新疆羊種布魯氏菌分離株的鑒定與L7/L12蛋白的原核表達和生物信息學分析
2017-04-26 03:32:26劉升江雅麗付強史慧君李爽孟露萍郭飛張輝陳創夫
新疆農業科學 2017年3期
劉升,江雅麗,付強,史慧君,李爽,孟露萍,郭飛,張輝,陳創夫
(1.新疆石河子大學動物科技學院,新疆石河子 832000;2.新疆石河子大學生命科學學院,新疆石河子 832003;3.新疆農業大學動物醫學學院,烏魯木齊 830052;4.新疆石河子大學醫學院,新疆石河子 832003)
新疆羊種布魯氏菌分離株的鑒定與L7/L12蛋白的原核表達和生物信息學分析
劉升1,江雅麗2,付強3,史慧君3,李爽1,孟露萍1,郭飛4,張輝1,陳創夫1
(1.新疆石河子大學動物科技學院,新疆石河子 832000;2.新疆石河子大學生命科學學院,新疆石河子 832003;3.新疆農業大學動物醫學學院,烏魯木齊 830052;4.新疆石河子大學醫學院,新疆石河子 832003)
【目的】分離并鑒定新疆羊種布魯氏菌。原核表達該菌的L7/L12蛋白,檢測該蛋白的反應原性及進行部分生物學分析。【方法】采用細菌劃線培養、形態學觀察、PCR檢測以及生化試驗進行布魯氏菌分離鑒定。利用常規分子生物學方法表達并純化羊種布魯氏菌分離株的L7/L12蛋白,應用Western Blot分析融合蛋白的反應原性。使用生物信息學軟件對該蛋白進行了生物信息學分析。【結果】分離鑒定確定該菌株為羊種布魯氏菌。經過測序與酶切鑒定,正確構建了表達載體pET-28a-L7/L12. SDS-PAGE試驗顯示純化的L7/L12蛋白為單一條帶;經過Western Blot檢測,該融合蛋白具有良好的反應原性。生物信息學分析顯示,該蛋白無跨膜區結構,不存在信號肽,二級結構α-螺旋為主并利用 Phyre2 服務器成功構建了該蛋白的三維模型。【結論】鑒定出該分離菌株,表達并純化了該菌的L7/L12融合蛋白,Western Blot證明該蛋白具有良好的反應原性,為后續該蛋白的亞單位疫苗研究奠定了基礎。
羊種布魯氏菌;L7/L12;蛋白純化;反應原性;生物學
0 引 言
【研究意義】布魯氏菌病(Brucellosis)簡稱布病,是由革蘭氏染色陰性、兼胞內寄生的布魯氏菌(Brucella)引起的一種人獸共患傳染病[1]。動物感染后,主要引起母畜的不孕、流產和公畜的睪丸炎等[2]。人感染后,主要表現為發熱、脊柱炎、關節炎、骨髓炎等,嚴重時可導致患者喪失勞動力[3]。全世界每年超過50×104人被感染,造成的直接經濟損失近30×108美元[4]。新疆是我國重要的牧區,也是我國布魯氏菌病的高發區,據不完全統計,布魯氏菌病在新疆80余縣、市和13個兵團師局均有不同程度的流行,造成的經濟損失達幾個億[5-6]。研究對新疆羊種布魯氏菌分離株L7/L12蛋白進行表達、鑒定,生物信息學初步分析其結構性質,期待能對后續研制適合新疆本地化新型亞單位疫苗制備提供幫助。【前人研究進展】L7/L12是布魯氏菌的一種核糖體蛋白,該蛋白參與布魯氏菌胞內蛋白合成[7]。目前,對L7/L12蛋白的研究主要集中于保護性抗原和布魯氏菌致病過程中的作用機制研究。現有報道,利用L7/L12蛋白免疫動物后,可誘導機體產生細胞免疫和體液免疫應答,并且具有一定的保護力[8]。利用減毒沙門氏菌異源表達牛種布魯氏菌的L7/L12融合蛋白免疫動物后,可保護機體免受牛種布魯氏菌野毒株的感染[9]。此外,一些研究也發現,L7/L12蛋白能夠刺激牛外周血淋巴細胞的增殖,使其分泌IFN-γ,并且L7/L12蛋白對布魯氏菌脂多糖誘導的巨噬細胞的凋亡有促進作用[10]。Sergio等證明L7/L12蛋白可以誘導T細胞的活化,刺激CD4+細胞釋放IFN-γ因子,對牛布魯氏菌病具有較好的防御能力[11,12]。【本研究切入點】從新疆某羊場流產羔羊體內分離出鑒定出布魯氏菌,并原核表達其L7/L12蛋白,進行部分生物信息學分析。【擬解決的關鍵問題】研究新疆該羊場布魯氏菌的感染情況,為該羊場免疫措施的制定提供了依據。對L7/L12蛋白原核表達、生物信息學分析,為制備本地化該分離株新型亞單位疫苗提供依據。
1 材料與方法
1.1 材 料
1.1.1 羊胎兒與菌株
新疆地區某羊場流產胎兒1只,12 h內收集新鮮病料;E.coliDH5α、E.coliBL21(DE3)為研究室保存;pMD19-T Simple載體購自Takara公司;表達載體pET-28a購自Promega公司。
1.1.2 主要試劑
