向日葵分枝和株高性狀的關聯分析定位研究
2017-05-15 08:23:33劉勝利柳延濤王沛政
海南熱帶海洋學院學報 2017年2期
劉勝利,段 維,王 鵬,柳延濤,王沛政
(1.新疆農墾科學院,新疆 石河子 832000; 2.海南熱帶海洋學院 生命科學與生態學院,海南 三亞 572022;3.新疆康地種業科技股份有限公司,烏魯木齊 830011)
向日葵分枝和株高性狀的關聯分析定位研究
劉勝利1,段 維3,王 鵬1,柳延濤1,王沛政2
(1.新疆農墾科學院,新疆 石河子 832000; 2.海南熱帶海洋學院 生命科學與生態學院,海南 三亞 572022;3.新疆康地種業科技股份有限公司,烏魯木齊 830011)
經過structure軟件分析,128份向日葵自交系材料基因型依據△K的變化,可分為2大類群.標記間的連鎖不平衡分析顯示,SSR標記間的r2值范圍在0-1之間,平均為0.02;在選用D′來計算標記位點間的連鎖不平衡時, 25%的成對位點組合標記中存在較高水平的連鎖不平衡(D′>0.4).關聯分析定位到 8個標記位點與向日葵分枝性狀相關聯,其中有6個位點p<0.001,分別是ORS679、ORS 227-1、ORS16、 ORS22、 ORS33和ORS38;解釋表型變異8%-16%不等,2個標記位點p<0.01,分別是標記ORS803和ORS 227-2;在株高性狀中,定位到3個位點與其顯著關聯(p<0.01),分別是ORS679,ORS9、ORS22,解釋表型變異4%-6%不等.該研究成果將會為今后利用分子標記技術在向日葵新品種選育方法上提供支持.
向日葵;群體結構; 關聯分析
0 引言
向日葵(Helianthus annuus L.)是特別適合于我國西北部干旱地區種植,具有耐鹽堿、耐瘠薄、抗旱、適應性強的農作物[1].油用向日葵籽實含油率較高,亞油酸含量在60%以上,目前為世界第4大油料作物[2].當前國內向日葵品種與國外品種綜合農藝性狀相比還有一定差距,主要原因是當前我國向日葵品種科研育種基礎研究還比較薄弱[3].我國向日葵育種主要采用常規育種,向日葵常規育種方法存在著盲目性大、周期長、效率低的問題,尤其當對多基因控制的數量性狀進行育種時,容易導致偏離預期的育種目標.近幾年,分子標記技術已發展成為提高向日葵育種工作效率的強有力武器,成為當前向日葵生物學研究的熱點.
向日葵的分枝農藝性狀是在向日葵生長初期,由向日葵葉腋處的腋芽萌發,形成分枝.向日葵多頭分枝性狀主要由單顯性基因控制、單隱性基因控制和雙隱性基因控制,在不同材料中,多頭分枝性狀可能受到不同基因的調控[4].向日葵株高主要是基因加性遺傳效應控制,也受非加性效應影響,屬于較復雜的數量性狀遺傳,受多基因調控外,環境條件一定程度影響遺傳表達[5-7].
在向日葵多頭和分枝定位研究方面,文獻[8]報道了利用RFLP標記,在F2群體中將向日葵多頭基因b1定位在LG 7上.文獻[9]利用了重組自交系群體將向日葵多頭基因定位于兩個SSR標記(ORS1088, ORS930)之間.文獻[10]則利用F2群體將多頭基因定位在標記TBR11-107和TBR4-720/TBRB-555 之間.文獻[11]指出了向日葵矮化突變基因由半顯性等位基因Rht1所控制,并將其定位到連鎖群12上.
