香的作用。SHI J H等[20]研究發現,嗜熱霉菌屬為汾酒曲曲心的優勢真菌,且該屬真菌在多種不良環境中都具有較好的熱穩定性和較高的活力,表明其對醬香型白酒制酒環境的適應性較好。嗜熱子囊菌能產生熱堿穩定性較好的過氧化氫酶,該酶廣泛運用于食品消毒、臨床分析、醫學診斷以及紡織、造紙、制漿等工業[21]。

表3 樣品中真菌序列的百分比含量Table 3 Percentage contents of fungal sequences in samples
在L7樣品中主要有兩個門類微生物—子囊菌門和擔子菌門,分別占49.5%和47.1%。在屬分類水平上,隱球酵母屬(Cryptococcus)占總序列的48.29%。此外,占總序列13.42%、6.32%、4.17%、2.59%、2.35%、1.8%、1.49%、1.46%的序列分別屬于曲霉菌屬(Aspergillus)、球果傘屬(Stro-bilurus)、假絲酵母屬(Candida)、木拉克酵母屬(Mrakia)、Kazachstania屬、梗孢酵母屬(Sterigmatomyces)、鎖霉菌屬(Itersonilia)、紅酵母菌屬(Rhodotorula)。還包括鏈孢霉屬(Neurospora)、嗜熱子囊菌屬(Thermoascus)、嗜熱霉屬(Thermomyces)、許旺酵母屬(Schwanniomyces)、絲衣霉屬(Byssochlamys)等。在L7樣品中特有的優勢真菌屬有隱球酵母屬(Cryptococcus)、木拉克酵母屬(Mrakia)、Kazachstania屬、梗孢酵母屬(Sterigmatomyces)、鎖霉菌屬(Itersonilia)、紅酵母菌屬(Rhodotorula)。其中,隱球酵母屬(Cryptococcus)為最優勢菌屬,關于隱球酵母屬在醬香型酒醅中的報道較少,大多報道表明隱球酵母屬具有較強的產酯能力[22],并且作為非釀酒酵母對醇類的形成影響也較大,并在酒精發酵中發揮重要作用,具體作用尚未明確,正在對隱球酵母屬在醬香型白酒中的作用機制做進一步的研究。
曲霉屬(Aspergillus)和假絲酵母屬(Candida)是醬香型白酒發酵過程中的優勢真菌屬,曲霉是釀酒過程中的一大類微生物,是釀造霉菌中種類最多的菌。由于醬香型白酒釀造環境偏酸性,曲霉分泌以酸性水解酶類為主,如酸性淀粉酶、酸性蛋白酶等,這些酶不僅耐乙醇,還具有耐酸特性,使曲霉能在酸性釀造環境條件下正常生長代謝,分解原料中的淀粉與蛋白質,為發酵提供持續性的動力[23]。假絲酵母是白酒釀造發酵過程中的主要產酒酵母,對醬香酒風味形成和感官品質具有重要影響。目前,在多種發酵類食品中均有報道,例如酒曲[24]、奶酪[25]等,該屬真菌可以產生大量的酯類化合物,例如乙酸乙酯、乙酸異戊酯等酯類物質[26]等,可為曲坯增加濃郁的酯香,此外假絲酵母還具有較強的產2-丁酮的能力[27]。劉蕓雅[28]對黃酒中的假絲酵母屬真菌與發酵中有機酸的相關性進行研究,結果表明在白酒發酵過程中該菌類對形成有機酸具有重要作用,并對酒體風格的形成具有重要影響。
3 結論
本研究使用高通量測序方法對酒醅樣品中微生物進行了分析研究。結果表明,在醬香型白酒發酵過程中,在第3輪次樣品中乳桿菌屬(Lactobacillus)和芽孢桿菌屬(Bacillus)占有絕對優勢,而在第7輪次樣品中鹽單胞菌屬(Halomonas)為最優勢菌屬;同時在第3輪次樣品中,嗜熱霉屬(Thermomyces)和嗜熱子囊菌屬(Thermoascus)是發酵過程真菌微生物中的絕對優勢菌,而在第7輪次樣品中隱球酵母屬(Cryptococcus)則為最優勢菌屬。對比第7輪次和第3輪次酒醅中微生物的分析結果,二者在微生態多樣性上存在較大的差別。分析認為可能正是因為第3輪次和第7輪次的酒醅中的微生物在白酒發酵過程中處于動態變化的差異性,導致兩個輪次生產出的白酒在產量和風味質量上存在較大的差異。
高通量測序技術與醬香型白酒釀造過程微生物研究的傳統方法相比較,由于傳統微生物技術的局限性,只能對酒醅中少量的功能微生物進行研究,遠不能滿足對復雜的微生物群落結構及其多樣性研究的需求[20,29];而高通量測序技術從基因組的水平上解析微生物群落結構,對于研究醬香型白酒生產過程中的微生物多樣性結構分析更加全面,檢測獲取的生物信息量更大,更能說明微生物結構的真實性,顯示出高通量技術在醬香型白酒生產中對復雜微生物生態結構研究中的明顯優勢[30-32]。因此該研究為探究白酒釀造微生物群落結構特征提供了一種可借鑒的方法,可為進一步研究醬香型酒醅中功能微生物提供參考。參考文獻:
[1]沈怡方.白酒生產技術全書[M].北京:中國輕工業出版社,1998:135-143.
[2]LI R,ZHU H,RUAN J,et al.De novo assembly of human genomes with massivelyparallel short read sequencing[J]. Genom Res,2010,20(2):65-72.
[3]YANG F,ZENG X,NING K,et al.Saliva microbiomes distinguish caries-active from healthy human populations[J]. ISME J,2012,6(1): 1-10.
[4]SUN W N,XIAO H Z,PENG Q,et al.Analysis of bacterial diversity of Chinese Luzhou-flavor liquor brewed in different seasons by Illumina Miseq sequencing[J].Ann Microbiol,2016,66(3):1293-1301.
[5]LIU M K,ZHAO K,TANG Y M,et al.Analysis of clostridium cluster i community diversity in pit mud used in manufacture of Chinese Luzhouflavor liquor[J].Food Sci Biotechnol,2015,24(3):995-1000.
[6]LI X R,MA E B,YAN L Z,et al.Bacterial and fungal diversity in the starter production process of fen liquor,a traditional Chinese liquor[J].J Microbiol,2013,51(4):430-438.
[7]LIANG H P,LI W F,LUO Q C,et al.Analysis of the bacterial community in aged and aging pit mud of Chinese Luzhou-flavour liquor by combined PCR-DGGE and quantitative PCR assay[J].J Sci Food Agr,2014, 95(13):2729-2735.
[8]LI P,LI S,CHENG L L,et al.Analyzing the relation between the microbial diversity of Daqu and the turbidity spoilage of traditional Chinese vinegar[J].Appl Microbiol Biotechnol,2014,98(13):6073-6084.
[9]LIU W J,ZHENG Y,KWOK L Y,et al.High-throughput sequencing for the detection of the bacterial and fungal diversity in Mongolian naturally fermented cow's milk in Russia[J].BMC Microbiol,2015,15(1):45.
[10]唐婧,蘇迪,徐小蓉,等.基于宏基因組學的茅臺酒酒曲細菌的多樣性分析[J].貴州農業科學,2014(11):180-183.
[11]田存萍.鹽單胞菌株在高鹽染料廢水處理中的應用研究[D].大連:大連理工大學,2006.
[12]不動桿菌屬、莫拉菌屬、金黃桿菌屬常規鑒[C].海口:海南省衛生廳醫院內感染監測中心,2007:6.
[13]劉玉華,王慧,胡曉珂.不動桿菌屬(Acinetobacter)細菌降解石油烴的研究進展[J].微生物學通報,2016,43(7):1579-1589.
[14]梁偉,李娟.非發酵革蘭陰性桿菌臨床耐藥性分析[J].當代醫學,2011,17(29):127-128.