卡那霉素、氨芐霉素、IPTG和NC膜購自Solarbio公司;2×EsTaqMasterMix、普通瓊脂糖凝膠DNA回收試劑盒、高純度質粒小提試劑盒、DNA分子量Marker、辣根過氧化物酶標記兔抗羊IgG均購自北京康為世紀生物公司;限制性核酸內切酶(BamHⅠ、XhoⅠ)、T4DNA連接酶、solutionⅠ連接酶均購自Takara公司;膜封閉液、DAB增強型顯色試劑盒從天根生化科技有限公司購置;蛋白分子量Marker購自Thermo公司;蛋白純化HisTrapTM FF柱購自美國GE Healthcare公司,弗氏完全佐劑購自Sigma公司。
1.1.3 主要儀器
PCR儀(BIO-RAD);UVP凝膠成像系統;高速冷凍離心機(Sigma,2-16K);CO2恒溫培養箱(DNP-9162);Nanodrop 2000分光光度計;AKTAxpress智能多維純化系統(蛋白純化儀);生物安全柜(Esco,Class Ⅱ-biohazard safe cabinet);微量移液器(eppendorf)。
1.1.4 培養基
布魯氏菌固體培養基(TSA)、布魯氏菌液體培養基(TSB)、布魯氏菌鑒別培養基(BMB)、酵母提取物、胰蛋白胨購自英國OXOID公司;氯化鈉購置于上海生工生物工程股份有限公司。
1.2 方 法
1.2.1 引物
研究所用的引物均由北京六合華大基因科技股份有限公司合成。列出引物序列。表1
表1 PCR引物序列
Table 1 Nucleotide sequences of PCR primers
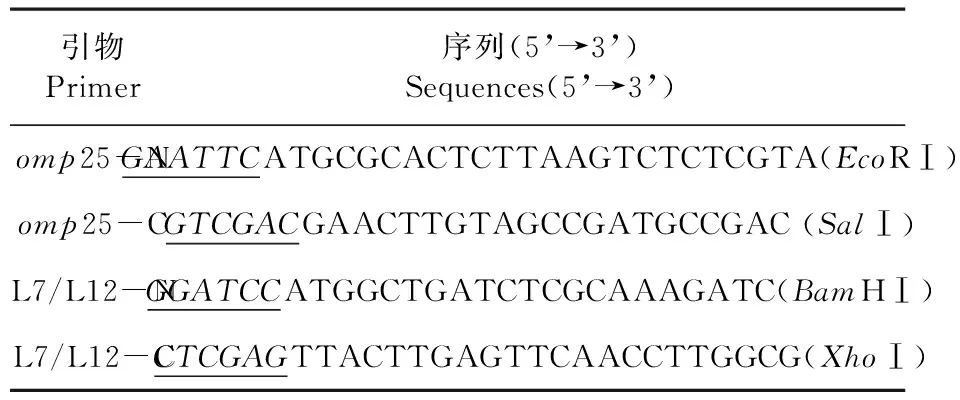
引物Primer序列(5’→3’)Sequences(5’→3’)omp25-NGAATTCATGCGCACTCTTAAGTCTCTCGTA(EcoRⅠ)omp25-CGTCGACGAACTTGTAGCCGATGCCGAC(SalⅠ)L7/L12-NGGATCCATGGCTGATCTCGCAAAGATC(BamHⅠ)L7/L12-CCTCGAGTTACTTGAGTTCAACCTTGGCG(XhoⅠ)
1.2.2 細菌分離與培養
P2級別實驗室內于生物安全柜中,取流產胎兒的脾臟劃線培養于布魯氏菌鑒別培養基,在37℃,5% CO2的環境中培養細菌。48~72 h后對細菌進行形態學觀察,利用改良萋-尼染色法對細菌染色后進行形態學觀察,初步鑒定陽性的布魯氏菌。
1.2.3 PCR鑒定
將初步鑒定為陽性的布魯氏菌擴大培養后,取0.5 mL菌液熱滅活后,作為PCR模版,利用omp25-N和omp25-C引物進行PCR擴增。PCR反應條件為:95℃ 5 min,94℃ 30 s,55℃ 45 s,72℃ 55 s,30個循環,72℃ 7 min。PCR產物用2%瓊脂糖凝膠電泳進行分析。
1.2.4 生化鑒定
將PCR鑒定為陽性的布魯氏菌按照《可感染人類的高致病性病原微生物菌(毒)種或樣本運輸管理規定》送至中國疾病預防控制中心傳染病預防控制所,對布魯氏菌進行生化鑒定,并命名。
1.2.5 L7/L12基因的擴增
以熱滅活的該分離株為模板,用L7/L12-N和L7/L12-C的引物,進行PCR擴增。PCR反應條件為:95℃ 5 min,94℃ 30 s,60℃ 30 s,72℃ 30 s,35個循環,72℃ 7 min。PCR產物用2%瓊脂糖凝膠電泳進行分析。
1.2.6 pMD19-T-L7/L12克隆載體的構建
PCR產物經過回收后,與pMD19-T simple載體連接。連接產物通過熱擊轉化的方法轉化至E.coli DH5α的感受態細胞中。經過氨芐抗性篩選和PCR鑒定后,將陽性克隆菌進行擴繁培養,提取質粒后,用限制性內切酶BamH I和XhoI進行酶切鑒定。將酶切鑒定正確的陽性菌送至北京六合華大基因科技股份有限公司進行DNA測序分析。
1.2.7 pET-28a-L7/L12表達載體的構建
將pMD19-T-L7/L12質粒和pET-28a質粒同時進行雙酶切,酶切后的pET-28a載體片段和L7/L12目的基因片段進行凝膠回收,將兩個片段使用solution I連接。連接產物通過熱擊轉化的方法轉化至E.coliBL21感受態細胞中。經過抗性篩選和PCR鑒定后,將陽性克隆菌進行擴繁培養,提取質粒后,用限制性內切酶BamHI和XhoI進行酶切鑒定。將酶切鑒定正確的陽性菌送至北京六合華大基因科技股份有限公司進行DNA測序分析。
1.2.8 重組蛋白L7/L12的誘導表達
