不同溫度對不同基因型棉花葉片葉綠素?zé)晒獾挠绊?/h1>
2017-04-26 03:09:34邢雙濤李志博湯麗魁趙瑞海魏亦農(nóng)
新疆農(nóng)業(yè)科學(xué) 2017年3期
邢雙濤,李志博,湯麗魁,趙瑞海,魏亦農(nóng)
(石河子大學(xué)農(nóng)學(xué)院/新疆生產(chǎn)建設(shè)兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室,新疆石河子 832000)
不同溫度對不同基因型棉花葉片葉綠素?zé)晒獾挠绊?/p>
邢雙濤,李志博,湯麗魁,趙瑞海,魏亦農(nóng)
(石河子大學(xué)農(nóng)學(xué)院/新疆生產(chǎn)建設(shè)兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室,新疆石河子 832000)
【目的】在不同溫度條件下,研究不同基因型棉花葉片葉綠素?zé)晒馓匦灾笜?biāo)的變化,為耐高溫棉花品種的篩選提供理論依據(jù)。【方法】分別在25、30、35、40、45和48℃的恒溫培養(yǎng)箱處理不同基因型棉花13-1品系、新海24號、13-1×新海24號(雜交種)、13-1×新海14號(雜交種),利用快速熒光測定儀(Handy PEA-100)測定不同基因型棉花葉片熒光特性主要參數(shù)。【結(jié)果】隨著溫度不斷升高,性能指數(shù)PIABS、最大光化學(xué)Fv/Fm、電子傳遞產(chǎn)額φEo表現(xiàn)為先升高后降低,而熱耗散DIO/CSO、初始熒光FO則先降低后升高;性能指數(shù)PIABS在40℃時(shí)4個(gè)不同棉花基因型棉花品系呈極顯著,比Fv/Fm更準(zhǔn)確地反映光合狀況。【結(jié)論】利用隸屬函數(shù)法分析4種不同基因型棉花品種(系),其中13-1×新海24號隸屬函數(shù)值最高,屬于耐熱品系。
棉花;溫度;最大光化學(xué)效率;性能指數(shù);葉綠素?zé)晒?/p>
0 引 言
【研究意義】光合作用在植物生長的過程中發(fā)揮著重要作用,且受環(huán)境的影響較大。高溫脅迫會導(dǎo)致光合作用受到抑制,影響光合效率[1]。棉花雖然是喜溫作物,但在生長發(fā)育與生殖發(fā)育過程中遇到35℃以上高溫,就會影響棉花得正常生長[2]。利用快速葉綠素?zé)晒鈨x來研究植物光合過程中光能利用,及外界環(huán)境因子對其影響,已成為作物逆境脅迫研究中的熱門技術(shù)[3]。因此研究不同基因型棉花葉片在不同高溫度下的葉綠素?zé)晒鈪?shù)變化規(guī)律,為鑒定棉花耐高溫材料提供依據(jù)。【前人研究進(jìn)展】董文慶等[4]發(fā)現(xiàn)棉花在不同生育時(shí)期受到高溫脅迫時(shí),不同的棉花品種熒光指標(biāo)Fv/Fm、FO、PIABS在蕾期差異最顯著,蕾期用Fv/Fm、FO、PIABS指標(biāo)量棉花光合耐熱性,性能指數(shù)PIABS比其它指標(biāo)更敏感地反映出棉花耐熱性。隨著溫度持續(xù)升高(25~50℃),對兩個(gè)不同基因型小麥光系統(tǒng)Ⅱ的能量熱耗散DIO/CSO差異不顯著,用于QA-以后的電子傳遞能量比例φEo表現(xiàn)顯著降低,初始熒光FO顯著升高,最大光化學(xué)效率Fv/Fm、性能指數(shù)PIABS均顯著下降[5]。杜國棟等(2011)[6]發(fā)現(xiàn)在高溫脅迫下,對仁用杏葉片放養(yǎng)復(fù)合體、PSⅡ反應(yīng)中心造成破壞,而且在50℃高溫導(dǎo)致初始熒光FO顯著升高,而Fv/Fm、PIABS分別顯著下降,導(dǎo)致高溫?fù)p害了PSⅡ受體側(cè)和供體側(cè)的功能,使光合機(jī)構(gòu)受害主要機(jī)制之一。【本研究切入點(diǎn)】目前相關(guān)研究僅在陸地棉品種中進(jìn)行,對于棉花不同的品種類型的研究相對較少。通過比較不同基因型品種及雜交種,進(jìn)一步研究棉花耐熱的生理機(jī)制。【擬解決的關(guān)鍵問題】在不同溫度條件下,研究不同基因型棉花葉片葉綠素?zé)晒馓匦灾笜?biāo)的變化,為進(jìn)一步選育耐高溫的棉花品種提供理論依據(jù)。
1 材料與方法
1.1 材 料
試驗(yàn)材料為13-1品系、新海24號、13-1×新海24號(雜交種)、13-1×新海14號(雜交種),以上品種(系)均由綠洲生態(tài)重點(diǎn)實(shí)驗(yàn)室育種室提供。品種(系)種植在新疆兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室培養(yǎng)溫室。
1.2 方 法
1.2.1 試驗(yàn)設(shè)計(jì)
采取盆栽試驗(yàn),以基質(zhì)和蛭石按1∶1比例混合裝進(jìn)直徑30 cm,高25 cm的花盆,每盆種3株,每個(gè)品種類型3盆。放在溫室生長,定期澆適量的水,溫室的溫度20℃,棉花長到5~6片葉時(shí),開始放到恒溫培養(yǎng)箱處理,光照強(qiáng)度3 000 lx,濕度65%。分別以25、30、35、40、45和48℃的溫度處理8 h,處理之后測定棉花熒光特性。
1.2.2 熒光特性測定
采用英國Hansatech公司的植物效率分析儀(Handy PEA-100)對棉花葉片進(jìn)行OJIP曲線測定,曲線由3 000 μmol/(m2·s)的飽和光誘導(dǎo),測定前用葉夾暗適應(yīng)15 min,從曲線得到相應(yīng)參數(shù),最大光化學(xué)效率(在t=0時(shí)),F(xiàn)v/Fm=φPo≡TRO/ABS=1-(FO/Fm);吸收的光量子將電子傳遞QA-下游的其它電子受體的概率(在t=0時(shí)),φEo≡ETO/ABS= [1-(FO/Fm)]·ψO;FO代表PSⅡ反應(yīng)中心開放時(shí)O點(diǎn)暗適應(yīng)最小熒光強(qiáng)度,與單位面積的熱耗散(在t=0時(shí)DIO/CSO)時(shí)相等;以吸收光能為基礎(chǔ)的性能指數(shù),PIABS≡(RC/ABS)·[φPo/(1-φPo)]·[ψO/(1-ψO)][7-9]。
1.2.3 棉花葉片耐熱性評價(jià)
對供試4種不同基因型棉花葉片的葉綠素?zé)晒庵笜?biāo)進(jìn)行評價(jià),用模糊隸屬函數(shù)法[10],對每項(xiàng)指標(biāo)求隸屬值,公式如下:
Y(μ)=(Y-Ymin)/(Ymax-Ymin).