綜上所述,以上向日葵分枝和株高性狀的基因定位研究均使用單一親本的雜交分離群體,研究所用標記也不盡相同,缺乏對向日葵整個群體資源材料進行數量性狀的篩選定位研究,因此研究所得出的結果有較大的局限性.由于不同研究者使用的分子標記不同,所得的研究結論在不同實驗室間也較難進行比較,限制了其在向日葵育種中的有效應用.利用關聯分析方法進行數量性狀基因定位研究周期短、分辨率高,可以同時對多個基因進行篩選,確定不同種質資源中所具有的等位基因及其對目標的貢獻,是目前研究復雜數量性狀基因變異的常用方法.影響向日葵分枝和株高性狀的等位基因變異在其基因組中廣泛分布,因此利用關聯分析方法可以有效地鑒定出這些功能等位基因的位點.本研究利用向日葵自交系為材料,利用隨機篩選的向日葵分子標記,分析向日葵資源材料自交系的遺傳結構.在此向日葵遺傳結構的基礎上,對向日葵多頭分枝和株高性狀進行關聯分析定位.該研究結果將為今后向日葵親本選育提供可靠的分子標記,加速向日葵自交系的培育,節約育種時間.同時也可以為今后該基因的圖位克隆打下良好的基礎.
1 材料與方法
1.1 試驗材料
向日葵不同自交系種子材料由新疆農墾科學院作物所提供.為了盡可能使研究的向日葵品系具有較高的遺傳多樣性,我們收集了128份向日葵油葵和食葵各種自交系(見表1),包括油葵保持系30份和恢復系43份;食葵保持系20份和恢復系35份.田間種植試驗株距 25 cm,行距 70 cm,栽培密度 1.4×104株/hm2.小區行長 6 m,4 行區,采用完全隨機區組設計,3 次重復,生育期田間管理同當地大田生產.開花期調查材料的分枝情況,收獲期測量株高.

表1 向日葵自交系材料

續表1
1.2 DNA提取、PCR擴增和聚丙烯酰胺凝膠電泳
DNA提取:供試材料種植于海南三亞農墾科學院試驗田.取新鮮葉片,當天取樣后直接提取基因組DNA.DNA提取采用百泰克新型快速植物基因組DNA提取試劑盒(離心柱型),最后得到的DNA樣品用TE(Tris-EDTA)緩沖液溶解,放在-20℃的冰箱內保存.
TaqDNA聚合酶、dNTPs、Marker ladder、溴酚藍、丙烯酰胺、甲叉雙丙烯酰胺和TEMED等主要生化試劑購自海南相關生物銷售商,為國產分析純.
SSR引物:SSR引物見文獻[12].在該文報道的向日葵序列標簽位點(sequence tagged site)序列中,隨機選取500對SSR引物,由上海生物工程公司合成引物,首先對16份向日葵自交系材料進行擴增篩選多態.
PCR 擴增、PCR反應程序以及變性聚丙烯酰胺凝膠參照有關文獻[13].
1.3 群體結構分析
應用STRUCTRE軟件,對供試群體進行基于數學模型的類群劃分.先設定群體數目(K)為2-10,5次重復,將MCMC設為100000次,其參數設定參見已發表文獻[14].
1.4 關聯分析
在預估群體的 K 值范圍,獲得穩定可靠的群體分類結果.在此基礎上以各個體 Q 值作為協變量進行群體矯正,將向日葵分枝和株高性狀的表型數據分別對標記變異進行回歸分析,評價標記等位變異的平均效應,依據顯著水平(p<0.05和p< 0.01) ,選取貢獻率高的主要位點[15].
2 結果分析
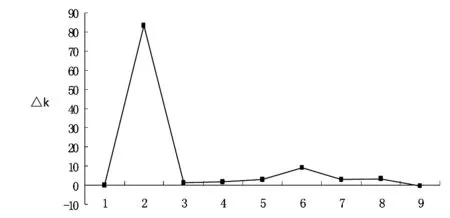
圖1 利用△K對向日葵群體結構的優化的雙邊圖表
2.1 向日葵資源材料的群體結構分析
128份向日葵資源材料群體基因型經過structure軟件運行后,結果表明供試群體的等位變異頻率特征類型數K=2時ΔK值最大(見圖1),而且差異顯著.在K為3-9之間,其△K基本降低到0附近,近似一條直線.因此依據△K的變化,向日葵自交系材料群體可以分為2大類群.
2.2 向日葵群體標記間的連鎖不平衡分析
向日葵群體標記間的連鎖不平衡分析顯示,對于全部群體SSR標記間的r2值的范圍在0-1之間,平均為0.02;而D′值在0-1之間,D′平均值達0.26.在選用D′來計算標記位點間的連鎖不平衡時,共有1320個SSR標記成對位點組合中存在連鎖不平衡,占所有標記可能成對位點組合的99%.其中25%的標記成對位點組合中存在較高水平的連鎖不平衡(D′>0.4),其中統計概率p<0.01的組合有67對,占5%.