將測序正確的克隆菌接種于LB液體培養基中,37℃過夜培養后,按1∶100的比例轉接至新鮮的LB液體培養基中,振蕩培養OD600nm≈0.4~0.6,取1 mL作為對照,剩余菌液中加入IPTG(終濃度為1 mmol/L)誘導表達。在至誘導后0、2、4、6和8 h分別取樣1 mL,離心后,棄去上清,對細菌沉淀進行處理后,SDS-PAGE電泳分析。
1.2.9 重組蛋白L7/L12的純化
將重組蛋白L7/L12誘導表達的菌體重懸于Lysis Buffer(50 mmol/L NaH2PO4,300 mmol/L NaCl,20 mmol/L imidazole,pH 8.0)中,液氮和42℃水浴反復凍融三次后超聲破菌,每次超聲10s,間歇10s,99次/周期,直至菌體清亮為止。12 000 r/min離心15 min,取上清和沉淀,分別進行SDS-PAGE電泳,檢測重組蛋白的表達位置。根據重組蛋白的表達位置,用AKTA express智能多維純化系統(蛋白純化儀)純化目的蛋白。將純化后的目的蛋白進行SDS-PAGE電泳,檢測蛋白的純化效果。
1.2.10 重組蛋白L7/L12的反應原性檢測
利用Western Blot分析重組蛋白L7/L12的反應原性。純化后的重組蛋白L7/L12經SDS-PAGE電泳后,轉膜至NC膜上,用封閉液37℃封閉1 h,隨后用TBST Buffer(20 mM Tris-HCl,150 mM NaCl,0.05%(v/v) Tween 20)漂洗三次;用布魯氏菌陽性血清作為一抗,37℃孵育1 h,TBST Buffer漂洗三次;用辣根過氧化物酶標記兔抗羊IgG(1∶5 000)作為二抗,37℃孵育1 h,TBST Buffer漂洗三次;利用DAB底物顯色試劑盒進行顯色,觀察重組蛋白L7/L12的反應原性。
1.2.11 生物信息學分析目的蛋白
通過 TMHMM Server v.2.0 軟件(http://www.cbs.dtu.dk/services/TMHMM/ ) 在線分析L7/L12蛋白的氨基酸序列,預測其蛋白跨膜區;應用SignalP 4.1 Server在線預測 L7/L12 蛋白的信號肽 (http://www.cbs.dtu.dk/services/SignalP/);SOPMA在線軟件預測 L7/L12蛋白的二級結構(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl );Phyre2 構建L7/L12蛋白三維結構模型(http://www.sbg.bio.ic.ac.uk/phyre2/webscripts/jobmonitor.cgi? Jobid = d97 9042d57b9240b);使用 PDBsum Generate( http: //www.ebi.ac.uk/thornton-srv/databases/ pdb-sum / Generate.html ) 在線對蛋白質結構模型進行評估。
2 結果與分析
2.1 布魯氏菌形態學觀察及omp25 PCR鑒定結果
取流產羊胎兒脾臟病料,培養72 h后,肉眼觀察到5個邊緣整齊、呈露滴狀、折光較亮的單克隆菌落,符合布魯氏菌的菌落形態。對5個單克隆菌落進行改良萋-尼染色,發現其中有1個菌落鏡下為單一的紅色短球桿菌。將該菌落擴繁后,進行PCR鑒定,在大約650 bp處出現了預期大小的條帶。通過以上的結果,初次判定布魯氏菌,命名為015株。圖1
2.2 生化鑒定
經中國疾病預防控制中心傳染病預防控制所對布魯氏菌的鑒定結果表明,從流產胎兒脾臟中分離獲得的布魯氏菌為羊種布魯氏菌生物3型。表2
2.3 L7/L12基因的擴增
以金屬浴滅活的015株為模板,高保真擴增出目的片段,位于375 bp左右,鑒定結果與預期一致。圖2
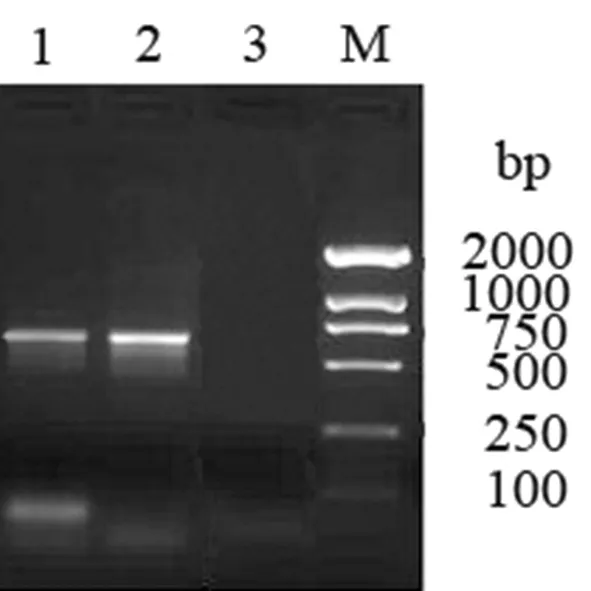
1:布魯氏菌015株;2:布魯氏菌16M株;3:陰性對照;M:DNA marker DL2 000
Lane 1:Brucella015; Lane 2:Brucella16M; Lane 3: negative control; Lane M: DNA marker DL2,000
圖1 布魯氏菌omp25基因PCR鑒定
Fig.1 Identification ofBrucellaomp25 gene PCR

1-3:PCR產物,4:陰性對照,M: DNA markerⅡ
Lane 1-3: the product of L7/L12 gene; Lane 4: negative control; Lane M: DNA markerⅡ
圖 2 L7/L12基因的擴增