公式中,Y指不同基因型棉花葉片某一項(xiàng)指標(biāo)測定值,Ymax為某一項(xiàng)指標(biāo)最大值,Ymin為指標(biāo)最小值,如果熒光參數(shù)與耐熱性呈負(fù)相關(guān),可以用反隸屬函數(shù)求函數(shù)值如:Y(μ)=1-(Y-Ymin)/(Ymax-Ymin)。
將不同溫度各個(gè)指標(biāo)耐熱隸屬值進(jìn)行累加,綜合值越大,耐熱性越強(qiáng)。
1.3 數(shù)據(jù)處理
數(shù)據(jù)用統(tǒng)計(jì)軟件SPSS 19.0進(jìn)行分析,單因素、多變量方差分析采用(LSD)和(Duncan)法進(jìn)行差異顯著性檢驗(yàn),采用Microsoft Excel 2013軟件作圖。
2 結(jié)果與分析
2.1 高溫對不同基因型棉花葉片熒光指標(biāo)的影響
2.1.1 高溫對不同基因棉花葉片F(xiàn)v/Fm的影響
Fv/Fm反映PSⅡ中心的光能轉(zhuǎn)換效率,F(xiàn)v/Fm降低表示植物光合抑制。研究表明,隨著溫度由25℃升至30℃時(shí),4個(gè)不同基因型棉花最大光化學(xué)效率Fv/Fm均顯著升高(除13-1×新海14號無顯著),與對照(25℃)相比,升高依次13-1×新海24號>新海24號>13-1品系>13-1×新海14號,但品系之間呈顯著性差異。35~45℃時(shí),13-1(品系)、新海24號下降幅度增大,13-1×新海24號和13-1×新海14號下降緩慢;13-1(品系)與新海24號之間有顯著性,13-1×新海24號、13-1×新海14號無顯著。當(dāng)溫度升高到48℃時(shí),最大光化學(xué)效率Fv/Fm除13-1×新海14號均顯著降低,降到最低是13-1品系,最高是13-1×新海14號,但品系之間差異不顯著。圖1
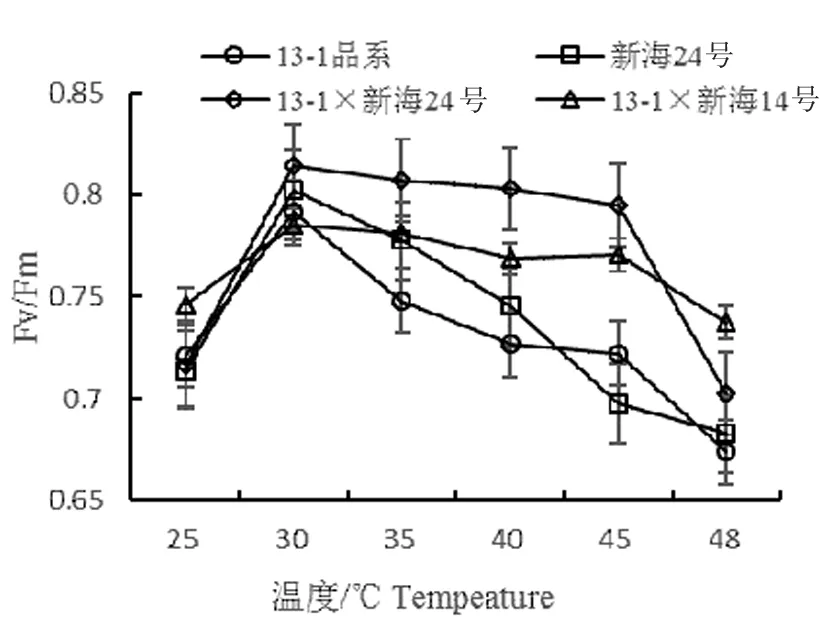
圖1 不同溫度下不同基因型棉花葉片的Fv/Fm變化
Fig.1 Effect of temperature on Fv/Fm in leaves of different varieties
2.1.2 高溫對不同基因棉花葉片φEo的影響
φEo反映了吸收的光量子將電子傳遞QA-下游的其它電子受體的比例,電子傳遞的量子產(chǎn)額φEo降低,電子受體比率降低。研究表明,隨著溫度升高,4個(gè)不同基因型ΦEo呈先升高后降低趨勢,30℃時(shí)出現(xiàn)峰值,新海24號、13-1×新海24號和13-1品系、13-1×新海14號差異顯著;48℃時(shí)降至最低,4個(gè)品系間無顯著。13-1×新海24號的φEo值在大于45℃迅速顯著下降,13-1品系、新海24號在40℃持續(xù)顯著下降。圖2
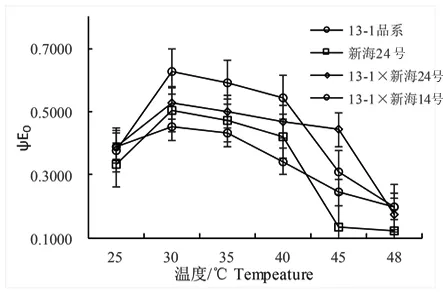
圖2 不同溫度下不同基因型棉花葉片的Fv/Fm變化
Fig.2 Effect of temperature on Fv/Fm in leaves of different varieties
2.1.3 高溫對不同基因棉花葉片F(xiàn)O的影響
初始熒光FO在暗適應(yīng)光系統(tǒng)中心完全開放的熒光產(chǎn)量狀態(tài),增加導(dǎo)致PSⅡ中心的失活性,衡量植物熱傷害的一種指標(biāo)。研究表明,隨著溫度不斷升高,30℃降到最低,48℃升到最高,13-1×新海24號呈極顯著差異,新海24號呈顯著差異,而13-1品系、13-1×新海14號無顯著性差異。圖3
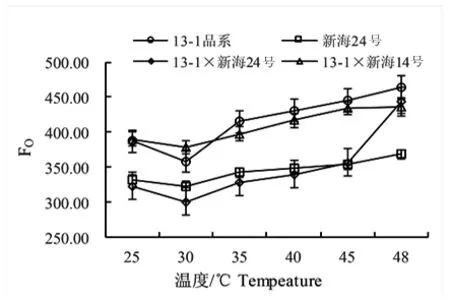
圖3 不同溫度下不同基因型棉花葉片的FO變化
Fig.3 Effect of temperature on FOin leaves of different varieties
2.1.4 高溫對不同基因棉花葉片DIO/CSO的影響
DIO/CSO反映了棉花葉片PSⅡ單位面積的熱耗散狀況,過剩熱耗散與內(nèi)囊體膜的能量有關(guān)。研究表明,隨著溫度不斷升高,DIO/CSO值在30℃降到最低,而后隨溫度的升高而升高,在48℃時(shí)DIO/CSO值升到最高,13-1(品系)、13-1×新海14號無顯著差異,新海24號呈顯著性差異,13-1×新海24號呈極顯著差異。圖4
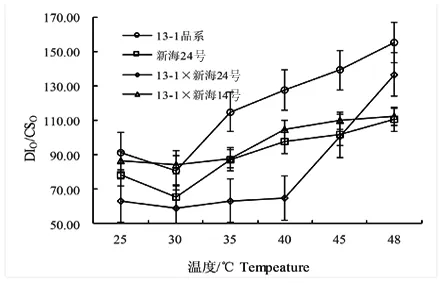
圖4 不同溫度下不同基因型棉花葉片DIO/CSO變化
Fig.4 Effect of temperature on DIO/CSOin leaves of different varieties
2.1.5 高溫對不同基因棉花葉片PIABS的影響
PIABS作為的光化學(xué)性能指數(shù),反映光合機(jī)構(gòu)的活性。研究表明,隨著溫度逐漸升高,4個(gè)不同基因型棉花性能指數(shù)PIABS均呈先上升后下降趨勢;48℃降至最低基本保持平衡;35~48℃時(shí),13-1×新海24號、新海24號、13-1×新海14號呈極顯著,13-1品系呈顯著,不同基因型棉花品種(系)在40℃差異顯著,其它溫度品系之間差異不顯著。圖5
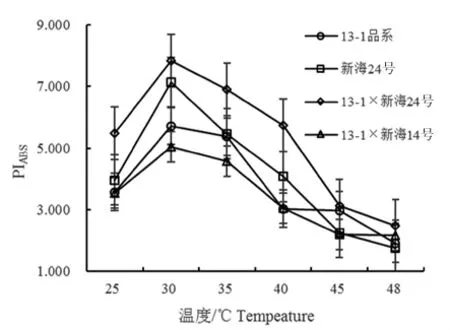
圖5 不同溫度下不同基因型棉花葉片的PIABS變化
Fig.5 Effect of temperature on PIABSin leaves of different varieties
2.2 不同基因型棉花葉片耐熱性評價(jià)
用模糊隸屬函數(shù)法,對4個(gè)不同基因型棉花葉片的3個(gè)葉綠素?zé)晒鈪?shù)進(jìn)行隸屬值計(jì)算。PIABS、φEO、Fv/Fm、FO、DIO/CSO指標(biāo)耐熱隸屬值進(jìn)行累加,得出不同基因型棉花葉片耐熱性綜合值。研究表明,不同基因型棉花葉片有差異的,13-1×新海24號耐熱性綜合值最大值9.76,是新海24號的1.52倍,13-1品系、13-1×新海14號各1.38倍。表1