[15]侯小歌,王俊英,李學思,等.宋河白酒發酵糟醅中厭氧及兼性厭氧菌的分離與鑒定[J].周口師范學院學報,2013,30(2):65-69.
[16]LV X C,WENG X,ZHANG W,et al.Microbial diversity of traditional fermentation starters for Hong Qu glutinous rice wine as determined by PCR-mediated DGGE[J].Food Control,2012,28(2):426-434.
[17]楊國華,邱樹毅,黃永光.醬香白酒生產中產香微生物研究[J].中國釀造,2011,30(4):24-27.
[18]馬榮山,劉婷,郭威.麩曲醬香酒醅中酵母菌的分離、篩選及應用[J].中國釀造,2008,27(1):17-18.
[19]李習.高產α-淀粉酶地衣芽孢桿菌的篩選及重組質粒的構建[D].武漢:湖北工業大學,2014.
[20]SHI J H,XIAO Y P,LI X R,et al.Analyses of microbial consortia in the starter of Fen liquor[J].Lett Appl Microbiol,2009,48(4):478-485.
[21]方芳,李寅,堵國成,等.一株嗜熱子囊菌產生的堿性耐熱過氧化氫酶及其應用潛力[J].生物工程學報,2004,20(3):423-428.
[22]相光明,趙祥穎,劉建軍.高產油脂隱球酵母菌發酵條件研究[J].糧食與油脂,2009(3):17-20.
[23]孫劍秋,劉雯雯,臧威,等.醬香型白酒酒醅中霉菌群落組成與功能酶活性[J].中國食品學報,2013,13(8):239-247.
[24]王海燕.PCR-DGGE技術對清香型汾酒微生物群落結構演變規律的研究[D].無錫:江南大學,2014.
[25]GOLIC N,CADEZ N,TERZIC-VIDOJEVIC A,et al.Evaluation of lactic acid bacteria and yeast diversity in traditional white pickled and fresh soft cheeses from the mountain regions of Serbia and lowland regions of Croatia[J]. Int J Food Microbiol,2013,166(2):294-300.
[26]王曉丹,龐博,陳孟強,等.醬香白酒酒醅中產香酵母分離與鑒定[J].食品安全質量檢測學報,2014,5(6):42-46.
[27]GUESSAN N,FLORENT K,DRI N,et al.Saccharomyces cerevisiae andCandida tropicalisas starter cultures for the alcoholic fermentation of tchapalo,a traditional sorghum beer[J].World J Microbiol Biotechnol,2010,26(4):693-699.
[28]劉蕓雅.紹興黃酒發酵中微生物群落結構及其對風味物質影響研究[D].無錫:江南大學,2015.
[29]黃永光,徐巖.醬香白酒釀造環境曲霉的分離及Aspergillums hennebergii酶分泌脅迫條件研究[J].食品與生物技術學報,2015,32(8):15-22.
[30]HUANG Y G,WU Q,XU Y.Isolation and identification of a blackAspergillumsstrain and the effect of its novel protease on the aroma of Moutai-flavoured liquor[J].J I Brewing,2014,120(3):268-276.
[31]鄧杰.基于高通量測序的濃香型白酒窖泥微生物群落結構研究[D].自貢:四川理工學院,2015.
[32]雷振河.采用高通量測序技術分析清香型白酒釀造微生物[J].食品與發酵工業,2015,41(9):164-167.
TS261.1
0254-5071(2017)05-0146-06
10.11882/j.issn.0254-5071.2017.05.031
2017-02-09
貴州省工業攻關項目(黔科合GZ字[2011]3015);貴州省科學技術廳重大專項(黔科合重大專項字[2012]601-5,黔科合重大專項字[2015]6012);校立項目(研理工2017020)
郭敏(1992-),女,碩士研究生,研究方向為醬香型白酒微生物。
*通訊作者:黃永光(1976-),男,研究員,博士,研究方向為醬香型白酒。