Fig.2 Amplification of L7/L12 gene by PCR表2 布魯氏菌015株的生化鑒定結果
Table 2 Biochemical identification of theBrucella015 strain

項目Item015標準株 Thestandardstain16M544A1330SCO2需求 CO2requirement————H2S產生 H2Sproduction—++++++三勝黃素 Acriflavin————染料抑菌aDyeinhibition硫堇++—+復紅+++—噬菌體裂解實驗(RTDb)LysitictestofphageTbRTD——+—Tb104RTD——++Iz+BK2++++單項特異性血清凝集cAgglutinationtestofMonospecificserumA+—++M++——R————鑒定結果Results種羊羊牛豬生物型3111
a: 布魯氏菌固體培基中染料濃度20 μg/mL (1:50 000);b:RTD:常規試驗稀釋度(在增殖菌中完全裂解的最高稀釋度);c:A=A單因子血清,M=M單因子血清,R=R單因子血清;d:16M、544A、1 330S分別為羊種、牛種、豬種布魯氏菌標準株
a: Dye concetration ofBrucellaagar medium 20 μg/mL (1:50,000); b: RTD: The rac of rational test dilution (the highest dilution rate, at which the Tb phage completely lysesBrucellaspp; c: A-A mono-specific serum, M=M mono-specific serum, R=R mono-specific serum; d: 16M, 544A, 1,330S are the standard stains ofBrucellamelitensis,Brucellaabortus,Brucellasuis
2.4 pET-28a-L7/L12表達載體的構建
重組質粒pET28a-L7/L12經BamHⅠ和XhoⅠ雙酶切后,電泳圖片顯示得到5 369 bp的pET-28a 載體片段和375 bp 的目的條帶。結果表明pET28a-L7/L12原核表達載體已成功構建。圖3
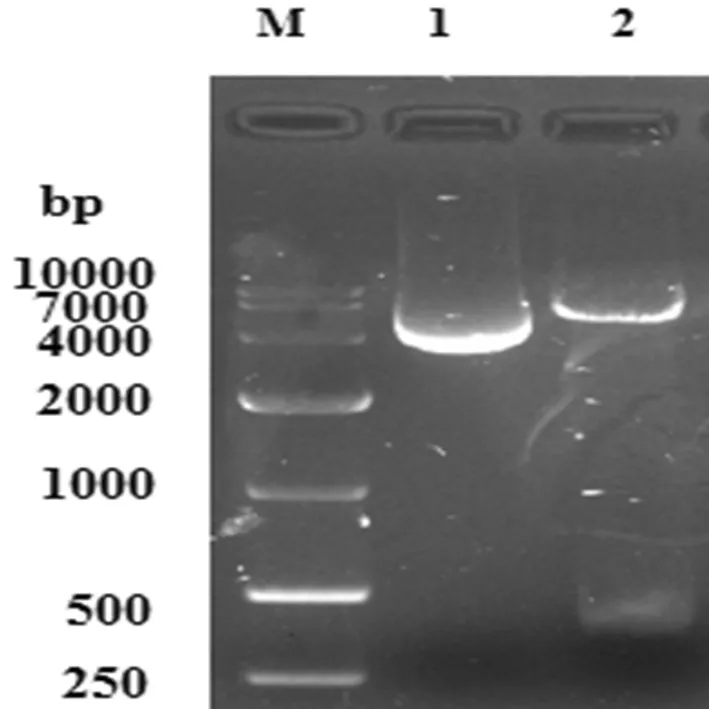
1:陰性對照;2:質粒pET-28a-L7/L12雙酶切;M:DNA marker DL10 000
Lane 1: the double digested plasmid of pET-28a-L7/L12 ; Lane 2: negative control;Lane M: DNA marker DL10,000
圖3 pET-28a-L7/L12雙酶切鑒定
Fig.3 The double digestion identification of pET-28a-L7/L12
2.5 重組蛋白L7/L12的誘導表達
將鑒定正確的pET-28a-L7/L12的重組質粒轉化至 BL21(DE3)感受態細胞中,篩選陽性重組菌后,IPTG誘導表達,SDS-PAGE結果表明目的蛋白位于大約19 ku處,與預計大小一致,且在8 h時表達量最高。圖4
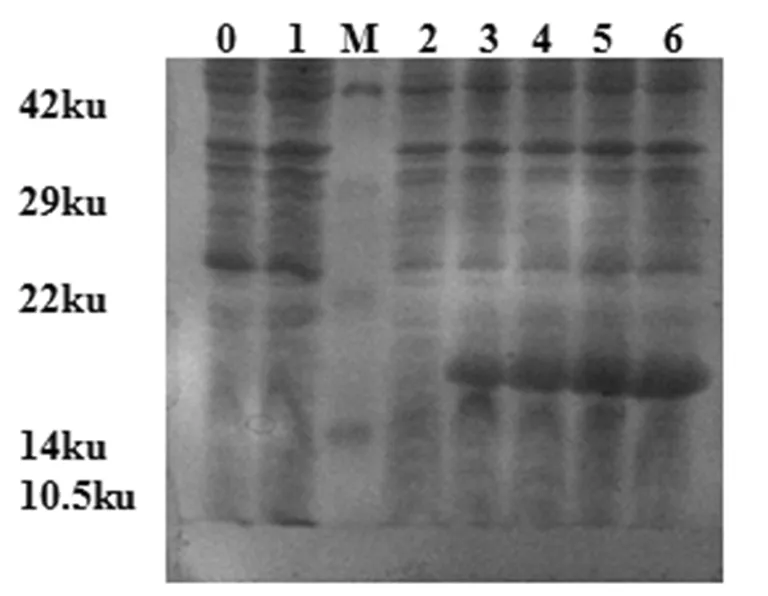
0: BL21(DE3)空菌誘導表達;1:轉化pET-28a質粒的BL21誘導表達;M:蛋白Marker;2-6:轉化pET-28a-L7/L12 質粒的BL21誘導表達0、2、4、6和8 h 0:induce expression BL21(DE3);1:induce expression BL21 transformed with pET-28a plasmid;M: Protein Marker; 2-6:induce expression BL21 transformed with pET-28a-L7/L12 plasmid for 0 h,2 h,4 h,6 h,8 h
圖4 表達產物的SDS-PAGE分析Fig4 The SDS-PAGE of pET-28a-L7/L12
2.6 重組蛋白L7/L12的純化
誘導8 h后的菌使用裂解液裂解后,取上清和沉淀驗證,SDS-PAGE結果表明,融合蛋白L7/L12在上清中存在(圖5a)。通過AKTA蛋白純化系統純化后的蛋白條帶清晰,雜帶較少,說明純化效果較好(圖5b)。圖5