表1 4個(gè)不同基因型棉花材料耐熱綜合評價(jià)值
Table 1 Heat value of four different genotype cotton materials
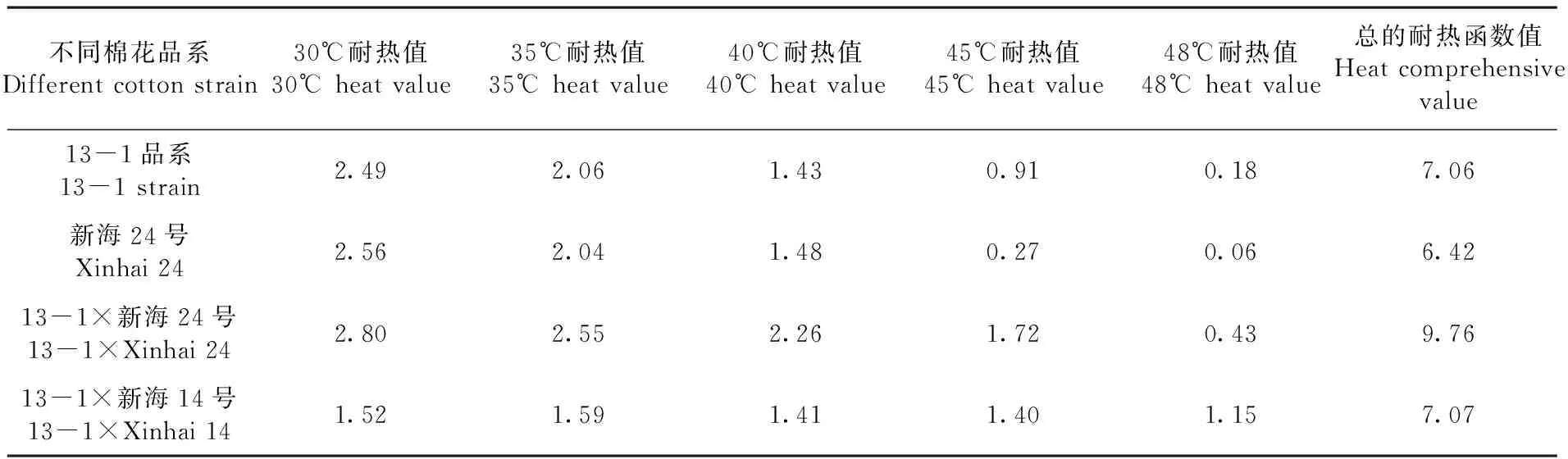
不同棉花品系Differentcottonstrain30℃耐熱值30℃heatvalue35℃耐熱值35℃heatvalue40℃耐熱值40℃heatvalue45℃耐熱值45℃heatvalue48℃耐熱值48℃heatvalue總的耐熱函數(shù)值Heatcomprehensivevalue13-1品系13-1strain2.492.061.430.910.187.06新海24號Xinhai242.562.041.480.270.066.4213-1×新海24號13-1×Xinhai242.802.552.261.720.439.7613-1×新海14號13-1×Xinhai141.521.591.411.401.157.07
3 討 論
當(dāng)前,葉綠素?zé)晒馓结樢呀?jīng)用到小麥、玉米、水稻作物抗逆的應(yīng)用,它能快速無損傷探測到高溫對光合特性的影響[11],也是評價(jià)植物耐熱性是否受損的良好指標(biāo)。因此,深入研究高溫對不同基因型棉花葉片PSⅡ的光合機(jī)構(gòu)的影響,具有重要意義。
研究中,在25~48℃,4種不同基因型棉花葉片F(xiàn)O、DIO/CSO隨著溫度升高而升高,這與張謐[12]Yamane[13]的研究認(rèn)為高溫導(dǎo)致FO的光合機(jī)構(gòu)和色素天線細(xì)胞分開,抑制了能量的傳遞,耗散剩多的光能研究一致。在高溫過程中,13-1×新海24號達(dá)到極顯著差異,新海24號呈顯著差異,而13-1品系、13-1×新海14號無顯著差異。FO升高反映了捕光色素復(fù)合體與PSⅡ的分離,也就是PSⅡ的中心內(nèi)囊體膜基粒減少。在35~40℃時(shí)4個(gè)不同基因型棉花品系FO顯著上升,說明PSⅡ和捕光色素復(fù)合體分離嚴(yán)重。在中等高溫下,初始熒光FO升高不顯著,說明對內(nèi)囊體膜變化是可逆的。其中,13-1×新海24號變化范圍最小,有利于在高溫下進(jìn)行光合反應(yīng),提高轉(zhuǎn)化效率。DIO/CSO反映了PSⅡ反應(yīng)中心以熱能消耗能量比率。前人研究表明:在高溫條件下,PSⅡ光抑制是QA-積累的電子傳遞鏈上電子向下傳遞減少,使PSⅡ反應(yīng)中心失去活性,轉(zhuǎn)成熱耗散能量,耗散過剩光能,隨著溫度升高,DIO/CSO增加[14]。研究表明:隨著溫度升高,DIO/CSO也增加,13-1×新海24號、新海24號呈顯著拐點(diǎn)在40℃,13-1×新海24號在小于40℃變化最平穩(wěn),耐熱性較強(qiáng),而13-1(品系)、13-1×新海14號無顯著差異,高溫拐點(diǎn)在35℃,耐熱性較弱。
在光能利用分配比率上,隨著溫度不斷升高,4種不同基因型棉花葉片PIABS、φEO、Fv/Fm先升高后下降趨勢,表明在大于30℃以上時(shí)最大光化學(xué)效率Fv/Fm降低主要是高溫影響了初始熒光FO增大和降低了最大熒光Fm,這是由于高溫導(dǎo)致PSⅡ放養(yǎng)復(fù)合體(OEC)的失活而產(chǎn)生。OEC失活,會導(dǎo)致QA接受電子減少,電子傳遞能力嚴(yán)重降低,導(dǎo)致PSⅡ中心受到嚴(yán)重破壞,這與前人[15]研究一致。φEO在大于30℃以上時(shí),PSⅡ放養(yǎng)復(fù)合體(OEC)會受到破壞,導(dǎo)致QA-大量電子增多,阻止了受體側(cè)QA電子傳遞到給QB,使量子產(chǎn)額φEO降低。性能指數(shù)包含(RC/CS、φPo和ψo(hù))三個(gè)指標(biāo),分別代表單位面積內(nèi)反應(yīng)中心的數(shù)量、最大光化學(xué)效率、反應(yīng)中心捕獲電子傳遞到電子傳遞鏈中QA來推動QA還原激子的比率,因此,準(zhǔn)確反映出棉花光合機(jī)構(gòu)整體狀況[16]。
不同基因型棉花葉片葉綠素?zé)晒鈪?shù)在高溫下的變化,表明4種不同基因型棉花葉片耐熱性不同,與13-1品系、13-1×新海14號、新海24號相比,13-1×新海24號在高溫PIABS、φEO、Fv/Fm下降比較平緩,F(xiàn)O、DIO/CSO上升緩慢,說明13-1×新海24號在高溫下可以降低PSⅡ中心受損,PSⅡ還能夠有少部分光化學(xué)反應(yīng)功能,熱耗散或者某些保護(hù)機(jī)制(光保護(hù)作用、葉黃素循環(huán)、類囊體膜脂)可以促進(jìn)向耐熱狀態(tài)轉(zhuǎn)變,對高溫具有較強(qiáng)熱穩(wěn)定性,更能適應(yīng)高溫,而13-1品系、13-1×新海14號、新海24號表現(xiàn)出在較高溫度下熱耗散與PSⅡ捕獲光能減弱,對高溫的耐受力較低。
4 結(jié) 論
高溫引起傷害PSⅡ受體電子傳遞受阻,放氧復(fù)合體受到損害,使PSⅡ中心光合反應(yīng)受到抑制,引起熱耗散增加,當(dāng)溫度繼續(xù)升高,會導(dǎo)致PSⅡ放養(yǎng)復(fù)合體受到嚴(yán)重破壞,捕獲光能減少,PSⅡ中心光合利用率降低。高溫脅迫明顯導(dǎo)致了4種不同基因型棉花耐高溫不同,利用方差分析及隸屬函數(shù)法等方法,對4種不同基因型棉花葉片葉綠素?zé)晒鈪?shù)PSⅡ功能綜合評價(jià)結(jié)果表明:性能指數(shù)PIABS在高溫下比Fv/Fm更靈敏,13-1×新海24號在高溫下表現(xiàn)出光化學(xué)效率、電子傳遞與熱耗散、吸收光能表現(xiàn)最好。13-1×新海24號在極端高溫占主要優(yōu)勢。
References)
[1] 歐祖蘭,曹福亮.植物耐熱性研究進(jìn)展[J]. 林業(yè)科技開發(fā),2008,(1):1-5.