2.3 向日葵群體分枝和株高性狀關聯分析及其等位變異表型效應
向日葵開花期調查統計結果表明,所研究的向日葵群體共有84份材料表現為不分枝,44份材料具有分枝性狀.收獲期向日株高性狀統計表明,株高為0.4米的有8份材料,0.5米的有6份材料,0.6米的有5份材料,0.7米的有2份材料, 0.8米的有26份材料, 0.9米的有6份材料, 1米的有49份材料, 1.1米的有15份材料, 1.2米的有7份材料, 1.3米的有4份材料,所有株高性狀呈現極端株高類型數目少,中等株高類型的材料占多數.
利用structure軟件中得到的各向日葵個體相應的Q值作為協變量,分別對向日葵分枝和株高性狀的表型值和標記變異進行回歸性分析,計算其相關聯的位點及其等位變異.結果顯示:在所檢測的利用62對ORS標記位點中,共有9個位點與性狀相關(見表2).其中8個位點與向日葵分枝性狀相關聯,其中有6個位點顯著關聯(p<0.001),分別是ORS679、ORS 227-1、ORS16、 ORS22、 ORS33和ORS38;解釋表型變異8%-16%不等,6個位點顯著關聯(p<0.01),分別是ORS803、ORS 227-2;在株高性狀狀中,有3個位點顯著關聯(p<0.01),分別是ORS679,ORS9、ORS22,解釋表型變異4%-6%不等.

表2 與性狀相關聯(p<0.01和p<0.001)的標記位點及其表型變異解釋率
3 討論
向日葵多頭植株花期較長,在向日葵雜交種制種過程中常作為父本,可以解決單頭親本間花期難于相遇的問題,顯著提高向日葵雜交制種的產量.因此在向日葵種質資源研究中向日葵多頭分枝性狀是很重要的農藝性狀.文獻[9]將多頭性狀定位到第10號染色體上的ORS1088和ORS908之間.本研究定位到8個位點與向日葵分枝性狀相關聯,分別是ORS679、ORS 227-1、ORS16、ORS22、ORS33、ORS38、ORS803、ORS 227-2,解釋表型變異8%-16%不等,和已發表的向日葵圖譜比對只有ORS679位于17號染色體,ORS803定位于16號染色體,其它標記未能與向日葵已知的連鎖群比對上.本研究中的關聯分析表明在所研究的向日葵資源材料中,向日葵分枝性狀由多數基因位點控制,一些定位到的向日葵分枝位點可能是以前文獻所沒報道過.向日葵莖干倒伏給向日葵生產造成嚴重的損失.然而降低植株高度將有助于增強莖干強度,減少倒伏,從而提高產量,因此現在育種的策略就是開發矮桿的種質資源,以利于其在育種中應用.本研究在株高性狀狀態中,有3個位點顯著關聯(p<0.01),分別是標記ORS679,ORS9、ORS22,解釋表型變異4%-6%不等.本研究檢測到的株高關聯位點不多,主要原因可能還是株高是由復雜數量性狀基因控制,同時所研究的分子標記數量還不夠多,因此在今后研究工作當中應繼續增加標記數量.
[1]崔良基,劉悅,王德興.我國發展向日葵生產潛力及對策 [J].雜糧作物,2008,28(5):336-338.
[2]羅偉強.氣相色譜法測定葵花籽油的脂肪酸[J].食品工業科技, 2003, 24(6):79-80.
[3]張明.國內外向日葵育種概況及動向[J].黑龍江農業科學, 2010(6):149-151.
[4]Hockett E A, Knowles P F.Inheritance of branching in sunflowers,HelianthusannuusL[J].Crop Sci .1970,10:432-436.
[5]王貴,劉學文,韓英.向日葵不育系恢復系主要性狀配合力分析[J].黑龍江農業科技,1989(4):12-16.
[6]Chandra B S, Kumar S S, Ranganadha A R G, Dudhe M Y.Combining ability studies for development of new hybrids over environments in sunflower(HelianthusannuusL.)[J].Journal of Agricultural Science, 2011,3(2):230-237.
[7]Andarkhor S A, Mastibege N, Rameeh V.Combining ability of agronomic traits in sunflower (HelianthusannuusL.) using LinexTester analysis[J].International Journal of Biology,2011,4(1):89-95.