2.7 重組蛋白L7/L12的反應原性檢測
將純化后的重組蛋白L7/L12轉至NC膜,采用羊布魯菌陽性血清作為一抗,HRP標記的兔抗羊抗體為二抗進行免疫印跡分析。Western Blot結果顯示在19 ku的位置上有一條明顯的特異性反應條帶,表明融合蛋白能被抗布魯氏菌的綿羊血清所識別,且目的蛋白具有良好的反應原性,與理論分析相符合。圖6
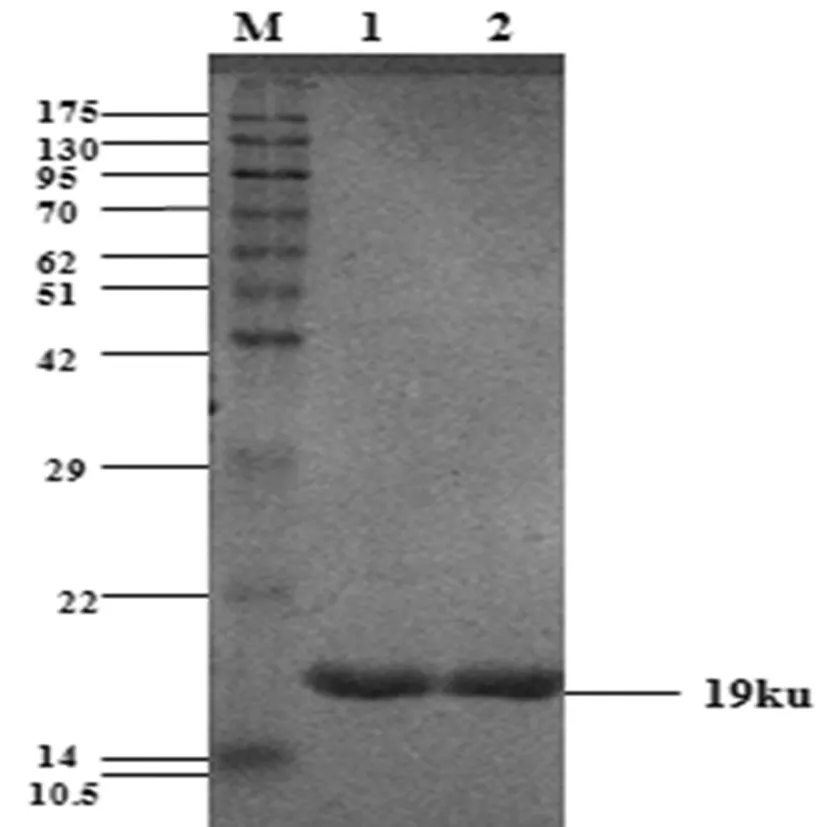
1-2:純化后的 L7/L12蛋白;M:蛋白MarkerSDS-PAGE analysis the effect of purified L7/L12 fusion protein 1-2:the purified L7/L12 protein ;M:Protein Marker
圖5 SDS-PAGE 電泳檢測L7/L12融合蛋白純化效果
Fig.5 SDS-PAGE analysis the effect of purified L7/L12 fusion protein

1-3:純化的His-L7/L12 融合蛋白;M:蛋白Marker
1-3: the purified His- L7/L12 fusion protein ;M: Protein Marker
圖6 融合蛋白His-L7/L12 Western Blot分析
Fig.6 The Western Blot analysis of fusion protein His-L7/L12
2.8 目的蛋白的生物信息學
通過TMHMM Server v.2.0在線軟件分析得出,目的蛋白無跨膜螺旋結構(圖7)。SignalP 4.1 Server 對該蛋白氨基酸序列分析顯示,該蛋白有信號肽(圖8)。利用 SOPMA 在線軟件分析該蛋白的二級結構,結果顯示,85個氨基酸參與形成α-螺旋,占68.55%。延伸鏈占8.87%,有13個氨基酸參與形成β-折疊,占10.48%,無規卷曲結構占12.10%(圖9)。通過 Phyre2 在線服務器構建出并優化目的蛋白的三維結構(圖10)。利用 PDBsum Generate在線評估(圖11)。從Ramachandran 圖中可以看出,在 121個氨基酸殘基中,有98個處于核心允許區,占 92.5% ,6 個處于額外允許區,占5.7% ,1個處于最大允許區,占 0.9% ,不可信區域為1,占 0.9%。這說明該模型具有一定的可信度。圖7~11
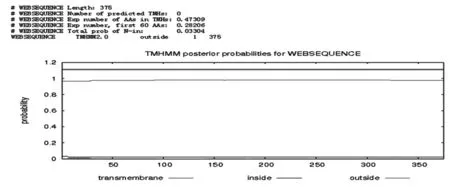
圖7 TMHMM Server v.2.0 預測L7/L12蛋白跨膜結構域
Fig.7 The prediction of transmembrane domain of L7/L12 protein by TMHMM Server v.2.0
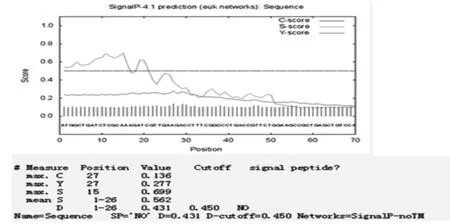
圖8 Signal P 4.1 Server 預測L7/L12蛋白信號肽
Fig.8 The prediction of signal peptide of L7/L12 protein by Signal P 4.1 Server
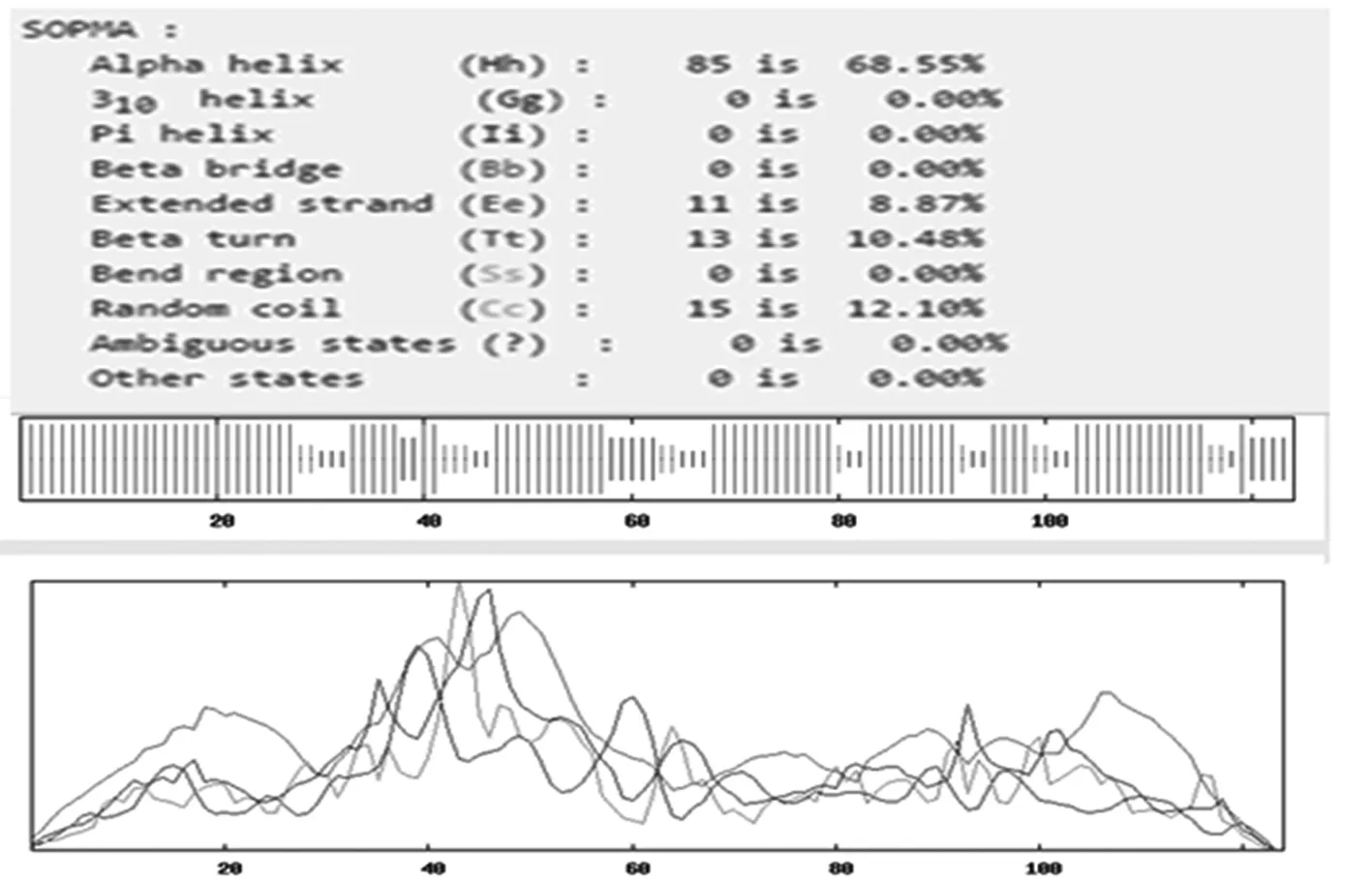
圖9 SOPMA 軟件推測的L7/L12蛋白二級結構
Fig.9 The prediction of secondary structure of L7/L12 protein by SOPMA Server
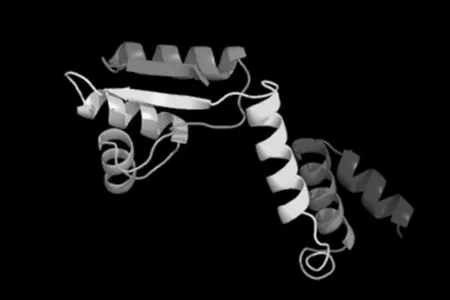
圖10 Phyre2 在線服務器預測 L7/L12 蛋白三級結構的構建
Fig.10 The construction of tertiary structure of L7/L12 protein by Phyre2
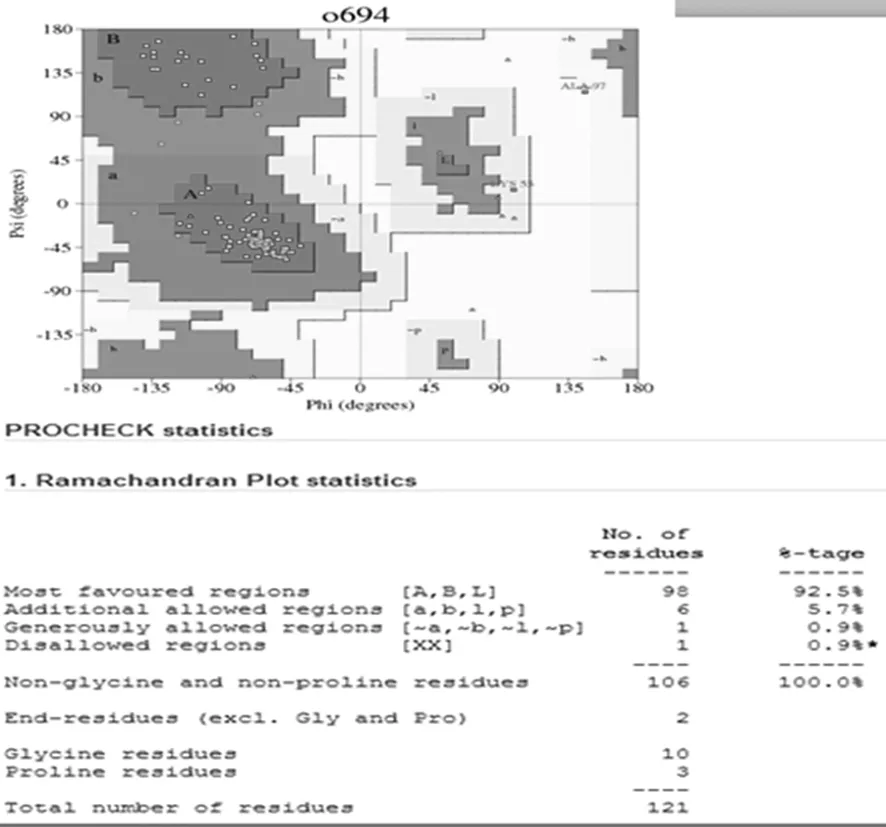
圖11 拉氏圖
Fig.11 The Ramachandran plot
3 討 論
布魯氏菌病是世界上具有嚴重危害性的人畜共患的傳染病之一,幾乎世界各地都有分布。從20世紀50年代中期,我國就開始了有組織有計劃地對人畜布病調查及防治[13]。20世紀80年代中后期,布魯氏菌病在世界部分地區有回升趨向,尤其在亞洲更為明顯[14]。90年代以來,布病疫情呈持續愈演愈烈的態勢,在我國西部、東北及華北省區疫情尤為嚴重。進入2000年以后,布病疫情強勢走高,每年報導的發病人數逐年上升,由總共報導的37種傳染病的第17位到2006年已上升至第10位,這種狀況在所有法定報告傳染病中是極其罕見的,進一步說明了當前布病疫情的嚴重性,研究及防治的迫切性[15]。