OU Zu - lan, CAO Fu-liang. (2008). Reviewed of plant heat resistance [J].JournalofForestryEngineering, (1):1-5. (in Chinese)
[2]宋桂成,王苗苗,曾斌,等.高溫對棉花生殖過程的影響[J]. 核農(nóng)學(xué)報(bào),2016,(2):404-411.
SONG Gui-cheng, WANG Miao-miao, ZENG Bin, et al. (2016). The effects of high-temperature on reproductive process in upland cotton [J].JournalofNuclearAgriculturalSciences, (2):404-411. (in Chinese)
[3]王瑞華,郭峰,魏亦農(nóng),等. 高溫下不同葡萄品種葉綠素?zé)晒馓匦匝芯縖J]. 北方果樹,2013,(6):11-12.
WANG Rui-hua, GUO Feng, WEI Yi-nong, et al. (2013). Research on different grape varieties' chlorophyll fluorescence characteristics at high temperature [J].NorthernFruits, (6):11-12. (in Chinese)
[4]董文慶. 棉花雜種優(yōu)勢的光合生理基礎(chǔ)及耐高溫品種篩選指標(biāo)研究[D]. 南京: 南京農(nóng)業(yè)大學(xué)碩士論文, 2013.
DONG Wen-qing. (2013).StudyonPhotosyntheticbasisofheterosisandscreeningforhigh-temperaturetoleranceinCotton[D]. Master Dissertation. Nanjing Agricultural University, Nanjing. (in Chinese)
[5] 陳鋒,田紀(jì)春,孟慶偉,等. 短期高溫脅迫對高產(chǎn)小麥品系灌漿后期旗葉光系統(tǒng)Ⅱ功能影響 [J]. 應(yīng)用生態(tài)學(xué)報(bào),2006,(10):1 854-1 858.
CHEN Feng, TIAN Ji - chun, MENG Qing -wei, et al. (2006). Effects of short term high temperature stress on flag leaf photosystem ll functions of high-yielding wheat at late grain -filling stage [J].ChineseJournalofAppliedEcology, (10):1,854-1,858. (in Chinese)
[6] 杜國棟,呂德國,趙玲,等. 高溫對仁用杏光合特性及PSⅡ光化學(xué)活性的影響 [J]. 應(yīng)用生態(tài)學(xué)報(bào),2011,(3):701-706.
DONG Guo - dong, LV De - guo, ZHAO Ling, et al. (2011). Effects of high temperature on leaf photosynthetic characteristics and photosystemⅡphotochemical activity of kernel-used apricot [J].ChineseJournalofAppliedEcology,2011,(3):701-706. (in Chinese)
[7]Lazar, D. (2006). The polyphasic chlorophyll a fluorescence rise measured under high intensity of exciting light.FunctionalPlantBiology,33(1):9-30.
[8]李鵬民,高輝遠(yuǎn),Reto J.快速葉綠素?zé)晒庹T導(dǎo)動力學(xué)分析在光合作用研究中的應(yīng)用[J]. 植物生理與分子生物學(xué)學(xué)報(bào),2005,31(6):559-566.
LI Peng - ming, GAO Hui - yuan, Reto J. (2005). Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study [J].JournalofPlantPhysiologyandMolecularBiology, 31(6):559-566. (in Chinese)
[9]劉洪展.高溫脅迫對不同衰老型小麥品種光合機(jī)構(gòu)的影響[D]. 泰安:山東農(nóng)業(yè)大學(xué)碩士論文,2002.
LIU Hong - zhan. (2002).Effectsofhightemperaturestressonphotosyntheticapparatusofwheatvarietieswithdifferentsenescencetypes[D]. Master Dissertation. Shandong Agricultural University, Tai'an. (in Chinese)
[10]王海珍,韓路,徐雅麗,等.胡楊異形葉葉綠素?zé)晒馓匦詫Ω邷氐捻憫?yīng)[J]. 生態(tài)學(xué)報(bào),2011,31(9):2 444-2 453.
WANG Hai - zhen, HAN Lu, Xu Ya - li. (2011). Response of chlorophyll fluorescence characteristics of populus euphratica heteromorpjic leaves to high temperature [J].ActaEcologicaSinica, 31(9):2,444-2,453. (in Chinese)
[11]Maxwell, K., & Johnson, G. N. (2000). Chlorophyll fluorescence-a practical guide.JournalofExperimentalBotany, 51(345):659-668.
[12]張謐,王慧娟,于長青.超旱生植物沙冬青高溫脅迫下的快速葉綠素?zé)晒鈩恿W(xué)特征[J]. 生態(tài)環(huán)境學(xué)報(bào),2009,18(6):2 272-2 277.
ZHANG Mi, WANG Hui - juan, YU Chang - qing. (2009). The examination of high temperature stress of ammopiptanthus mongolicus by chlorophyll fluorescence induction parameters [J].EcologyandEnvironment, 18(6):2,272-2,277. (in Chinese)
[13]Yamane, Y., Shikanai, T., Kashino, Y., Koike, H., & Satoh, K. (2000). Reduction of q(a) in the dark: another cause of fluorescence f(o) increases by high temperatures in higher plants.PhotosynthesisResearch, 63(1):23-34.
[14]?quist, G., Hurry, V. M., Huner, N. P. A. (1993). The temperature dependence of the redox state of QA and susceptibility of photosynthesis to photoinhibition,PlantPhysiology&Biochemistry,31(5):683-691.
[15] 蘇曉瓊,王美月,束勝,等.外源亞精胺對高溫脅迫下番茄幼苗快速葉綠素?zé)晒庹T導(dǎo)動力學(xué)特性的影響[J]. 園藝學(xué)報(bào),2013,40(12):2 409-2 418.