[8]Gentzbittel L, Mestries E, Mouzeyar S, Mazeyrat F, Badaoui S, Vear F, de Labrouhe DT, Nicolas P.A Composite map of expressed sequences and phenotypic traits of the sunflower (HelianthusannuusL.) genome[J].Theoretical and Applied Genetics, 1999,99(99):218-234.
[9]Tang S, Leon A, Bridges WC, Knapp SJ.Quantitative trait loci for genetically correlated seed traits are tightly linked to branching and pericarp pigment loci in sunflower.Crop Science, 2006, 46(2):721-734.
[10]Rojas-Barros P, Hu J, Jan C.Molecular mapping of an apical branching gene of cultivated sunflower (HelianthusannuusL.) [J].Theoretical and Applied Genetics, 2008, 117(1):19-28.
[11]Ramos M L, Altieri E, Bulos M, Sala C A.Phenotypic characterization, genetic mapping and candidate gene analysis of a source conferring reduced plant height in sunflower[J].Theoretical and Applied Genetics,2013, 126(1): 251-263.
[12]Tang S, Yu J K, Slabaugh B, et al.Simple sequence repeat map of the sunflower genome[J].Theoretical and Applied Genetics.2002,105 (8):1124-1136.
[13]夏春蘭,衛澤,張雪鈺,等.改良測序聚丙烯酰胺凝膠和普通聚丙烯酰胺凝膠技術在玉米分子標記中的應用研究[J].瓊州學院學報, 2014, 21(5): 64-68.
[14]張俊,王靜毅,陳友,等.基于模型的香蕉種質資源群體結構聚類分析及其親緣關系分析[J].熱帶作物學報, 2014,35(2):232-238.
[15]文自翔,趙團結,鄭永戰,等.中國栽培和野生大豆農藝及品質性狀與 SSR 標記的關聯分析II優異等位變異的發掘[J].作物學報, 2008,34(8):1339-1349.
(編校:李由明)
Correlation Analysis on the Branches and Plant Height of the Sunflower
LIU Sheng-li1, DUAN Wei1, WANG Peng1,LIU Yan-Tao1,WANG Pei-zheng2
(1.Xinjiang Academy of Agricultural Reclamation Science, Shihezi Xinjiang 832000, China;2.School of Life Science and Ecology, Hainan Tropical Ocean University, Sanya Hainan 572022, China;3.Xinjiang Kangdi Seed Science & Technology Co.Ltd, Urumqi 830011, China)
According to the variety of △K, 128 sunflower inbred lines, analyzed by using the structure software, were divided into 2 groups.The Linkage disequilibrium between the markers shows that r2value is in the range of 0-1 with an average of 0.02.When D′ was applied to calculate the linkage disequilibrium between loci, 25% of the marker paired point combination has a high level of linkage disequilibrium (D′>0.4).Result of the correlation analysis showed that 8 marker loci were found to correlate with branch traits, of which, 6 loci—ORS679, ORS 227-1,ORS16, ORS22, ORS33 and ORS38 with phenotypic variation ranging from 8%-16%—arep<0.001, 2 loci—ORS 803 and 227-2 arep<0.01, 3 loci—ORS679, ORS9, ORS22—are signif-icantly associated with the traits of plant height withp<0.01, with their phenotypic variation ranging from 4%-6%.The collection of the identified candidate loci provides a pool of promising for the future application of molecular marker technology in the sunflower breeding.
sunflower; population structure; correlation analysis
格式:劉勝利,段維,王鵬,等.向日葵分枝和株高性狀的關聯分析定位研究[J].海南熱帶海洋學院學報,2017,24(2):80-84+95.
2016-09-15
新疆兵團現代農業科技攻關與成果轉化項目(2016AC024,2016AC027);三亞市院地科技合作項目(2013YD41)
劉勝利(1967-),男,山東寧津人, 新疆農墾科學院研究員,研究方向為向日葵新品種選育與栽培推廣.
王沛政(1972-),男,陜西長安人,海南熱帶海洋學院熱帶生物與農學院副教授,博士,研究方向為分子遺傳育種.
S565.5
A
2096-3122(2017) 02-0080-05
10.13307/j.issn.2096-3122.2017.02.16