L7/L12核糖體蛋白是布魯氏菌重要的免疫性抗原之一,結構保守,是良好的疫苗候選抗原蛋白[16]。另外,L7/L12蛋白在布魯氏菌逃逸機體免疫應答后所引起的Ⅳ型超敏反應中也扮有重要角色[17]。除了引起細胞免疫反應外,L7/L12也可以引起體液免疫應答[18]。Tabynov 等[19,20]將L7/L12 蛋白與流感病毒載體結合包裹成類病毒顆粒作為疫苗抗原,免疫豚鼠和小鼠后發現該抗原可作為疫苗預防B.abortus544株所引起的動物感染。Luo等[21]研究表明L7/L12作為布魯氏菌疫苗抗原的機制之一是天然的L7/L12蛋白內含有T淋巴細胞的表位,從而能夠刺激機體產生Th1類細胞免疫應答,分泌IFN-γ刺激巨噬細胞活化,從而清除藏匿在巨噬細胞中的布魯氏菌。因此,研究由此構建新疆地區分離株的L7/L12蛋白表達,對于進一步研制新疆地區的亞單位疫苗具有現實意義。
試驗成功分離到新疆羊種生物3型布魯氏菌并成功構建L7/L12蛋白重組表達載體。表達的重組蛋白能與布魯氏菌陽性血清反應,說明表達的L7/L12在大腸桿菌中得到了正確折疊,其免疫學活性沒有受到影響。免疫印跡實驗證明了L7/L12蛋白具有良好的反應原性。
試驗運用生物信息學相關知識分析,結果表明該基因所表達的蛋白并沒有跨膜結構,說明該蛋白可能不參與細胞內外信號的轉導。該蛋白沒有信號肽,說明該蛋白屬于非分泌性蛋白,不能被布魯氏菌分泌到細胞外發揮作用。其二級結構主要以α-螺旋為主,和絕大多數蛋白的二級結構一樣,都是通過骨架上的羰基和酰胺基團之間形成的氫鍵維持該二級結構的穩定。利用生物信息學軟件Phyre2 等構建了出了L7/L12蛋白的三維模型,這有利于蛋白結構、功能的可視化分析。經分析發現,與其同源性的模型具有免疫調控功能,間接驗證了該蛋白可能具有免疫調控的功能。布魯氏菌的免疫逃避是一個復雜的過程,其分子機制尚不完全清楚。試驗通過構建布魯氏菌L7/L12基因的原核表達載體并對其反應原性進行鑒定以及生物信息學分析,為進一步探究布魯氏菌疫苗研究提供了一些參考。同時,該蛋白作為新疆該地區羊種布魯氏菌的疫苗候選分子具有一定的參考價值。
4 結 論
實驗以羊流產胎兒為病料,經過劃線培養、改良萋-尼染色法染色觀察其形態,對疑似菌進行omp25PCR鑒定,初步判斷出分離的菌中有布魯氏菌,命名為015。經中國疾病預防控制中心傳染病預防控制所對布魯氏菌通過一系列的生化鑒定,最終確定了該菌為羊種布魯氏菌生物3型,這表明新疆該羊場存在羊種布魯氏菌3型的感染。L7/L12蛋白參與布魯氏菌的蛋白質合成以及發揮保護性抗原的作用,為此通過原核表達的方式大量表達出L7/L12蛋白,并使用Western Blot方法檢測出該表達蛋白有良好的反應原性。生物學軟件對該蛋白進行了一些常規的預測,為了解該蛋白性質提供參考。
References)
[1] Pappas, G., Papadimitriou, P., Akritidis, N., Christou, L., & Tsianos, E. V. (2006). The new global map of human brucellosis.LancetInfectiousDiseases, 6(2):91-99.
[2] Ficht, T. A. (2003). Intracellular survival of brucella: defining the link with persistence.VeterinaryMicrobiology, 92(3):213-223.
[3] Godfroid, J., Cloeckaert, A., Liautard, J. P., Kohler, S., Fretin, D., & Walravens, K., et al. (2005). From the discovery of the malta fever's agent to the discovery of a marine mammal reservoir, brucellosis has continuously been a re-emerging zoonosis.VeterinaryResearch, 36(3):313-326.