SU Xiao - qiong, WANG Mei - yue, SHU Sheng, et al. (2013). Effects of exogenous Spd on the fast chlorophyll fluorescence induction dynamics in tomato seedlings under high temperature stress [J].ActaHorticulturaeSinica, 40(12):2,409-2,418. (in Chinese)
[16] Van Heerden, P. D., Strasser, R. J., & Krüger, G. H. (2004). Reduction of dark chilling stress in n-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics.PhysiologiaPlantarum, 121(2):239-249.
Supported by: Autonomous Region Key Technology R&D Program "Effects of high temperature on photosynthetic characteristics, chlorophyll fluorescence and antioxidant system of cotton" (31560406)
WEI Yi-nong(1964-), male, professor, poctoral degree, cotton genetics & breeding
Effects of Different Temperatures on Chlorophyll Fluorescence of Different Genotypes of Cotton Leaves
XING Shuang-tao, LI Zhi-bo, TANG Li-kui, ZHANG Rui-hai, WEI Yi-nong
(KeyLaboratoryofOasisEco-agricultureofXinjiangProductionandConstructionCorps,CollegeofAgronomy,ShiheziUniversity,ShiheziXinjiang832003,China)
【Objective】 This study aims to investigate the differences of leaf chlorophyll fluorescence characteristic parameters between different genotypes of cotton under different temperatures and to provide a theoretical basis to screen sui
Table indexes for evaluating high temperature - tolerance of cotton varieties. 【Method】Chlorophyll fluorescence characteristic parameters in leaves of cotton seedlings of four different genotypes (13-1 strain, Xinhai 24, 13-1×Xinhai 24 hybrid, 13-1×Xinhai 14 hybrid) were determined by rapid fluorescence detector (Handy PEA -100) under 25℃, 30℃, 35℃, 40℃, 45℃ and 48℃ respectively.【Result】The performance index of PIABS, maximum photochemical Fv/Fm and electron transfer yield φEo increased at first and decreased subsequently as the temperature increased, the heat dissipation DIO/CSO, initial fluorescence FOwas on the contrary. PIABSperformance index between the four different genotypes of strains was extremely significant at 40℃, and it could more accurately reflect photosynthetic status than Fv/Fm. 【Conclusion】Using the method of subordinate function to analyze the four different genotypes, it is found that 13-1×Xinhai 24(hybrid) has the highest subordinate function value, and we believe that it belongs to heat resistant strains.
cotton; photosynthetic mechanism; temperature; maximum photochemical efficiency; performance index
10.6048/j.issn.1001-4330.2017.03.002
2016-01-03
新疆維吾爾自治區(qū)科技支撐項(xiàng)目“高溫對棉花光合特性、葉綠素?zé)晒饧翱寡趸w系的影響”(31560406)
邢雙濤(1988-),男,陜西渭南人,碩士研究生,研究方向?yàn)樽魑镞z傳育種,(E-mail)xingst1988@163.com
魏亦農(nóng)(1964-),男,新疆庫爾勒人,教授,博士,研究方向?yàn)槊藁ㄟz傳育種,(E-mail)weiyinong@163.com
S562
A
1001-4330(2017)03-0403-06
邢雙濤,李志博,湯麗魁,趙瑞海,魏亦農(nóng)
(石河子大學(xué)農(nóng)學(xué)院/新疆生產(chǎn)建設(shè)兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室,新疆石河子 832000)
不同溫度對不同基因型棉花葉片葉綠素?zé)晒獾挠绊?/p>
邢雙濤,李志博,湯麗魁,趙瑞海,魏亦農(nóng)
(石河子大學(xué)農(nóng)學(xué)院/新疆生產(chǎn)建設(shè)兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室,新疆石河子 832000)
【目的】在不同溫度條件下,研究不同基因型棉花葉片葉綠素?zé)晒馓匦灾笜?biāo)的變化,為耐高溫棉花品種的篩選提供理論依據(jù)。【方法】分別在25、30、35、40、45和48℃的恒溫培養(yǎng)箱處理不同基因型棉花13-1品系、新海24號、13-1×新海24號(雜交種)、13-1×新海14號(雜交種),利用快速熒光測定儀(Handy PEA-100)測定不同基因型棉花葉片熒光特性主要參數(shù)。【結(jié)果】隨著溫度不斷升高,性能指數(shù)PIABS、最大光化學(xué)Fv/Fm、電子傳遞產(chǎn)額φEo表現(xiàn)為先升高后降低,而熱耗散DIO/CSO、初始熒光FO則先降低后升高;性能指數(shù)PIABS在40℃時(shí)4個(gè)不同棉花基因型棉花品系呈極顯著,比Fv/Fm更準(zhǔn)確地反映光合狀況。【結(jié)論】利用隸屬函數(shù)法分析4種不同基因型棉花品種(系),其中13-1×新海24號隸屬函數(shù)值最高,屬于耐熱品系。
棉花;溫度;最大光化學(xué)效率;性能指數(shù);葉綠素?zé)晒?/p>
0 引 言
【研究意義】光合作用在植物生長的過程中發(fā)揮著重要作用,且受環(huán)境的影響較大。高溫脅迫會導(dǎo)致光合作用受到抑制,影響光合效率[1]。棉花雖然是喜溫作物,但在生長發(fā)育與生殖發(fā)育過程中遇到35℃以上高溫,就會影響棉花得正常生長[2]。利用快速葉綠素?zé)晒鈨x來研究植物光合過程中光能利用,及外界環(huán)境因子對其影響,已成為作物逆境脅迫研究中的熱門技術(shù)[3]。因此研究不同基因型棉花葉片在不同高溫度下的葉綠素?zé)晒鈪?shù)變化規(guī)律,為鑒定棉花耐高溫材料提供依據(jù)。【前人研究進(jìn)展】董文慶等[4]發(fā)現(xiàn)棉花在不同生育時(shí)期受到高溫脅迫時(shí),不同的棉花品種熒光指標(biāo)Fv/Fm、FO、PIABS在蕾期差異最顯著,蕾期用Fv/Fm、FO、PIABS指標(biāo)量棉花光合耐熱性,性能指數(shù)PIABS比其它指標(biāo)更敏感地反映出棉花耐熱性。隨著溫度持續(xù)升高(25~50℃),對兩個(gè)不同基因型小麥光系統(tǒng)Ⅱ的能量熱耗散DIO/CSO差異不顯著,用于QA-以后的電子傳遞能量比例φEo表現(xiàn)顯著降低,初始熒光FO顯著升高,最大光化學(xué)效率Fv/Fm、性能指數(shù)PIABS均顯著下降[5]。杜國棟等(2011)[6]發(fā)現(xiàn)在高溫脅迫下,對仁用杏葉片放養(yǎng)復(fù)合體、PSⅡ反應(yīng)中心造成破壞,而且在50℃高溫導(dǎo)致初始熒光FO顯著升高,而Fv/Fm、PIABS分別顯著下降,導(dǎo)致高溫?fù)p害了PSⅡ受體側(cè)和供體側(cè)的功能,使光合機(jī)構(gòu)受害主要機(jī)制之一。【本研究切入點(diǎn)】目前相關(guān)研究僅在陸地棉品種中進(jìn)行,對于棉花不同的品種類型的研究相對較少。通過比較不同基因型品種及雜交種,進(jìn)一步研究棉花耐熱的生理機(jī)制。【擬解決的關(guān)鍵問題】在不同溫度條件下,研究不同基因型棉花葉片葉綠素?zé)晒馓匦灾笜?biāo)的變化,為進(jìn)一步選育耐高溫的棉花品種提供理論依據(jù)。
1 材料與方法
1.1 材 料
試驗(yàn)材料為13-1品系、新海24號、13-1×新海24號(雜交種)、13-1×新海14號(雜交種),以上品種(系)均由綠洲生態(tài)重點(diǎn)實(shí)驗(yàn)室育種室提供。品種(系)種植在新疆兵團(tuán)綠洲生態(tài)農(nóng)業(yè)重點(diǎn)實(shí)驗(yàn)室培養(yǎng)溫室。
1.2 方 法
1.2.1 試驗(yàn)設(shè)計(jì)
采取盆栽試驗(yàn),以基質(zhì)和蛭石按1∶1比例混合裝進(jìn)直徑30 cm,高25 cm的花盆,每盆種3株,每個(gè)品種類型3盆。放在溫室生長,定期澆適量的水,溫室的溫度20℃,棉花長到5~6片葉時(shí),開始放到恒溫培養(yǎng)箱處理,光照強(qiáng)度3 000 lx,濕度65%。分別以25、30、35、40、45和48℃的溫度處理8 h,處理之后測定棉花熒光特性。
1.2.2 熒光特性測定
采用英國Hansatech公司的植物效率分析儀(Handy PEA-100)對棉花葉片進(jìn)行OJIP曲線測定,曲線由3 000 μmol/(m2·s)的飽和光誘導(dǎo),測定前用葉夾暗適應(yīng)15 min,從曲線得到相應(yīng)參數(shù),最大光化學(xué)效率(在t=0時(shí)),F(xiàn)v/Fm=φPo≡TRO/ABS=1-(FO/Fm);吸收的光量子將電子傳遞QA-下游的其它電子受體的概率(在t=0時(shí)),φEo≡ETO/ABS= [1-(FO/Fm)]·ψO;FO代表PSⅡ反應(yīng)中心開放時(shí)O點(diǎn)暗適應(yīng)最小熒光強(qiáng)度,與單位面積的熱耗散(在t=0時(shí)DIO/CSO)時(shí)相等;以吸收光能為基礎(chǔ)的性能指數(shù),PIABS≡(RC/ABS)·[φPo/(1-φPo)]·[ψO/(1-ψO)][7-9]。
1.2.3 棉花葉片耐熱性評價(jià)
對供試4種不同基因型棉花葉片的葉綠素?zé)晒庵笜?biāo)進(jìn)行評價(jià),用模糊隸屬函數(shù)法[10],對每項(xiàng)指標(biāo)求隸屬值,公式如下:
Y(μ)=(Y-Ymin)/(Ymax-Ymin).