[4] Pappas, G., Memish, Z., Earhart, K., Pappas, G., Sadek, R., & Hsueh, P. R. (2010). The changing brucella ecology: novel reservoirs, new threats.InternationalJournalofAntimicrobialAgents, 36 Suppl 1(5):8-11.
[5] 任德坤, 常青, 師茂林,等. 新疆布魯氏菌病疫情現狀與防治對策[J] . 疾病預防控制通報, 2008, 23(5):36-37.
REN De-kun, CHANG Qing, SHI Mao-lin, et al. (2008). Epidemic Sitation and Its Control of Brucellosis in Xinjiang[J] .Diseasepreventionandcontrolcommunications, 23(5):36-37.
[6] 閆晶華, 李金平, 米吉提,等. 新疆動物布魯氏菌病現狀分析[J] . 新疆畜牧業, 2011,(10):60.
YAN Jing-hua, LI Jin-ping, MI Jiti, et al. (2011). Present Situation Analysis of animal brucellosis[J] .XinjiangAnimalHusbandry, (10):60.
[7] Godinic-Mikulcic, V., Jaric, J., Greber, B. J., Franke, V., Hodnik, V., & Anderluh, G., et al. (2014). Archaeal aminoacyl-trna synthetases interact with the ribosome to recycle trnas.NucleicAcidsResearch, 42(8):5,191-5,201.
[8] Kurar, E., & Splitter, G. A. (1997). Nucleic acid vaccination of brucella abortus ribosomal l7/l12 gene elicits immune response.Vaccine, 15(17-18):1,851-1,857.
[9] Zhao, Z., Li, M., Luo, D., Xing, L., Wu, S., & Duan, Y., et al. (2009). Protection of mice from brucella infection by immunization with attenuated salmonella enterica serovar typhimurium expressing a l7/l12 and bls fusion antigen of brucella.Vaccine, 27(38):5,214-5,219.
[10] 曹滌非. 流產布魯氏菌 Omp25 與 L7/L12 促巨噬細胞凋亡的分子機制 [D]. 哈爾濱:東北農業大學博士論文, 2012.
CAO Di-fei. (2012).MolecularMechanismofAbortusofBrucellaOmp25andL7/L12InduceMacrophageApoptosis[D]. PhD Dissertation. Northeast Agricultural University, Harbin.(in Chinese)
[11] Oliveira, S. C., & Splitter, G. A. (1996). Immunization of mice with recombinant l7/l12 ribosomal protein confers protection against brucella abortus infection.Vaccine, 14(10): 959-962.
[12] Mallick, A. I., Singha, H., Chaudhuri, P., Nadeem, A., Khan, S. A., & Dar, K. A., et al. (2007). Liposomised recombinant ribosomal l7/l12 protein protects balb / c, mice against brucella abortus, 544 infection.Vaccine, 25(18):3,692-3,704.