公式中,Y指不同基因型棉花葉片某一項(xiàng)指標(biāo)測定值,Ymax為某一項(xiàng)指標(biāo)最大值,Ymin為指標(biāo)最小值,如果熒光參數(shù)與耐熱性呈負(fù)相關(guān),可以用反隸屬函數(shù)求函數(shù)值如:Y(μ)=1-(Y-Ymin)/(Ymax-Ymin)。
將不同溫度各個(gè)指標(biāo)耐熱隸屬值進(jìn)行累加,綜合值越大,耐熱性越強(qiáng)。
1.3 數(shù)據(jù)處理
數(shù)據(jù)用統(tǒng)計(jì)軟件SPSS 19.0進(jìn)行分析,單因素、多變量方差分析采用(LSD)和(Duncan)法進(jìn)行差異顯著性檢驗(yàn),采用Microsoft Excel 2013軟件作圖。
2 結(jié)果與分析
2.1 高溫對不同基因型棉花葉片熒光指標(biāo)的影響
2.1.1 高溫對不同基因棉花葉片F(xiàn)v/Fm的影響
Fv/Fm反映PSⅡ中心的光能轉(zhuǎn)換效率,F(xiàn)v/Fm降低表示植物光合抑制。研究表明,隨著溫度由25℃升至30℃時(shí),4個(gè)不同基因型棉花最大光化學(xué)效率Fv/Fm均顯著升高(除13-1×新海14號無顯著),與對照(25℃)相比,升高依次13-1×新海24號>新海24號>13-1品系>13-1×新海14號,但品系之間呈顯著性差異。35~45℃時(shí),13-1(品系)、新海24號下降幅度增大,13-1×新海24號和13-1×新海14號下降緩慢;13-1(品系)與新海24號之間有顯著性,13-1×新海24號、13-1×新海14號無顯著。當(dāng)溫度升高到48℃時(shí),最大光化學(xué)效率Fv/Fm除13-1×新海14號均顯著降低,降到最低是13-1品系,最高是13-1×新海14號,但品系之間差異不顯著。圖1
圖1 不同溫度下不同基因型棉花葉片的Fv/Fm變化
Fig.1 Effect of temperature on Fv/Fm in leaves of different varieties
2.1.2 高溫對不同基因棉花葉片φEo的影響
φEo反映了吸收的光量子將電子傳遞QA-下游的其它電子受體的比例,電子傳遞的量子產(chǎn)額φEo降低,電子受體比率降低。研究表明,隨著溫度升高,4個(gè)不同基因型ΦEo呈先升高后降低趨勢,30℃時(shí)出現(xiàn)峰值,新海24號、13-1×新海24號和13-1品系、13-1×新海14號差異顯著;48℃時(shí)降至最低,4個(gè)品系間無顯著。13-1×新海24號的φEo值在大于45℃迅速顯著下降,13-1品系、新海24號在40℃持續(xù)顯著下降。圖2
圖2 不同溫度下不同基因型棉花葉片的Fv/Fm變化
Fig.2 Effect of temperature on Fv/Fm in leaves of different varieties
2.1.3 高溫對不同基因棉花葉片F(xiàn)O的影響
初始熒光FO在暗適應(yīng)光系統(tǒng)中心完全開放的熒光產(chǎn)量狀態(tài),增加導(dǎo)致PSⅡ中心的失活性,衡量植物熱傷害的一種指標(biāo)。研究表明,隨著溫度不斷升高,30℃降到最低,48℃升到最高,13-1×新海24號呈極顯著差異,新海24號呈顯著差異,而13-1品系、13-1×新海14號無顯著性差異。圖3
圖3 不同溫度下不同基因型棉花葉片的FO變化
Fig.3 Effect of temperature on FOin leaves of different varieties
2.1.4 高溫對不同基因棉花葉片DIO/CSO的影響
DIO/CSO反映了棉花葉片PSⅡ單位面積的熱耗散狀況,過剩熱耗散與內(nèi)囊體膜的能量有關(guān)。研究表明,隨著溫度不斷升高,DIO/CSO值在30℃降到最低,而后隨溫度的升高而升高,在48℃時(shí)DIO/CSO值升到最高,13-1(品系)、13-1×新海14號無顯著差異,新海24號呈顯著性差異,13-1×新海24號呈極顯著差異。圖4
圖4 不同溫度下不同基因型棉花葉片DIO/CSO變化
Fig.4 Effect of temperature on DIO/CSOin leaves of different varieties
2.1.5 高溫對不同基因棉花葉片PIABS的影響
PIABS作為的光化學(xué)性能指數(shù),反映光合機(jī)構(gòu)的活性。研究表明,隨著溫度逐漸升高,4個(gè)不同基因型棉花性能指數(shù)PIABS均呈先上升后下降趨勢;48℃降至最低基本保持平衡;35~48℃時(shí),13-1×新海24號、新海24號、13-1×新海14號呈極顯著,13-1品系呈顯著,不同基因型棉花品種(系)在40℃差異顯著,其它溫度品系之間差異不顯著。圖5
圖5 不同溫度下不同基因型棉花葉片的PIABS變化
Fig.5 Effect of temperature on PIABSin leaves of different varieties
2.2 不同基因型棉花葉片耐熱性評價(jià)
用模糊隸屬函數(shù)法,對4個(gè)不同基因型棉花葉片的3個(gè)葉綠素?zé)晒鈪?shù)進(jìn)行隸屬值計(jì)算。PIABS、φEO、Fv/Fm、FO、DIO/CSO指標(biāo)耐熱隸屬值進(jìn)行累加,得出不同基因型棉花葉片耐熱性綜合值。研究表明,不同基因型棉花葉片有差異的,13-1×新海24號耐熱性綜合值最大值9.76,是新海24號的1.52倍,13-1品系、13-1×新海14號各1.38倍。表1
表1 4個(gè)不同基因型棉花材料耐熱綜合評價(jià)值
Table 1 Heat value of four different genotype cotton materials
不同棉花品系Differentcottonstrain30℃耐熱值30℃heatvalue35℃耐熱值35℃heatvalue40℃耐熱值40℃heatvalue45℃耐熱值45℃heatvalue48℃耐熱值48℃heatvalue總的耐熱函數(shù)值Heatcomprehensivevalue13-1品系13-1strain2.492.061.430.910.187.06新海24號Xinhai242.562.041.480.270.066.4213-1×新海24號13-1×Xinhai242.802.552.261.720.439.7613-1×新海14號13-1×Xinhai141.521.591.411.401.157.07
3 討 論
當(dāng)前,葉綠素?zé)晒馓结樢呀?jīng)用到小麥、玉米、水稻作物抗逆的應(yīng)用,它能快速無損傷探測到高溫對光合特性的影響[11],也是評價(jià)植物耐熱性是否受損的良好指標(biāo)。因此,深入研究高溫對不同基因型棉花葉片PSⅡ的光合機(jī)構(gòu)的影響,具有重要意義。
研究中,在25~48℃,4種不同基因型棉花葉片F(xiàn)O、DIO/CSO隨著溫度升高而升高,這與張謐[12]Yamane[13]的研究認(rèn)為高溫導(dǎo)致FO的光合機(jī)構(gòu)和色素天線細(xì)胞分開,抑制了能量的傳遞,耗散剩多的光能研究一致。在高溫過程中,13-1×新海24號達(dá)到極顯著差異,新海24號呈顯著差異,而13-1品系、13-1×新海14號無顯著差異。FO升高反映了捕光色素復(fù)合體與PSⅡ的分離,也就是PSⅡ的中心內(nèi)囊體膜基粒減少。在35~40℃時(shí)4個(gè)不同基因型棉花品系FO顯著上升,說明PSⅡ和捕光色素復(fù)合體分離嚴(yán)重。在中等高溫下,初始熒光FO升高不顯著,說明對內(nèi)囊體膜變化是可逆的。其中,13-1×新海24號變化范圍最小,有利于在高溫下進(jìn)行光合反應(yīng),提高轉(zhuǎn)化效率。DIO/CSO反映了PSⅡ反應(yīng)中心以熱能消耗能量比率。前人研究表明:在高溫條件下,PSⅡ光抑制是QA-積累的電子傳遞鏈上電子向下傳遞減少,使PSⅡ反應(yīng)中心失去活性,轉(zhuǎn)成熱耗散能量,耗散過剩光能,隨著溫度升高,DIO/CSO增加[14]。研究表明:隨著溫度升高,DIO/CSO也增加,13-1×新海24號、新海24號呈顯著拐點(diǎn)在40℃,13-1×新海24號在小于40℃變化最平穩(wěn),耐熱性較強(qiáng),而13-1(品系)、13-1×新海14號無顯著差異,高溫拐點(diǎn)在35℃,耐熱性較弱。
在光能利用分配比率上,隨著溫度不斷升高,4種不同基因型棉花葉片PIABS、φEO、Fv/Fm先升高后下降趨勢,表明在大于30℃以上時(shí)最大光化學(xué)效率Fv/Fm降低主要是高溫影響了初始熒光FO增大和降低了最大熒光Fm,這是由于高溫導(dǎo)致PSⅡ放養(yǎng)復(fù)合體(OEC)的失活而產(chǎn)生。OEC失活,會導(dǎo)致QA接受電子減少,電子傳遞能力嚴(yán)重降低,導(dǎo)致PSⅡ中心受到嚴(yán)重破壞,這與前人[15]研究一致。φEO在大于30℃以上時(shí),PSⅡ放養(yǎng)復(fù)合體(OEC)會受到破壞,導(dǎo)致QA-大量電子增多,阻止了受體側(cè)QA電子傳遞到給QB,使量子產(chǎn)額φEO降低。性能指數(shù)包含(RC/CS、φPo和ψo(hù))三個(gè)指標(biāo),分別代表單位面積內(nèi)反應(yīng)中心的數(shù)量、最大光化學(xué)效率、反應(yīng)中心捕獲電子傳遞到電子傳遞鏈中QA來推動QA還原激子的比率,因此,準(zhǔn)確反映出棉花光合機(jī)構(gòu)整體狀況[16]。
不同基因型棉花葉片葉綠素?zé)晒鈪?shù)在高溫下的變化,表明4種不同基因型棉花葉片耐熱性不同,與13-1品系、13-1×新海14號、新海24號相比,13-1×新海24號在高溫PIABS、φEO、Fv/Fm下降比較平緩,F(xiàn)O、DIO/CSO上升緩慢,說明13-1×新海24號在高溫下可以降低PSⅡ中心受損,PSⅡ還能夠有少部分光化學(xué)反應(yīng)功能,熱耗散或者某些保護(hù)機(jī)制(光保護(hù)作用、葉黃素循環(huán)、類囊體膜脂)可以促進(jìn)向耐熱狀態(tài)轉(zhuǎn)變,對高溫具有較強(qiáng)熱穩(wěn)定性,更能適應(yīng)高溫,而13-1品系、13-1×新海14號、新海24號表現(xiàn)出在較高溫度下熱耗散與PSⅡ捕獲光能減弱,對高溫的耐受力較低。
4 結(jié) 論
高溫引起傷害PSⅡ受體電子傳遞受阻,放氧復(fù)合體受到損害,使PSⅡ中心光合反應(yīng)受到抑制,引起熱耗散增加,當(dāng)溫度繼續(xù)升高,會導(dǎo)致PSⅡ放養(yǎng)復(fù)合體受到嚴(yán)重破壞,捕獲光能減少,PSⅡ中心光合利用率降低。高溫脅迫明顯導(dǎo)致了4種不同基因型棉花耐高溫不同,利用方差分析及隸屬函數(shù)法等方法,對4種不同基因型棉花葉片葉綠素?zé)晒鈪?shù)PSⅡ功能綜合評價(jià)結(jié)果表明:性能指數(shù)PIABS在高溫下比Fv/Fm更靈敏,13-1×新海24號在高溫下表現(xiàn)出光化學(xué)效率、電子傳遞與熱耗散、吸收光能表現(xiàn)最好。13-1×新海24號在極端高溫占主要優(yōu)勢。
References)
[1] 歐祖蘭,曹福亮.植物耐熱性研究進(jìn)展[J]. 林業(yè)科技開發(fā),2008,(1):1-5.
OU Zu - lan, CAO Fu-liang. (2008). Reviewed of plant heat resistance [J].JournalofForestryEngineering, (1):1-5. (in Chinese)
[2]宋桂成,王苗苗,曾斌,等.高溫對棉花生殖過程的影響[J]. 核農(nóng)學(xué)報(bào),2016,(2):404-411.
SONG Gui-cheng, WANG Miao-miao, ZENG Bin, et al. (2016). The effects of high-temperature on reproductive process in upland cotton [J].JournalofNuclearAgriculturalSciences, (2):404-411. (in Chinese)
[3]王瑞華,郭峰,魏亦農(nóng),等. 高溫下不同葡萄品種葉綠素?zé)晒馓匦匝芯縖J]. 北方果樹,2013,(6):11-12.
WANG Rui-hua, GUO Feng, WEI Yi-nong, et al. (2013). Research on different grape varieties' chlorophyll fluorescence characteristics at high temperature [J].NorthernFruits, (6):11-12. (in Chinese)
[4]董文慶. 棉花雜種優(yōu)勢的光合生理基礎(chǔ)及耐高溫品種篩選指標(biāo)研究[D]. 南京: 南京農(nóng)業(yè)大學(xué)碩士論文, 2013.