[13] 姜順求.布病防治手冊[K] .北京:人民衛生出版社,1986: 264- 268.
JIANG Shun-qiu. (1986).BrucellosisPreventionManual[K]. Beijing: People's Medical Publishing House: 264- 268. (in Chinese)
[14] 尚德秋.布魯氏菌病再度肆虐及其原因[J] .中國地方病防治雜志, 2001, 16(1): 29- 34.
SHANG De-qiu. (2001). Brucellosis epidemic once again raging and its causes [J] .ChineseJournalofControlofEndemicDiseases, 16(1): 29- 34.(in Chinese)
[15] 崔步云.中國布魯氏菌病疫情監測與控制[J] .疾病監測,2007, 22(10): 649.
CUI Bu-yun. (2007). China brucellosis epidemic monitoring and control [J].DiseeasSuvreillance, 22(10): 649.(in Chinese)
[16] Oliveira, S. C., & Splitter, G. A. (1996). Immunization of mice with recombinant l7/l12 ribosomal protein confers protection against brucella abortus infection.Vaccine, 14(10): 959-962.
[17] Mallick, A. I., Singha, H., Chaudhuri, P., Nadeem, A., Khan, S. A., & Dar, K. A., et al. (2007). Liposomised recombinant ribosomal l7/l12 protein protects balb / c, mice against brucella abortus, 544 infection.Vaccine, 25(18):3,692-3,704.
[18] Al, D. S., Nckler, K., Scholz, H. C., Tomaso, H., Bogumil, R., & Neubauer, H. (2006). Immunoproteomic characterization of brucella abortus 1119-3 preparations used for the serodiagnosis of brucella infections.JournalofImmunologicalMethods, 309(1-2):34-47.
[19] Tabynov, K., Kydyrbayev, Z., Ryskeldinova, S., Yespembetov, B., Zinina, N., & Assanzhanova, N., et al. (2014). Novel influenza virus vectors expressing brucella l7/l12 or omp16 proteins in cattle induced a strong t-cell immune response, as well as high protectiveness against b. abortus infection.Vaccine, 32(18): 2,034-2,041.
[20] Tabynov, K., Sansyzbay, A., Kydyrbayev, Z., Yespembetov, B., Ryskeldinova, S., & Zinina, N., et al. (2014). Influenza viral vectors expressing the brucella omp16 or l7/l12 proteins as vaccines against b. abortus infection.VirologyJournal, 11(1): 69.
[21] Deyan Luo, Bing Ni, Peng Li, Wei Shi, Songle Zhang, Yue Han, Liwei Mao, Yangdong He, Yuzhang Wu, Xiliang Wang. (2006). Protective immunity elicited by a divalent dna vaccine encoding both the l7/l12 and omp16 genes of brucella abortus in balb/c mice.Infection&Immunity,74(5):2,734-2,741.
Supported by: General Programs of the National Natural Science Foundation of China"Study on the molecular regulation of TceSR and TcfSR in the pathogenesis of Brucella" (31572491).
CHEN Chuang-fu(1962-), male, professor, doctoral supervisor, mainly engaged in the study of pathogenic microorganisms pathogenic mechanism
Identification of Isolate Strain of Brucella Melitensis in Xingjiang and Prokaryotic Expression and Bioinformatics Analysis of Its L7/L12 Protein
LIU Sheng1, JIANG Ya-li2, FU Qiang3, SHI Hui-jun3, LI Shuang1, MENG Lu-ping1, GUO Fei4, ZHANG Hui1, CHEN Chuang-fu1
(1.CollegeofAnimalScienceandTechnology,ShiheziUniversity,ShiheziXinjiang832000,China; 2.CollegeofLifeTechnology,ShiheziUniversity,ShiheziXinjiang832000,China; 3.CollegeofVeterinaryMedicine,XinjiangAgriculturalUniversity,Urumqi830052,China; 4.SchoolofMedicine,ShiheziUniversity,ShiheziXinjiang832000,China)
【Objective】 To isolate and identify Xinjiang Brucella melitensis, this project focuses on prokaryotic expression of the L7/L12 protein of the bacteria,detection of its reactionogenicity and and partial biological analysis. 【Method】Isolation and identification of Brucella melitensis by using bacterial streak culture, morphological observation, biochemical test and PCR detection. Using conventional and molecular biological methods to express and purify the L7/L12 protein of this Brucella melitensis. Expression and purification isolate strain L7/L12 protein by using the conventional molecular biological methods, and analysis of the fusion protein reactionogenicity by Western Blot . Using bioinformatics software to analyze some functions of this protein. 【Result】After identification, the strain was identified as Brucella melitensis. After enzyme digestion and sequencing, the expression vector pET-28a-L7/L12 was correctly constructed. SDS-PAGE tests showed that the purified L7/L12 protein was a single band. The fusion protein had good reactionogenicity by Western Blot detection. Bioinformatics analysis showed the protein had no trans-membrane domain and no signal peptide.Its secondary structure was mainlyα-helix.And the three-dimensional structure of the protein was constructed by Phyre2 Server. 【Conclusion】The isolated strain was identified successfully and L7/L12 fusion protein of this isolate strain was expressed and purified. Blot Western test proved that the protein had a good reactionogenicity, which laid the foundation for the protein follow-up research of the subunit vaccine.
Brucellamelitensis;L7/L12; protein purification; reactionogenicity;biological analysis
10.6048/j.issn.1001-4330.2017.03.022
2016-11-24
國家自然科學基金面上項目“TceSR和TcfSR在布魯氏菌致病過程中的分子調控機制研究”(31572491)
劉升(1992-),女,安徽宿州人,碩士研究生,研究方向為分子病毒學,(E-mail)1341753096@qq.com
陳創夫(1962-),男,廣東潮州人,教授,博士生導師,研究方向為病原微生物致病機理,(E-mail)ccf-xb@163.com
S852.1;S188
A
1001-4330(2017)03-0564-10
猜你喜歡
國畫家(2022年2期)2022-04-13 09:07:46
四川文學(2021年4期)2021-07-22 07:11:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
絲綢之路(2014年9期)2015-01-22 04:24:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
兒童與健康(2011年4期)2011-04-12 00:00:00
新疆人文地理(2009年7期)2009-09-29 09:56:14