DONG Wen-qing. (2013).StudyonPhotosyntheticbasisofheterosisandscreeningforhigh-temperaturetoleranceinCotton[D]. Master Dissertation. Nanjing Agricultural University, Nanjing. (in Chinese)
[5] 陳鋒,田紀(jì)春,孟慶偉,等. 短期高溫脅迫對高產(chǎn)小麥品系灌漿后期旗葉光系統(tǒng)Ⅱ功能影響 [J]. 應(yīng)用生態(tài)學(xué)報(bào),2006,(10):1 854-1 858.
CHEN Feng, TIAN Ji - chun, MENG Qing -wei, et al. (2006). Effects of short term high temperature stress on flag leaf photosystem ll functions of high-yielding wheat at late grain -filling stage [J].ChineseJournalofAppliedEcology, (10):1,854-1,858. (in Chinese)
[6] 杜國棟,呂德國,趙玲,等. 高溫對仁用杏光合特性及PSⅡ光化學(xué)活性的影響 [J]. 應(yīng)用生態(tài)學(xué)報(bào),2011,(3):701-706.
DONG Guo - dong, LV De - guo, ZHAO Ling, et al. (2011). Effects of high temperature on leaf photosynthetic characteristics and photosystemⅡphotochemical activity of kernel-used apricot [J].ChineseJournalofAppliedEcology,2011,(3):701-706. (in Chinese)
[7]Lazar, D. (2006). The polyphasic chlorophyll a fluorescence rise measured under high intensity of exciting light.FunctionalPlantBiology,33(1):9-30.
[8]李鵬民,高輝遠(yuǎn),Reto J.快速葉綠素?zé)晒庹T導(dǎo)動力學(xué)分析在光合作用研究中的應(yīng)用[J]. 植物生理與分子生物學(xué)學(xué)報(bào),2005,31(6):559-566.
LI Peng - ming, GAO Hui - yuan, Reto J. (2005). Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study [J].JournalofPlantPhysiologyandMolecularBiology, 31(6):559-566. (in Chinese)
[9]劉洪展.高溫脅迫對不同衰老型小麥品種光合機(jī)構(gòu)的影響[D]. 泰安:山東農(nóng)業(yè)大學(xué)碩士論文,2002.
LIU Hong - zhan. (2002).Effectsofhightemperaturestressonphotosyntheticapparatusofwheatvarietieswithdifferentsenescencetypes[D]. Master Dissertation. Shandong Agricultural University, Tai'an. (in Chinese)
[10]王海珍,韓路,徐雅麗,等.胡楊異形葉葉綠素?zé)晒馓匦詫Ω邷氐捻憫?yīng)[J]. 生態(tài)學(xué)報(bào),2011,31(9):2 444-2 453.
WANG Hai - zhen, HAN Lu, Xu Ya - li. (2011). Response of chlorophyll fluorescence characteristics of populus euphratica heteromorpjic leaves to high temperature [J].ActaEcologicaSinica, 31(9):2,444-2,453. (in Chinese)
[11]Maxwell, K., & Johnson, G. N. (2000). Chlorophyll fluorescence-a practical guide.JournalofExperimentalBotany, 51(345):659-668.
[12]張謐,王慧娟,于長青.超旱生植物沙冬青高溫脅迫下的快速葉綠素?zé)晒鈩恿W(xué)特征[J]. 生態(tài)環(huán)境學(xué)報(bào),2009,18(6):2 272-2 277.
ZHANG Mi, WANG Hui - juan, YU Chang - qing. (2009). The examination of high temperature stress of ammopiptanthus mongolicus by chlorophyll fluorescence induction parameters [J].EcologyandEnvironment, 18(6):2,272-2,277. (in Chinese)
[13]Yamane, Y., Shikanai, T., Kashino, Y., Koike, H., & Satoh, K. (2000). Reduction of q(a) in the dark: another cause of fluorescence f(o) increases by high temperatures in higher plants.PhotosynthesisResearch, 63(1):23-34.
[14]?quist, G., Hurry, V. M., Huner, N. P. A. (1993). The temperature dependence of the redox state of QA and susceptibility of photosynthesis to photoinhibition,PlantPhysiology&Biochemistry,31(5):683-691.
[15] 蘇曉瓊,王美月,束勝,等.外源亞精胺對高溫脅迫下番茄幼苗快速葉綠素?zé)晒庹T導(dǎo)動力學(xué)特性的影響[J]. 園藝學(xué)報(bào),2013,40(12):2 409-2 418.
SU Xiao - qiong, WANG Mei - yue, SHU Sheng, et al. (2013). Effects of exogenous Spd on the fast chlorophyll fluorescence induction dynamics in tomato seedlings under high temperature stress [J].ActaHorticulturaeSinica, 40(12):2,409-2,418. (in Chinese)
[16] Van Heerden, P. D., Strasser, R. J., & Krüger, G. H. (2004). Reduction of dark chilling stress in n-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics.PhysiologiaPlantarum, 121(2):239-249.
Supported by: Autonomous Region Key Technology R&D Program "Effects of high temperature on photosynthetic characteristics, chlorophyll fluorescence and antioxidant system of cotton" (31560406)
WEI Yi-nong(1964-), male, professor, poctoral degree, cotton genetics & breeding
Effects of Different Temperatures on Chlorophyll Fluorescence of Different Genotypes of Cotton Leaves
XING Shuang-tao, LI Zhi-bo, TANG Li-kui, ZHANG Rui-hai, WEI Yi-nong
(KeyLaboratoryofOasisEco-agricultureofXinjiangProductionandConstructionCorps,CollegeofAgronomy,ShiheziUniversity,ShiheziXinjiang832003,China)
【Objective】 This study aims to investigate the differences of leaf chlorophyll fluorescence characteristic parameters between different genotypes of cotton under different temperatures and to provide a theoretical basis to screen sui
Table indexes for evaluating high temperature - tolerance of cotton varieties. 【Method】Chlorophyll fluorescence characteristic parameters in leaves of cotton seedlings of four different genotypes (13-1 strain, Xinhai 24, 13-1×Xinhai 24 hybrid, 13-1×Xinhai 14 hybrid) were determined by rapid fluorescence detector (Handy PEA -100) under 25℃, 30℃, 35℃, 40℃, 45℃ and 48℃ respectively.【Result】The performance index of PIABS, maximum photochemical Fv/Fm and electron transfer yield φEo increased at first and decreased subsequently as the temperature increased, the heat dissipation DIO/CSO, initial fluorescence FOwas on the contrary. PIABSperformance index between the four different genotypes of strains was extremely significant at 40℃, and it could more accurately reflect photosynthetic status than Fv/Fm. 【Conclusion】Using the method of subordinate function to analyze the four different genotypes, it is found that 13-1×Xinhai 24(hybrid) has the highest subordinate function value, and we believe that it belongs to heat resistant strains.
cotton; photosynthetic mechanism; temperature; maximum photochemical efficiency; performance index
10.6048/j.issn.1001-4330.2017.03.002
2016-01-03
新疆維吾爾自治區(qū)科技支撐項(xiàng)目“高溫對棉花光合特性、葉綠素?zé)晒饧翱寡趸w系的影響”(31560406)
邢雙濤(1988-),男,陜西渭南人,碩士研究生,研究方向?yàn)樽魑镞z傳育種,(E-mail)xingst1988@163.com
魏亦農(nóng)(1964-),男,新疆庫爾勒人,教授,博士,研究方向?yàn)槊藁ㄟz傳育種,(E-mail)weiyinong@163.com
S562
A
1001-4330(2017)03-0403-06
